Serranus scriba (Linnaeus, 1758)
Окунь-зебра, каменный окунь (Rus),
Painted comber
(Eng)
Синонимы:
Holocentrus argus Spinola, 1807
Holocentrus maroccanus Bloch & Schneider, 1801
Perca scriba Linnaeus, 1758
Serranus papilionaceus Valenciennes, 1832
Атлас-определитель рыб: Книга для учащихся. Н.А.Мягков 1994 г.
Окунь каменный — Serranus scriba (Linnaeus, 1758)
Тело высокое и заметно сжатое с боков. Один спинной плавник. Хвостовой плавник без выемки, чуть выпуклый. Зубы характерные, щитинковидные, но в передней части челюстей есть и клыковидные. D 11—16. Окраска тела зеленовато-серая с бурым налетом. По бокам проходит несколько темных поперечных полос. Длина тела около 30 см. Обитает среди прибрежных камней в Черном море.
Декоративное рыбоводство. А.М.Кочетов 1991 г.
В Черном море — окунь-зебра (Serranus scriba). Тело умеренно удлиненное, сжато с боков. Рыбы — хищники. Известны случаи нереста в неволе. Аквариум от 300 л.
Иллюстрированная энциклопедия рыб. Ст.Франк 1983 г.
Каменный окунь (Serranus scriba) и Serranus cabrilla интересен тем, что является гермафродитом. Обе половые железы развиваются одновременно и осуществляется самооплодотворение. Каменный окунь, обитающий в Адриатическом море, обычно держится среди прибрежных камней и скал. Достигает 28 см. Его основная окраска оранжевая до красной с поперечными синеватыми полосами. У этих окуней весьма вкусное мясо.
Ихтиопланктон Черного моря. Т.В.Дехник 1973 г.
КАМЕННЫЙ ОКУНЬ — SERRANUS SCRIBA (LINNE)
Каменный окунь — функциональный гермафродит с возможным в естественных условиях перекрестным оплодотворением и самооплодотворением (Dufosse, 1856; Van Oordt, 1929; D'Ancona, 1949; Салехова, 1963).
В Черном море впервые искусственное оплодотворение икры спермой той же особи произвела К. С. Ткачева (1952). Икра оплодотворилась и через полтора часа наблюдалось образование зародышевого диска. Детальные исследования Л. П. Салеховой (1963) по размножению и развитию икры каменного окуня показали, что эмбриональное развитие протекает нормально как при оплодотворении икринок спермой той же особи, так и при перекрестном оплодотворении. Из наблюдений за выживанием икры в процессе развития выявлено, что более жизнестойкой оказывается икра, полученная перекрестным оплодотворением. Среди личинок, выклюнувшихся из самооплодотворенной икры, были уродливые формы, тогда как из икры перекрестного оплодотворения все выклюнувшиеся личинки были нормальными.
Условия размножения. Нерест каменного окуня в Черном море происходит летом.
Первые сведения о нахождении икринок в планктоне приведены в работе В. А. Водяницкого (1930а), который сообщает, что в июле и начале августа икринки этого вида в очень большом количестве вылавливались у входа в Новороссийскую бухту. По наблюдениям Е. Г. Косякиной (1938), икринки встречались в планктоне Новороссийской бухты с июня до середины сентября, в наибольшем количестве — в конце июня — начале июля.
В районе Карадага нерест Serranus scriba происходит с июля по сентябрь (Виноградов, 1948; Ткачева, 19526; Овен, 1959). У Севастополя икринки встречались в июле — августе (Дука, 1959). У Кавказского побережья нахождение икринок отмечено в июле при температуре воды 20,6—25,7° и солености — 15—18,3‰ (Дехник, Павловская, 1950).
В Одесском заливе икринки и личинки каменного окуня не обнаружены (Зайцев, 1959а). В Азовском море в районе Казантипского маяка при температуре воды на поверхности 24,4° и солености 11,55‰ нами были найдены в планктоне нормально развивающиеся икринки каменного окуня.
В Средиземном море нерест Serranus scriba начинается в мае и заканчивается в августе (Lo Bianco, 1908—1909).В период наших наблюдений в Средиземном море икрннки Serranus sp. были отмечены в уловах в июне и июле при температуре воды 20,5—25,1° и солености 37,5—38,5‰.
Таким образом, как в Черном море, так и в Средиземном Serranus scriba размножается в наиболее теплое время года. Развитие икринок и личинок этого вида происходит, с одной стороны, при значительном диапазоне солености (15—38,5‰), с другой — при сравнительно небольших колебаниях температуры воды (19—26°).
Суточный ритм размножения. Вымет и оплодотворение икры каменного окуня происходят в сумеречное время. Икринки на этапе дробления встречаются в планктоне между 16 и 02 ч; максимальное количество вылавливается в период между 20 и 24 ч (табл. 14). Только что выметанные икринки с обозначающимся протоплазматическим бугорком встречаются начиная от 16 до 22 ч; в последующие часы (22—02) икринки находятся на более поздних стадиях развития. Далее в течение суток последовательно прослеживается развитие каждого суточного вымета (Дехник, 1961).

Наблюдения Л. П. Салеховой (1963) за изменением зрелости гонад в течение суток показали, что в ночные, утренние и дневные часы только единичные рыбы имели гонады с текущими молоками; рыб с текучей икрой не было. В 16—20 ч у большинства окуней гонады были готовы к выбою: при легком нажиме на брюшко вытекала зрелая икра.
Строение икринок. Икринки каменного окуня пелагические, сферической формы, с одной маленькой жировой каплей. Оболочка прозрачная, желток гомогенный. Диаметр живых икринок колеблется, по нашим данным, от 1,00 до 1,13 мм (в среднем 1,03 мм;), жировой капли — от 0,12 до 0,15 мм (в среднем 0,13 мм). Размер икринок, по В. А. Водяницкому и И. И. Казановой (1954),— 0,99 мм, жировой капли — 0,12—0,15 мм. В Средиземном море икринки несколько мельче — 0,90 мм, жировая капля — 0,12 мм (Raffaele, 1888).
Икринки Serranus scriba сходны с икринками другого вида — Serranus cabrilla. Размер икринок и жировых капель одинаков, характер пигментации развивающихся зародышей совершенно сходен (Raffaele, 1888; Bertoline, 1933) В основном сходны и выклюнувшиеся личинки обоих видов.
Эмбриональное развитие. Описание эмбрионального развития каменного окуня составлено в основном по наблюдениям Л. П. Салеховой (1963) за искусственно оплодотворенной икрой. Некоторая детализация строения развивающегося зародыша дана по нашим наблюдениям над икринками, выловленными в море.
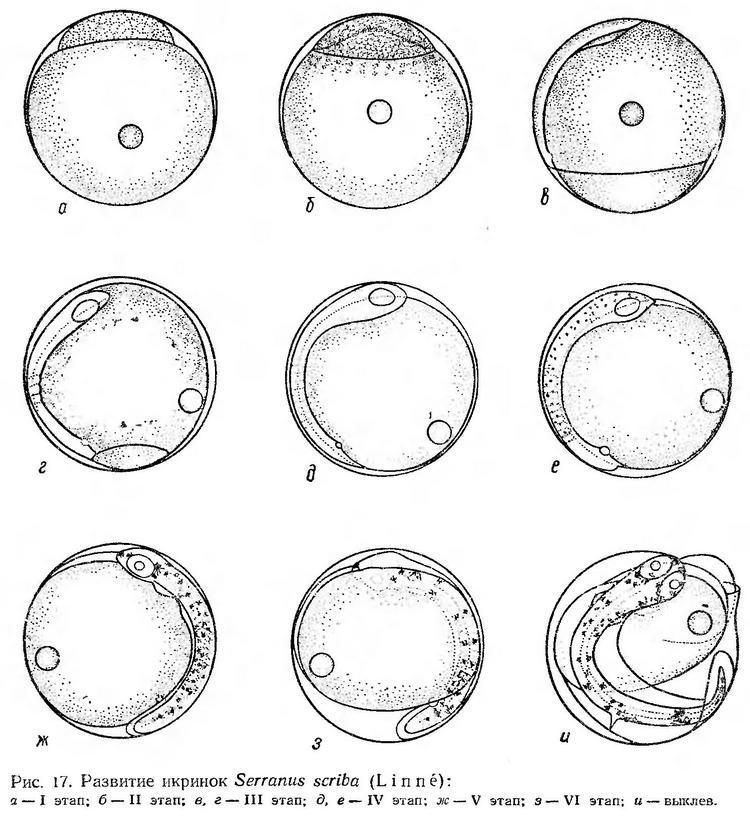
I этап — дробление бластомеров при температуре воды 24—24,6° продолжается 3,5—4,5 ч. Бластодиск занимает небольшую часть поверхности желтка. Длина его основания 0,63—0,65 мм, высота — 0,13—0,14 мм (рис. 17, а, табл. 15).
Образование эпителиальной бластулы при указанной температуре происходит в течение двух часов. Длина основания бластодиска и его высота
заметно увеличиваются; в перибласте появляются пузыревидные образования (рис. 17, б).
III этап — обрастание желтка бластодермой и гаструляция — наиболее продолжительный. Через 4—4,5 ч после начала обрастания края бластодиска доходят до середины желтка. Намечается зародышевая полоска. Когда бластодиск охватывает около 3/4 поверхности желтка, расширенный головной конец достигает анимального полюса (рис. 17, в). Незадолго перед замыканием бластопора намечается нервный ствол, глазные бокалы, появляются первые туловищные сегменты, купферов пузырек (рис. 17, г). Процесс гаструляции заканчивается в возрасте 15—16 ч.
После замыкания бластопора число миотомов быстро увеличивается. Обособляются глаза, начинают формироваться мозговые доли (рис. 17, д). В конце этапа перед образованием хвостовой почки намечаются слуховые капсулы, на голове и на теле появляются точечные клетки черного и желтого пигмента (рис. 17, е). IV этап длится 3—4 ч.
В начале V этапа развития купферов пузырек исчезает. Наряду с ростом хвостового отдела дифференцируются хвостовые миотомы. Образуются зачатки сердца, кишечника, печени. В глазах появляются хрусталики, намечается плавниковая кайма (рис. 17, ж). В конце V этапа (через 3—3,5 ч после появления хвостовой почки при температуре воды 24°) отмечается характерное расположение пигмента: 2 пары крупных желтых клеток сосредоточены в преорбитальной и посторбитальной областях и 5 симметричных пар клеток — в туловищном и хвостовом отделах на дорсальной и вентральной сторонах тела. Между ними, главным образом на спине, довольно равномерно располагаются точечные меланофоры.
К началу пульсации сердца хвост эмбриона немного заходит за середину желтка (рис. 17, з). Эмбрион слабо подергивается. Сердце пульсирует 84—86 раз в минуту. Намечаются зачатки грудных плавников. Происходит развитие всех внутренних органов. Плавниковая кайма широкая. Эмбрион быстро растет. К моменту выклева он охватывает 2/3 поверхности желтка. Желтый пигмент сгруппирован в 7 пар крупных пятен. Точечные меланофоры равномерно расположены на спинной стороне тела (рис. 17, и). На жировой капле пигмент имеется не всегда. Пульсация сердца учащается. Перед выклевом частота биения достигает 100—120 ударов в минуту. Выклев происходит с головного конца. Голова упирается в оболочку (рис. 17, и). В этом месте происходит разрыв, через который выпячивается сначала передняя часть желточного мешка, затем голова, туловище и хвост эмбриона. VI этап при температуре воды 21,8—24,0° длится 12—15 ч.
Весь процесс эмбрионального развития каменного окуня при температуре воды 21,8—24,6° продолжался 22—28 ч (табл. 15).

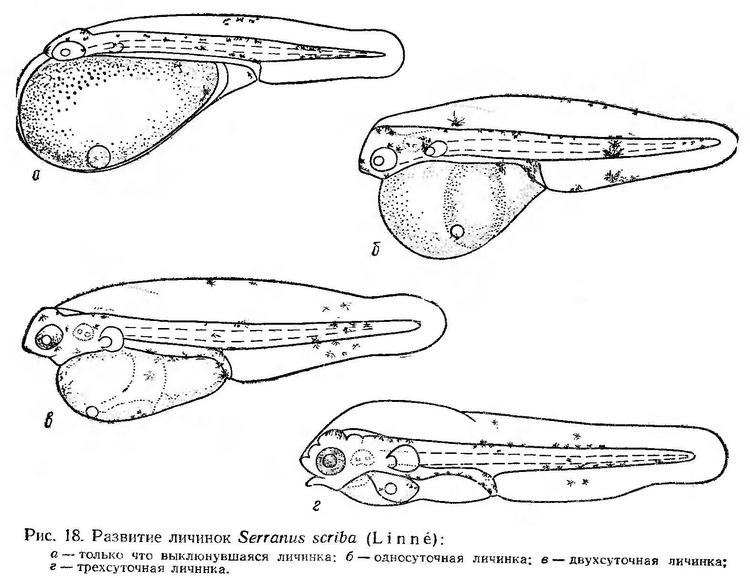
Постэмбриональное развитие. Длина только что выклюнувшихся личинок 1,9—2,3 мм. Она имеет большой яйцевидный желточный мешок, выступающий перед головой. Жировая капля расположена или у переднего нижнего края желточного мешка, или почти у его середины. Голова плотно прижата к желточному мешку. Анус открывается в край плавниковой каймы за серединой тела; антеанальное расстояние составляет около 63% длины тела. Грудные плавники зачаточные. Распределение пигмента такое же, как у эмбрионов перед выклевом. Кроме этого, клетки желтого и черного пигмента появляются в средней части спинной плавниковой каймы по ее верхнему краю и на анальном сосочке (рис. 18, а). На жировой капле желтый и черный пигмент имеется не всегда. По наблюдениям Л. П. Салеховой (1963), у только что выклюнувшихся личинок Serranus scriba на плавниковой кайме пигмента нет, в области жировой капли имеются звездчатые серо-желтые пигментные клетки. Л. С. Овен (1959) отмечает наличие меланофоров у личинок Serranus scriba и на плавниковой кайме, и на жировой капле. Интересно, что поданным Рафаэле (1888) у только что выклюнувшихся личинок этого вида имеется пигмент на плавниковой кайме и на жировой капле.
Только что выклюнувшиеся личинки плавают под углом к поверхности воды брюшной стороной и головой вверх. Продолжительное время (до 2 мин) личинки неподвижно парят в воде, затем совершают быстрые короткие броски. При этом личинки принимают горизонтальное положение, но не переворачиваются, а, как и в покое, ориентированы брюшной стороной кверху.
У односуточных личинок намечается ротовая ямка, увеличиваются грудные плавники. Антеанальное расстояние сокращается до 52—53% длины тела за счет удлинения хвостового отдела. В передней части дорсальной плавниковой каймы образуется оводненная полость. Усиливается пигментация тела. Появляется пигмент на нижней стороне кишечника и на вентральной плавниковой кайме (рис. 18, б). Личинки становятся более подвижными.
На вторые сутки развития намечается ротовая щель, начинается пигментация глаз. Черный пигмент перемещается на вентральную сторону. Усиливается меланинная пигментация на нижней стороне кишечника. Личинки при движении на короткое время переворачиваются спиной вверх; в покое они по-прежнему обращены брюшной стороной кверху.
Двухсуточные личинки в наших опытах достигали 2,40 — 2,55 мм в длину. Анус расположен на уровне середины тела. Рот прорезался. Грудные плавники приобрели подвижность. Оводненная полость увеличилась (рис. 18, в). Личинки длительное время находятся в движении; периоды покоя сокращаются до 30—50 сек.
По наблюдениям Л. П. Салеховой (1963), у трехсуточных личинок сохраняется небольшой остаток желточного мешка и жировая капля. Анус занимает более переднее положение; антеанальное расстояние составляет 46% длины тела. Глаза интенсивно пигментированы. С дорсальной части плавниковой каймы пигментные клетки исчезают. Значительно меньше их становится в постанальном отделе тела, особенно на вентральной части плавниковой каймы (рис. 18, г по Л. П. Салеховой, 1963).
Распределение и численность икринок и личинок. Икринки каменного окуня обнаруживаются в планктоне у Крымского и Кавказского побережий обычно в небольшом количестве в поверхностных слоях воды. В июне икринки Serranus scriba встречаются редко единичными экземплярами. Наиболее часто они ловятся в июле. В отдельных случаях их численность в это время достигает 130—150 экз/м2 и в среднем колеблется в разные годы от 4 до 18 экз/м2. В августе количество икринок Serranus scriba в планктоне обычно меньше, чем в июле: не более 20 экз/м2 и в среднем от 1 до 6 экз/м2. В открытом море в значительном удалении от берегов икринки каменного окуня не встречались.
Нахождение личинок Serranus scriba в Черном море в литературе не отмечено. Указание В. А. Водяницкого (1930а) на нахождение в планктоне личинки этого вида, судя по приведенному описанию, по-видимому, ошибочно.
За весь период наших наблюдений в районе Севастополя было выловлено 18 личинок каменного окуня размером от 1,5 до 3,4 мм. По горизонтам лова они распределяются следующим образом: на 0 м выловлено 6 личинок, на 10 м — 7 личинок, в слое 25—0 м — 5 личинок.
В Средиземном море личинки и мальки этого вида были собраны Датской океанографической экспедицией в поверхностном слое на двух станциях в районе Гибралтарского пролива и на одной станции в Критском море (Fage, 1918).
По нашим данным (Дехник, Синюкова, 1964), личинки каменного окуня встречались в планктоне в разных районах Средиземного моря, преимущественно в Критском и Адриатическом морях. Численность их в улове обычно составляла 1—2 экз., и только на одной станции в проливе Отранто была выловлена 21 личинка, 12 из этого количества были взяты с горизонта 25 м.
Вертикальное распределение личинок Serranus scriba в Средиземном море по нашим данным ограничено преимущественно слоем 25—50 м. Из большого числа ловов в верхних горизонтах (до 25 м) личинки Serranus scriba обнаружены только в одном лове (4 экз.). С нижних горизонтов (ниже 50 м) личинки вылавливались очень редко и могут рассматриваться как прилов из вышележащих слоев (Дехник, Синюкова, 1964).
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
В восточной части Атлантического
океана от Португалии до Анголы, в
Средиземном и Черном морях обитает каменный
окунь (Serranus scriba). Основным
отличием этого вида от описанных является
наличие на жаберной крышке трех
довольно сильных шипов. Это относительно
небольшие рыбы, длиной до 30 см,
живущие преимущественно в прибрежных
камнях, имеют бурую окраску с несколькими
темными поперечными полосами на боках.
На непарных плавниках несколько рядов
темно-красных пятен. Самой интересной
особенностью этой рыбы является то, что
она гермафродит. Половые железы
одной и той же рыбы содержат как икру,
так и молоки. Созревание икры и молок
обычно происходит поочередно, таким
образом, одна и та же особь может
участвовать в нересте то как самец, то как
самка. Иногда происходит одновременное
созревание икры и молок, в результате чего
возможно оплодотворение икры молоками
той же рыбы. Каменный окунь
сравнительно малочисленная рыба и в
незначительных количествах добывается
крючковыми орудиями лова.