Trachinus draco Linnaeus, 1758
Большой дракончик, Большой морской дракончик, Большой морской дракон, Морской дракончик, Морской дракон, Большой морской скорпион, Змейка, Драконовый трахинус, Трахинус драко (Rus),
Greater weever, Greater weever fish, Common weever, Stingfish
(Eng)
Синонимы:
Echiichthys draco (Linnaeus, 1758)
Trachinus draco Linnaeus, 1758
Trachinus lineatus Bloch & Schneider, 1801
Атлас-определитель рыб: Книга для учащихся. Н.А.Мягков 1994 г.
Дракончик морской — Trachinus draco (Linnaeus, 1758)
Тело покрыто мелкой чешуей. У переднего края орбиты глаза лежат 2—3 крупных косых шипа, направленных назад и вверх. Первый спинной плавник с мощными колючками и коротким основанием. D1 VI —VII. D2 28—31. А II 27—31. В срединном ряду 77—85 чешуек. Окраска тела серовато-желтая с короткими косыми поперечными полосами коричневого цвета. Большая часть первого спинного плавника с черным пятном. Достигает длины тела до 35 см (обычно до 25 см). Обитает на дне в прибрежной зоне Черного моря.
Внимание. Уколы колючек зарывшегося в песок дракончика очень болезненны, так как лучи первого спинного плавника и заглазничные шипы рыбы связаны с железой, вырабатывающей довольно сильный яд.
Иллюстрированная энциклопедия рыб. Ст.Франк 1983 г.
В Атлантическом океане и Средиземном море живет большой дракончик (Truchinus draco), длиной до 40 см. Его ядовитые железы содержат очень сильный яд, весьма опасный и для человека. Икра пелагическая. Рот относительно велик и направлен косо вверх. Морские дракончики подстерегают добычу, до глаз зарывшись в песок.
Ихтиопланктон Черного моря. Т.В.Дехник 1973 г.
МОРСКОЙ ДРАКОН — TRACHINUS DRACO (LINNE)
Условия размножения. Первые сведения о размножении морского дракона в Черном море приводит С. А. Зернов (1913). По его данным, производители со зрелыми, готовыми к вымету половыми продуктами встречались в июне и июле. У берегов Румынии нерест был отмечен в конце мая, июне и июле (Borcea, 1933). По мере дальнейших исследований представления о размножении Trachinus draco в Черном море уточнялись. Имеющиеся в настоящее время данные свидетельствуют о том, что период размножения этого вида в Черном море значительно растянут. Нерест начинается в конце мая — начале июня и продолжается до середины октября (Borcea, 1927, 1933; Водяницкий, 1930а; Косякина, 1938; Пчелина, 1940; Виноградов, 1948, 1949; Дехник и Павловская, 1950; Водяницкий и Казанова, 1954; Зайцев, 1959а; Овен, 1959; Смирнов, 1959; Дука, 1959; Георгиев и др., 1960). Разгар нереста приходится на наиболее теплое время года (июль — август).
В период размножения морской дракон держится в основном у берегов на небольших глубинах. Икринки встречаются в планктоне преимущественно в трех—пятимильной прибрежной зоне (Дехник и Павловская, 1950; Овен, 1959; Георгиев и др., 1960) и в меньшем количестве до 10 миль от берега (Овен, 1959; Георгиев и др., 1960; наши наблюдения).
Температура воды в местах нахождения икринок и личинок колеблется от 18 до 25°. Ю. П. Зайцев (1959а) отмечает нахождение икринок при температуре 12—12,5°. Оптимальный температурный режим для развития икринок и личинок, соответственно разгару нереста, наблюдающемуся, как отмечено, в июле и августе, создается при прогреве воды до 22—24°.
Режим солености Черного моря вполне благоприятен для нормального развития икры и личинок морского дракона. Судя по очень редкой встречаемости икры и личинок Trachinus draco в опресненном Одесском заливе (Зайцев, 1959а), можно считать, что размножение этого вида в Черном море ограничено уровнем средней солености (в Одесском заливе единичные икринки были обнаружены при солености 17,70 — 17,96‰). Однако несколько развивающихся икринок морского дракона было найдено нами в Азовском море при солености 11,55‰.
В Средиземном море Trachinus draco размножается весной и летом. В это время в планктоне встречаются икринки и в уловах зрелые самцы и самки (Padoa, 1956д; Lee, 1966). В Северном море нерест происходит с июня по август (Hoek; цит. по Padoa, 1956д).
Таким образом, размножение Trachinus draco в разных районах ареала приурочено к теплому периоду года. В Черном и Северном морях интенсивный нерест происходит в разгар лета, т. е. при наиболее высокой для этих районов температуре воды. В Средиземном море, соответственно более раннему прогреву воды, начало нереста сдвинуто на весенний период, а разгар его приходится на первую половину лета.
Соленость в районах размножения Trachinus draco, соответственно ареалу, колеблется в значительных пределах. Икринки нормально развиваются во взвешенном состоянии при солености 17—18‰ в Черном море, при средней океанической солености в Северном море и при более высокой солености Средиземного моря (38—39‰). Данные о размножении этого вида в Азовском море нуждаются в подтверждении.
О суточном ритме размножения. Данные по нахождению икринок на разных этапах развития в течение суток не свидетельствуют о строгой периодичности их поступления в планктон (табл. 30).
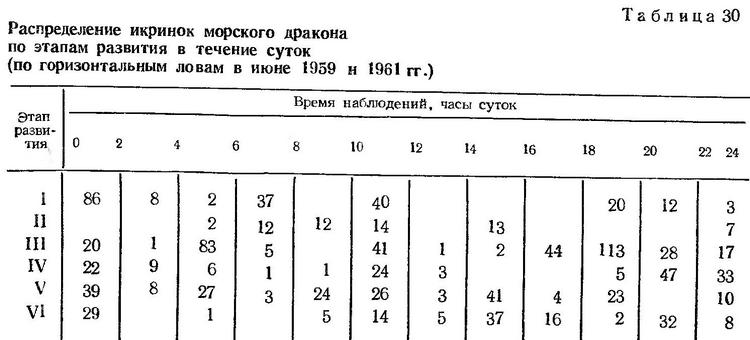
Икринки на этапе дробления встречаются в планктоне в разные часы суток и не отмечены в уловах лишь в небольшой период времени (от 12 до 18 ч). Икринки на III, IV и V, VI этапах вылавливаются круглосуточно. Одновременно в уловах могут быть обнаружены икринки на всех шести этапах развития.
Следовательно, в отличие от других видов с четко выраженным суточным ритмом размножения, нерест Trachinus draco не приурочен к какому-либо определенному времени, а происходит в разное время суток. Чем объясняется отсутствие суточной периодичности размножения у этого вида, сказать в настоящее время трудно.
Строение икринок. Икринки Trachinus draco пелагические, сферической формы, с одной жировой каплей. Оболочка прозрачная, но сравнительно плотная и упругая, отличается ячеистой (пористой) структурой поверхности. Желток гомогенный (рис. 38).
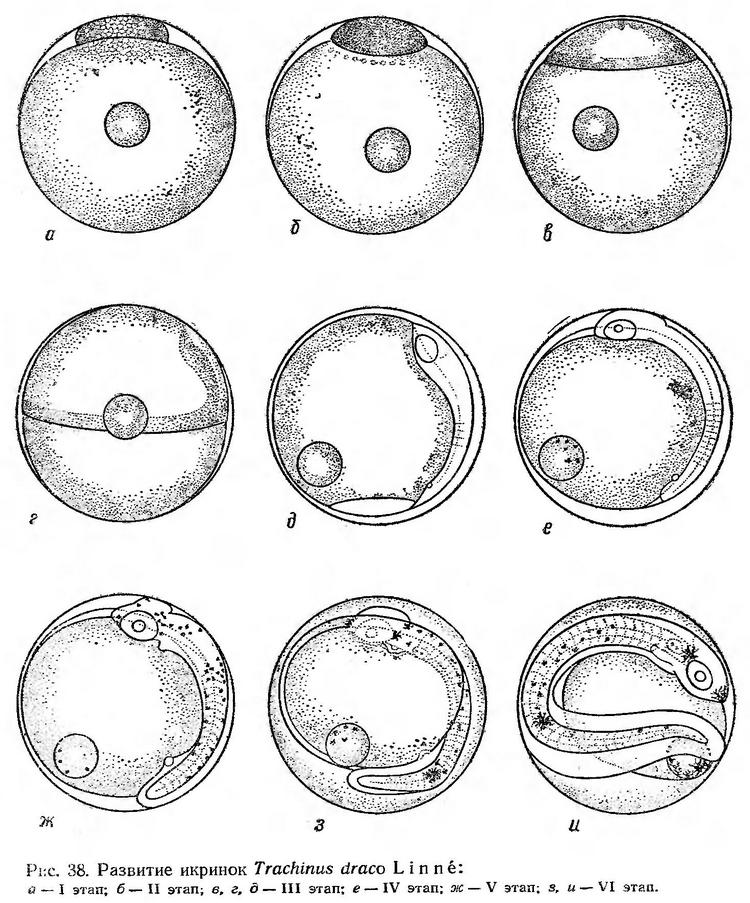
Размеры икринок морского дракона, по данным различных исследователей, колеблются от 0,94 до 1,15 мм, жировой капли — от 0,19 до 0,25 мм (Водяницкий и Казанова, 1954; Зайцев, 1959а; Георгиев и др., 1960; наши измерения). Размер икринок этого вида из Северного моря — 0,94—1,11 мм, жировой капли — 0,19—0,23 мм (Padoa, 1956д).
Эмбриональное развитие морского дракона прослежено на икринках, выловленных в море.
Бластодиск развивающихся икринок составляет примерно десятую часть по отношению к диаметру желтка. Его высота равна 0,10 мм, длина основания — 0,50 мм (рис. 38 а, табл. 31). По мере формирования эпителиальной бластулы бластодиск немного уплощается, в перибласте появляются пузыревидные образования, которые постепенно распространяются по поверхности желтка (рис. 38,6). Эпителиальная бластула при температуре воды 22° формируется в течение 3 ч.
Обрастание желтка и гаструляция начинаются быстрым увеличением основания бластодиска. У одного его края появляется зародышевый узелок (рис. 38, в). Когда края бластодермы доходят до середины желтка, зародышевая полоска принимает довольно четкие очертания. Головной конец заметно расширен и почти достигает анимального полюса (рис. 38, г). В конце III этапа развития перед замыканием бластопора намечаются зачатки глаз, появляются первые туловищные сегменты (рис. 38, д). III этап при температуре 22—20,6° продолжался около 9 ч.
После замыкания бластопора число туловищных сегментов быстро увеличивается; намечаются зачатки внутренних органов, слуховых капсул, в глазах обозначены хрусталики. На жировой капле появляются меланофоры; на теле пигмента нет (рис. 38, е).
На V этапе наряду с ростом хвостового отдела дифференцируются хвостовые сегменты. Развиваются зачатки внутренних органов. Формируется сердце, закладывается кишечная трубка, намечается плавниковая кайма. На теле появляются мелкие точечные меланофоры (рис. 38, ж).
Через 7 ч после появления хвостовой почки при температуре 20—20,3° начинает пульсировать сердце, эмбрион слабо подергивается. К этому времени тело эмбриона охватывает около 2/3 поверхности желтка. Сердце пульсирует 80—90 раз в минуту. Меланинная пигментация усиливается. Меланофоры густо рассеяны по всему телу. На рыле, за глазами и в хвостовом отделе имеются пятна ярко-желтого пигмента, которые в проходящем свете приобретают черную окраску (рис. 38, з). Перед выклевом хвост эмбриона смыкается с головой, желточный мешок уменьшается. Эмбрион энергично двигается (рис. 38, и).
Этап подвижного эмбриона у морского дракона значительно растянут во времени по сравнению с другими размножающимися летом рыбами с пелагической икрой. При температуре воды 20—20,6° он продолжается 38—40 ч, при температуре 22—23° — 32—34 ч.
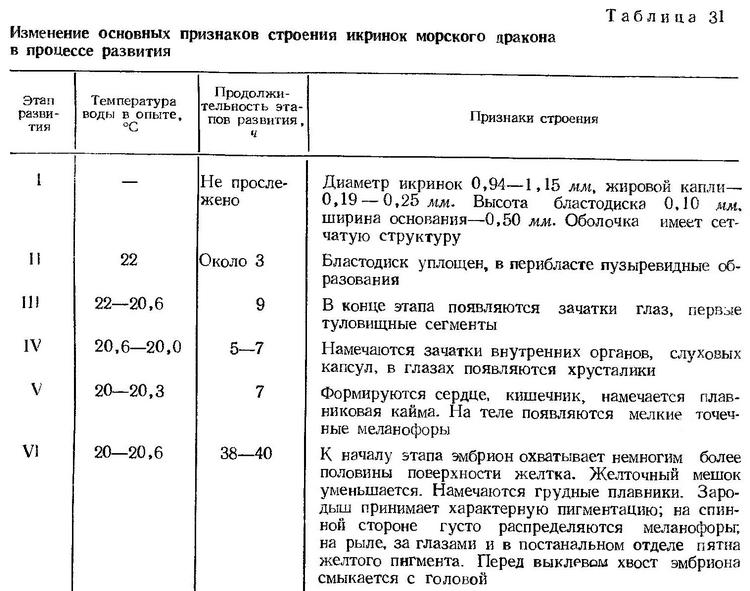
Постэмбриональное развитие. В связи с длительным периодом эмбрионального развития и особенно шестого этапа личинки морского дракона выклевываются сравнительно хорошо развитыми. Длина выклюнувшихся личинок 2,70—2,95 мм (по Зайцеву, 1959а, — 2,35—2,48 мм). Голова не соединена с желточным мешком, и уже обозначена ротовая ямка. Тело стройное, прогонистое, с довольно коротким туловищем и более длинным хвостовым отделом. Антеанальное расстояние составляет 41—43% длины тела. Жировая капля расположена в нижней передней части желточного мешка. Грудные плавники в виде небольших зачатков. Анус открывается в край плавниковой каймы. Сердце пульсирует очень быстро — 130—140 биений в минуту. Пигментация тела характерная. За глазами и в средней части хвостового отдела на брюшной стороне расположены крупные желтые пигментные клетки, причем в хвостовом отделе имеются одна или две такие клетки. На спинной стороне от рыла вдоль всего туловища и передней части хвостового отдела расположены ветвистые меланофоры. Ряд черных пигментных клеток имеется на брюшной стороне вдоль всего хвостового отдела. Поверхность жировой капли также пигментирована меланином (рис. 39, а).
Только что выклюнувшиеся личинки плавают в горизонтальном положении брюшной стороной вверх. Паузы между перемещениями длительные — от 25—30 сек до 2 мин. Моменты движения продолжаются не более 2—3 сек. Личинки двигаются или короткими бросками в горизонтальной плоскости, не меняя положения, или винтообразно вверх, быстро переходя в состояние покоя и принимая прежнее перевернутое положение.
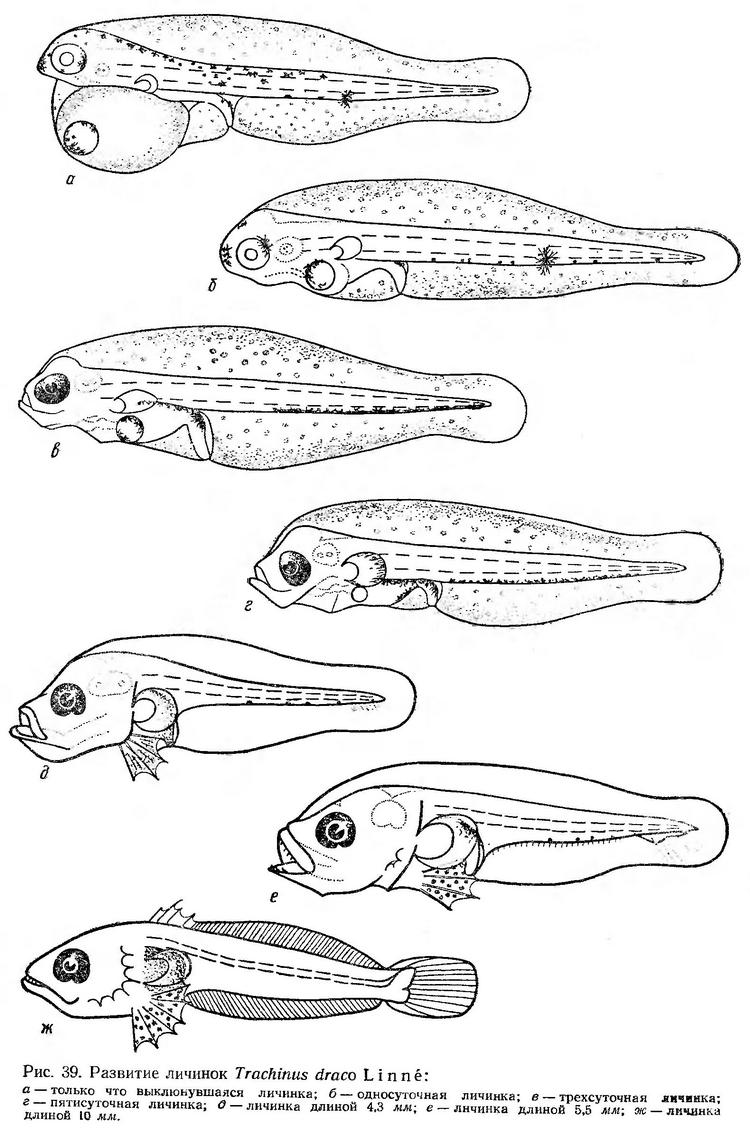
В течение первых суток развития желточный мешок сильно сокращается. Голова полностью освобождается от него. Грудные плавники увеличиваются, они широко расставлены в стороны, но еще неподвижны. Хвостовой отдел значительно удлиняется, благодаря чему личинки начинают двигаться быстрее, стремительнее. Меланинный пигмент перемещается полностью на брюшную сторону. На спине у некоторых личинок сохраняются 1—2 меланофора в туловищном или в хвостовом отделе. За глазами и на брюшной стороне в средней части хвостового отдела сохраняются крупные желтые пятна. На рыле имеется скопление черного пигмента. В спинной части плавниковой каймы образовалась большая, заполненная водой полость (рис. 39, б). Движения личинок по-прежнему в основном в форме быстрых стремительных скачков, после которых наступает длительный покой (до 1,5—2 мин).
У двухсуточных личинок остается небольшой желточный мешок, а размер жировой капли почти не изменяется. Личинки увеличиваются в длину до 3,1—3,2 мм. Рот открыт, но еще неподвижен. В глазах появился пигмент. Грудные плавники широко расставлены, при движении быстро вибрируют, в покое неподвижны. Пигмент только на брюшной стороне и на жировой капле. Желтые клетки исчезают.
Личинки начинают плавать в горизонтальном положении — спиной кверху или чуть под углом головой вниз. Периоды покоя сократились до 5—30 сек. Движения довольно плавные и время от времени стремительные, скачкообразные, при этом личинки преодолевают расстояние в 1—1,5 см.
В возрасте трех суток сохраняется очень маленький остаток желтка; жировая капля уменьшается. Рот приобретает подвижность. Меланинная пигментация, как у двухсуточных личинок: ветвистые пигментные клетки расположены вдоль всей брюшной стороны тела. У основания анального сосочка имеется крупный меланофор (рис. 39, в). Личинки очень подвижны. Паузы короткие; моменты движения — 2—3 сек. Время от времени личинки совершают стремительные броски на значительные расстояния.
К пятисуточному возрасту желточный мешок резорбируется, но еще сохраняется небольшая жировая капля (рис. 39, г). Личинки плавают в разных направлениях с помощью быстрых вибрирующих изгибов хвостового отдела и движения плавников.
Дальнейшее описание дано по фиксированному материалу.
Личинки длиной 4,3 мм уже хорошо развиты. Голова у них становится массивной; туловищный отдел значительно укорачивается. Впереди грудных развиваются большие брюшные плавники. Нижняя челюсть слегка выдается за верхнюю, которая к этому времени уже может выдвигаться. Начинается образование опорных элементов нижней лопасти хвостового плавника (рис. 39, д). По наблюдениям В. И. Синюковой (неопубликованные данные), личинки таких размеров питаются науплиусами Copepoda (49,6% по весу и 84,7% по количеству ) и взрослыми формами Oithona minuta (50,4% по весу и 15,3% по количеству).
У личинок длиной 5,5 мм появляются острые зубы на челюстях, на предкрышках образуются шипы, продолжается образование скелета хвостового плавника (рис. 39, е). При длине около 10 мм заканчивается формирование всех непарных плавников. Тело личинок удлиненное, сжатое с боков. Челюсти снабжены большими острыми зубами (рис. 39, ж).
Распределение и численность икринок и личинок. Икринки и личинки морского дракона встречаются в летнее время вдоль всего побережья Крыма и Кавказа, в северо-западной части моря, у побережья Болгарии.
В Новороссийской бухте икринки вылавливались обычно в небольшом количестве (Водяницкий, 1930а; Косякина, 1938); в Одесском заливе и у берегов Болгарии — также единичными экземплярами (Зайцев, 1959а; Георгиев и др., 1960). В значительном количестве отмечено их нахождение в Каркинитском заливе (Павловская, 1950), у Карадага (Овен, 1959), в районе Туапсе — Сухуми (Дехник и Павловская, 1950), в Севастопольской бухте (Дука, 1959). В районе Севастополя икринки и личинки морского дракона — постоянные компоненты ихтиопланктона в течение всего лета и первой половины осени.

Икринки распределяются во всем теплом слое моря (табл. 32). В уловах с глубины 15 и 20 м (Евпаторийский район, август 1957 г. и прибрежные районы Кавказа, август 1965 г.) насчитывалось до 30 икринок этого вида. Численность икринок в уловах значительно колеблется как в течение нерестового сезона, так и в разные годы наблюдений. В июне они встречаются обычно единичными экземплярами. В июле и августе в районе Севастополя численность икринок достигает в отдельных случаях 80—120 экз., а в среднем не более 7—20 экз/м2. Икринки морского дракона продолжают встречаться в значительном количестве в сентябре и первой половине октября (до 30 экз/м2).
Личинки вылавливаются обычно единичными экземплярами. Только в один период — в августе 1961 г. в районе Севастополя было отмечено 8 личинок этого вида под 1 м2 поверхности моря.
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
Большой дракончик (Trachinus draco)
по виду очень похож на малого
дракончика, но крупнее его, длина 30—45 см,
окраска более темная с косыми полосами
на боках. Плавники желтые с черными
краями; первый спинной плавник черный.
Большой дракончик живет в более
глубоких местах, иногда попадается сотнями.
Область распространения его — от
Балтийского моря до Черного. Половозрелости достигает в 3-летнем возрасте.
Держится на дне, но иногда всплывает к
поверхности. В Черном море нерест с июня
до октября.