Bothrocara hollandi (Jordan & Hubbs, 1925)
Синонимы:
Allolepis hollandi Jordan & Hubbs, 1925
Allolepis nazumii Mori, 1956
Lycogramma crystallonota Popov, 1933
Рыбы Японского моря и сопредельных частей Охотского и Желтого морей. Часть 4. (Perciformes). Г.У.Линдберг, З.В.Красюкова 1975 г.
1. Allolepis hollandi Jordan et Hubbs, 1925 (рис. 144).
Allolepis hollandi Jordan et Hubbs, Mem. Carnegie Mus., 1925 : 323, pl. 12, fig. 2 (Фукуи). — Таранец, Краткий определитель..., 1937 : 165. — Линдберг, Предварительный список..., 1947 : 171. — Matsubara, Fish morphol. a. hierar., 1955 : 782. — Fowler, Synopsis..., 1958 : 317, fig. 41. — Линдберг и др., Исслед. дальневост. морей СССР, 6, 2, 1959 : 251.
Lycogramma crystallonota Schmidt, в: Попов, Исслед. морей СССР, 1933 : 151.
24499. Японское море. 4 X 1931. Д. И. Охрямкин. 3 экз.
32227. Японское море. 1933. Н. М. Сомова. 1 экз.
33081. Охотское море. 27 VIII 1949. П. Ю. Шмидт. 3 экз.
33091. Японское море. 10 VIII 1933. Н. Спасский. 2 экз.
37967. Японское море, зал. Петра Великого. 1 XI 1965. И. И. Серобаба. 3 экз.
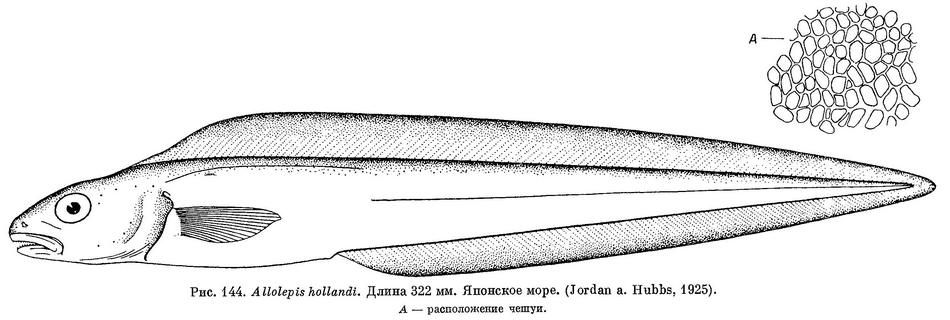
D 115; Р 17. Длина головы и туловища 1.95 раза в длине тела до основания хвостового плавника; голова 6.15; высота 9.0 раза в той же длине. Очень характерно расположение чешуи на теле (см. рис. 144, А). Цвет светлый, розовато-коричневый, темнее вдоль основания спинного плавника, на конце рыла и на крышечной кости. Вертикальные плавники по краю черноватые (Jordan a. Hubbs, 1925).
Просмотр наших экземпляров (длиной от 113 до 250 мм) и анализ рентгенограмм 9 экз. показали значительные вариации меристических и пластических признаков. Число позвонков варьирует от 115 до 125 (туловищных 17—19, хвостовых 97—107), число лучей спинного плавника от 110 до 122, анального от 90 до 110. Скелет свободной части хвостового плавника состоит из 8—10 основных и 2 краевых лучей. Антеанальное расстояние изменяется в пределах 32.8—36.6% абсолютной длины тела. Боковая линия выражена свободно сидящими невромастами, состоит из двух ветвей — верхней, которая начинается от верхнего конца жаберной щели и, поднимаясь вверх, проходит вблизи основания спинного плавника до вертикали конца первой четверти спинного плавника, и срединной ветви, начинающейся за вертикалью конца верхней ветви и проходящей по средней линии тела до хвостового плавника. (Боковая линия заметна не сразу, а требует внимательного рассмотрения, поэтому, наверно, Джордэн и Хаббс пишут, что она выражена неясно и редуцирована до неотчетливой полоски). Поры сейсмосенсорной системы головы представлены 8 подглазничными, 4 заглазничными, 1 предглазничной, 1 непарной межглазничной, 3 предкрышечными и 4 нижнечелюстными. Поры затылочного ряда нами не обнаружены (на фиксированном материале).
Длина 322 мм (Jordan а. Hubbs, 1925).
Распространение. В Японском море известен из зал. Петра Великого (наши экземпляры), из Воньсаня (Mori, 1952 : 131), в сев. части Татарского пролива (Таранец, 1937б : 165), у япономорского побережья Хоккайдо (Ueno, 1971 : 87), о. Садо (Honma, 1963 : 21), в зал. Тояма (Katayama, 1948 : 25), у побережья Фукуи (Jordan a. Hubbs, 1925 : 323), в зал. Вакаса (Takegawa a. Morino, 1970 : 383), в р-не Санин (Mori, 1956а : 22). В Охотском море у северо-восточного берега Сахалина (Шмидт, 1950 : 120) и в зал. Анива (Ueno, 1971 : 87).
2. Allolepis nazumi Mori, 1956.
Allolepis nazumi Mоri, Sci. Rep. Hyogo Univ. Agric., 2, 2, Nat. Sci., 1956 : 29 (банка Ямато, центральная часть Японского моря). — Fowler, Synopsis..., 1958 : 318.
(Так как нам не удалось ознакомиться с авторским описанием и изображением этого вида, мы приводим его характеристику по Фаулеру (Fowler, 1958) и условно вводим в состав рода как самостоятельный вид. Но, по-видимому, при особо внимательном рассмотрении этот вид придется считать лишь синонимом Allolepis hollandi.)
D 110; А 88—91; Р 14; жаберных тычинок 2—3 + 12, короткие. Боковая линия очень неотчетливая. Высота тела 7.67—8.1 раза в абсолютной длине тела; голова и тело сжаты с боков на всем протяжении. Рыло выдается вперед, 3.1 раза в длине головы; глаза 4.2—4.8 раза в этой же длине; верхнечелюстная кость достигает вертикали переднего края глаза. Верхняя челюсть длиннее нижней. Зубы на челюстях, сошнике и нёбных. Голова голая, за исключением части позади глаз, имеющей очень мелкую, округлую, погруженную в кожу чешую. Тело без чешуи на затылке, на передней половине брюха и полоски позади основания грудного плавника. Этой особенностью в основном и отличается от Allolepis hollandi Jordan et Hubbs (Fowler, 1958).
Длина 298 мм.
Распространение. В Японском море указан для банки Ямато в центральной части этого моря.
Рыбы северных морей СССР. А.П.Андрияшев 1954 г.
Чешуя у Anguilla anguilla (Linne) очень мелкая, удлиненной формы, образует группы, расположенные наподобие паркета. Замечательно, что еще более правильное паркетное строение имеет чешуйчатый покров Allolepis hollandi Jord. et Hubbs из семейства Zoarcidae (Lycogramminae) и Ophidium barbatum L. из Ophidiidae. Последняя рыбка зарывается в грунт совершенно особым способом: плывя над дном головой вперед, она внезапно меняет направление на обратное и, мгновенно втыкаясь задним концом тела в грунт, быстро погружает в него все тело. Не связано ли паркетное расположение чешуй с функцией движения «задним ходом» (хвостом вперед) в воде, между камней, в песке и т. д.?