Hippoglossoides elassodon Jordan & Gilbert, 1880
Узкозубая палтусовидная камбала, охотоморская палтусовидная камбала (Rus),
Flathead sole
(Eng)
Синонимы:
Hippoglossoides elassodon Jordan & Gilbert, 1880
Рыбы Японского моря и сопредельных частей Охотского и Желтого морей. Часть 6. (Pleuronectiformes). Г.У.Линдберг, В.В.Федоров 1993 г.
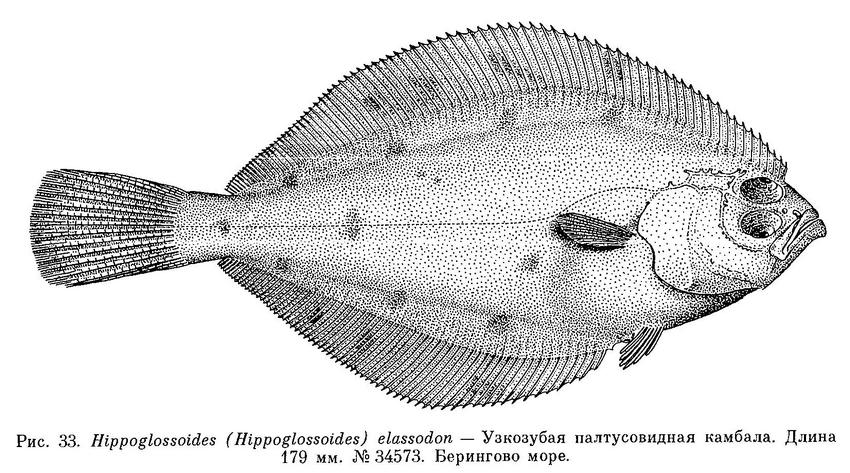
2. Hippoglossoides (Hippoglossoides) elassodon Jordan et Gilbert, 1881 — Узкозубая палтусовидная камбала (рис. 33, 34).
Hippoglossoides elassodon Jordan, Gilbert, Proc. U. S. Nat. Mus., 3, 1881b : 278, 454 (Сиэтл, Такома). — Шмидт, Рыбы Восточных морей..., 1904 : 225 (синонимия, описание). — Jordan, Starks, Proc. U. S. Nat. Mus., 31, 1906а : 189, fig. 8 (синонимия, описание). — Hubbs, Proc. U. S. Nat. Mus., 48, 1915 : 466 (описание). — Hubbs, Annot. Zool. Jap, 9, 4, 1918 : 369 (сравнение). — Norman, Flatfishes (Heterosomata)..., 1934 : 299, fig. 221 (синонимия, описание) . — Matsubara, Fish morph. a. hier, 2, 1955 : 1268. — Wilimovsky et al., Fish. Mar. Surv. Res. Dev., Tech. Rep, 34, 1967 : 87 (описание, отличие). — Alderdiсe, Forrester, J. Fish. Res. Bd. Can., 31, 1974 : 1899 (описание). — Forrester et al., J. Fish. Res. Bd. Can., 34, 4, 1977 : 455 (разделение рода на виды). — Богданов и др., в: «Генетика и размножение морских животных», 2, 1979 : 29 (видовая самостоятельность Hippoglossoides elassodon). — Коваль, Богданов, Вопр. ихтиол, 22, 4, 1982 : 679 (родовая систематика на основе генетических признаков).
Hippoglossoides elassodon elassodon, Schmidt, Ann. Mag. Nat. Hist., 8, 16, 1915 : 299 (описание, различия). — Солдатов, Линдберг, Обзор рыб..., 1930 : 395 (частью: исключить из синонимии Hippoglossoides katakurae Snyder). — Шмидт, Рыбы Охотского моря, 1950 : 227 (синонимия, описание). — Фадеев, Изв. ТИНРО, 102, 1978а : 17 (разделение рода на виды и подвиды).
17899, 17905. Японское море, Татарский прол., о. Сахалин. 26 VIII 1911. ДВЭ. 3+2 экз.
18533. Японское море, 50°58' с. ш., 141°04' в. д. 29 IV 1908. ДВЭ. 1 экз.
18538. Японское море, 50°45' с. ш., 141°55' в. д. Глубина 75 м. 17 VI 1910. Ф. А. Дербек. 3 экз.
18540. Японское море, 50°22' с. ш., 141°05' в. д. Глубина 100 м. 25 V 1912. ДВЭ. 3 экз.
48099. Японское море, Татарский прол. Глубина 139 м. 23 VIII 1949. КСЭ. 2 экз.
Охотское море: 12348, 12367, 13065, 13084, 18539, 45173, 45174, 45411.
Северные Курильские о-ва: 48091—48098, 48100, 48219, 48220, 48224, 48225.
Юго-Восточная Камчатка: 33492—33494, 34575, 34576, 42657, 43013, 44013, 45172.
Берингово море: 33495, 34508, 34573, 34574, 34583, 45175.
Зап. Аляска: 43003, 43704, 43987, 48226.
Калифорнийский зал.: 38516 (РТ «Адлер». 22 V 1965. Траление № 61, 29°10' с. ш., 115°14' з. д. Л. С. Кодолов. 1 экз.).
D 77—84; А 59—63; V 6; l.l. поры 88—92, чешуи 110—120; жаберных тычинок ? + 15—17; зубов 35+45 / 25+35; пилорических придатков 4 (Jordan, Gilbert, 1881b : 278).
D (76) 79—86; A 60—67 (69); P 10—12; С 18, ветвистых 12; жаберных тычинок ? + (14) 15—18 (19); l.l. 87—94, выше боковой линии 36—42 ряда чешуи (Norman, 1934 : 300).
D (70) 75—88; А (58) 60—68; l.l. 87—98; жаберных тычинок 4—5 + (15) 17—19 = (20) 21—24 (Kuronuma, 1943 : 115).
D 75—85; А 55—68; V 6; l.l. 86—91, выше боковой линии 40—50 рядов чешуи; жаберных тычинок ? + 16—24; позвонков 42—46 (Hart, 1973 : 612).
D 73—94; А 52—71; жаберных тычинок ? + 14—25 (Фадеев, 1978а : 17).
D 75—94; А 55—72; Р 9—12; V 6; l.l. 86—94; жаберных тычинок 4—5 + 14—24 (Masuda et al., 1984 : 336).
С 3 + 12 + 3 = 18; позвонков (12) 13 (14), ср. 12.9 + 32—33 (34), ср. 32.86 (Sakamoto, 1984 : 176, 186).
По 39 экз. длиной 33—266 мм коллекции ЗИН: D (76) 80—88, ср. 82.9; А (57) 61—69 (73), ср. 65.0; Р 10—11 (14), ср. 10.7 / 10—11 (13), ср. 10.7, С (2) 3 + 6 (7) + 6 + 3 = (17) 18 (19), ср. 18.1; l.l. (84) 86—93, ср. 89.3, рядов чешуи выше боковой линии (28) 32—34 (38), ср. 32.5, ниже боковой линии (30) 31—37 (41), ср. 35.1; жаберных тычинок (2) 3—5, ср. 4.1 + (15) 16—19 (20), ср. 17.4 = (18) 19—23 (25), ср. 21.4. В процентах стандартной длины: длина головы 27.2—30.1, ср. 28.8; наибольшая высота тела 32.5—53.3, ср. 42.7; наименьшая высота тела (хвостового стебля) 8.0—10.6, ср. 9.7; длина хвостового стебля 5.9—7.4, ср. 6.4; длина грудного плавника глазной стороны тела 10.3—15.5, ср. 12.8.
Единой точки зрения на таксономическое положение 3 форм палтусовидной камбалы не существует. В последние годы Вилимовски с соавторами (Wilimovsky et al., 1967 : 87) на основе анализа ограниченных морфологических материалов (только количество лучей в D и А) пришли к выводу о существовании двух видов: северного — Hippoglossoides robustus и широко распространенного — Hippoglossoides elassodon. Некоторые отличия были отмечены (но без таксономических выводов) у этих форм по икре и личинкам (Alderdice, Forrester, 1974 : 1899). Форрестер с соавторами (Forrester et al., 1977 : 455), используя электрофорез мышечного миогена и гемоглобина, а также морфологические данные рыб, собранных в Японском, Охотском и Беринговом морях, и применяя дискриминантный анализ морфологических данных, пришли к заключению о наличии 3 видов: Hippoglossoides elassodon, Hippoglossoides robustus и Hippoglossoides dubius. Н. С. Фадеев (1978а : 3) попытался изучить географическую изменчивость некоторых признаков (число лучей в D и А, число жаберных тычинок) по пробам на 45 станциях в северной части Тихого океана от зал. Петра Великого в Японском море на восток до штата Вашингтон. Анализируя степень статистических различий сначала по пробам, а потом по группам проб, и сопоставляя ее с данными по электрофорезу сыворотки крови, автор находит, что Hippoglossoides dubius является самостоятельным видов, а Hippoglossoides elassodon разделяется на 2 подвида: Hippoglossoides elassodon elassodon и Hippoglossoides elassodon robustus. Л. В. Богданов с соавторами (1979 : 155, 1981 : 29) и Коваль и Богданов (1982 : 685), используя электрофоретические данные сыворотных и мышечных белков, устанавливают, что Hippoglossoides dubius и Hippoglossoides robustus не обнаруживают качественных различий по белковым и ферментным системам и, скорее всего, являются конспецифичными формами (или подвидами) одного вида, а Hippoglossoides elassodon — самостоятельным видом. В последних своих работах Н. С. Фадеев (1984 : 82, 83, 1987 : 25, 29, 31) принимает все 3 формы в ранге самостоятельных видов.
Узкозубая палтусовидная камбала обитает на шельфе и материковом склоне на глубинах от 6 до 1050 м, максимальная встречаемость — от 80 до 350 м. Зимой она, освобождая верхние части шельфа, перемещается на большие глубины, где образует локальные малоподвижные скопления, и распределяется в водах теплого промежуточного слоя с температурой 2—4 °С. Весной концентрации ее рассредоточиваются на большой площади и располагаются на глубинах 30—350 м в слое остаточного зимнего охлаждения при температуре воды от —0.4 до +3.0 °С. Нагул проходит в основном в тех же районах шельфа, где до этого был нерест, в водах, прогретых до температуры 0.5—4.5 °С. В промысловых уловах преобладают рыбы длиной 29—41 см, массой 0.23—0.75 кг. Половое созревание наступает при достижении у самцов длины 18—37 см и возраста 6—9 лет, у самок соответственно 18—39 см и 7—8 лет. Нерест отмечен на глубинах 20—100 м при температуре воды у дна от —0.4 до +2.0 °С. Сроки нереста: у берегов о. Хоккайдо в Охотском море в феврале-апреле, у берегов Камчатки как в Охотском море, так и в Тихом океане в апреле-июне, у северных Курильских о-вов в июне, в северной части Берингова моря в мае-июле, в южной части Берингова моря в феврале-мае. Икра мелкая, 0.3—1.1 мм в диаметре, пелагическая, развивается при температуре воды в поверхностных слоях от —1.2 до +5.4 °С. Плодовитость 35.5—100.1 тыс. икринок. Предличинка выклевывается при длине 5.3—6.9 мм. Метаморфоз начинается при достижении длины 10.7 мм и заканчивается при переходе к донному образу жизни при длине 28 мм. В пище наблюдаются как бентосные (черви, креветки, двустворчатые моллюски, офиуры), так и рыбы, и планктонные животные (гипереиды, эвфаузиевые, сагитты). Состав пищи сильно изменяется в зависимости от района сбора материалов и характера рельефа дна и грунтов. Зимой интенсивность питания резко снижается, число пустых желудков достигает 50 %, рыбы и их скопления малоподвижны. Обзор биологии этого вида приведен в работах как отечественных (Расс, Желтенкова, 1948; Моисеев, 1953а; Мусиенко, 1954, 1970; Перцева-Остроумова, 1961; Скалкин, 1963; Фадеев, 1970е, 1971а, 1971б, 1984, 1987; Николотова, 1972, 1975а, 1975б; Табунков, 1979; Чернышева, Швецов, 1979; Булатов, 1982), так и зарубежных (Suychiro, 1934; Takeuchi, Imai, 1959; Forrester, Alderdice, 1968; Forrester et al., 1977) исследователей.
В последние годы в связи с обнаружением у Hippoglossoides elassodon эпидермальной папилемы вирусной природы (Wellings et al., 1963, 1964, 1965, 1967; Wellings, Chuinard, 1964) появился интерес к изучению строения кожи и чешуи (Brown, Wellings, 1969, 1970; Willings, 1969а, 1969b; Willings, Brown, 1989).
Имеет промысловое значение для Дальнего Востока. Промысловые уловы по видам не разбираются и статистические данные носят сборный характер. В районах обитания нескольких видов палтусовидных камбал доля их в общих уловах камбаловых составляет в среднем за год в северной части Татарского прол. 17.2 %, вылов — 240 т, соответственно у Юго-Западного Сахалина — 7.6 % и 600 т, у Западной Камчатки — 3.9 % и 30 т, в восточной части Берингова моря — 3.4 % и 3200 т.
Рыбаки Японии вылавливали в Беринговом море в среднем больше наших рыбаков: 13.2 % и 18 300 т (Иванков, Иванкова, 1974; Фадеев, 1971б, 1978а, 1987; Alderdice, Forrester, 1974).
Длина 560 мм, масса 1.6 кг (Фадеев, 1987 : 115).
Распространение. В Японском море известен из северной части Татарского прол. у берегов о. Сахалин, где отмечался очень редко (№№ 17899, 17905, 18533, 18538, 18540, 48099). Указание на воды Приморья, Юго-Западного Сахалина и о. Хоккайдо к северу от мыса Сякотан (Ueno, 1971 : 99), на воды у тихоокеанских берегов о-вов Хонсю и Хоккайдо (Фадеев, 1987 : 31) сомнительно, по-видимому, это Hippoglossoides dubius. В Охотском море встречается от зал. Анива на север вдоль восточных берегов о. Сахалин до о. Св. Ионы (Линдберг, 1959 : 526; Ueno, 1971 : 99), у Западаной Камчатки (Моисеев, 1953а : 147) и в южной части моря (Ueno, 1971 : 99). У Курильских о-вов к северу от скал Ловушки (Перцева-Остроумова, 1961 : 54). Восточная Камчатка. Берингово море, зал. Аляска, Британская Колумбия, Вашингтон, Орегон и Северная Калифорния на юг до мыса Рейс (Hart, 1973 : 613; Eschmeyer et al., 1983 : 286) и даже в Калифорнийском зал. (№ 38516). Южная часть Чукотского моря (Quast, Hall, 1972 : 37).
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
Различают четыре вида. Три вида собственно
палтусовидных камбал — охотоморская
(Hippoglossoides elassodon), япономорская (Hippoglossoides dubius)
и северная (Hippoglossoides robustus) — живут в морях
Тихого океана.
Держатся они, в
особенности охотоморская, на
сравнительно больших глубинах.
Рыбы северных морей СССР. А.П.Андрияшев 1954 г.
Hippoglossoides robustus Gill et Townsend отличается от распространенного южнее типичного Hippoglossoides elassodon Jord. et Gilb. меньшим числом лучей в D и А, меньшим числом тычинок на нижней части 1-й жаберной дуги (9—13 вместо 15—19), большим верхним глазом, более удлиненным телом, более широким межглазничным промежутком, покрытым двумя рядами чешуй, пятнистой окраской молоди и др. (Андрияшев, 1937 : 324).