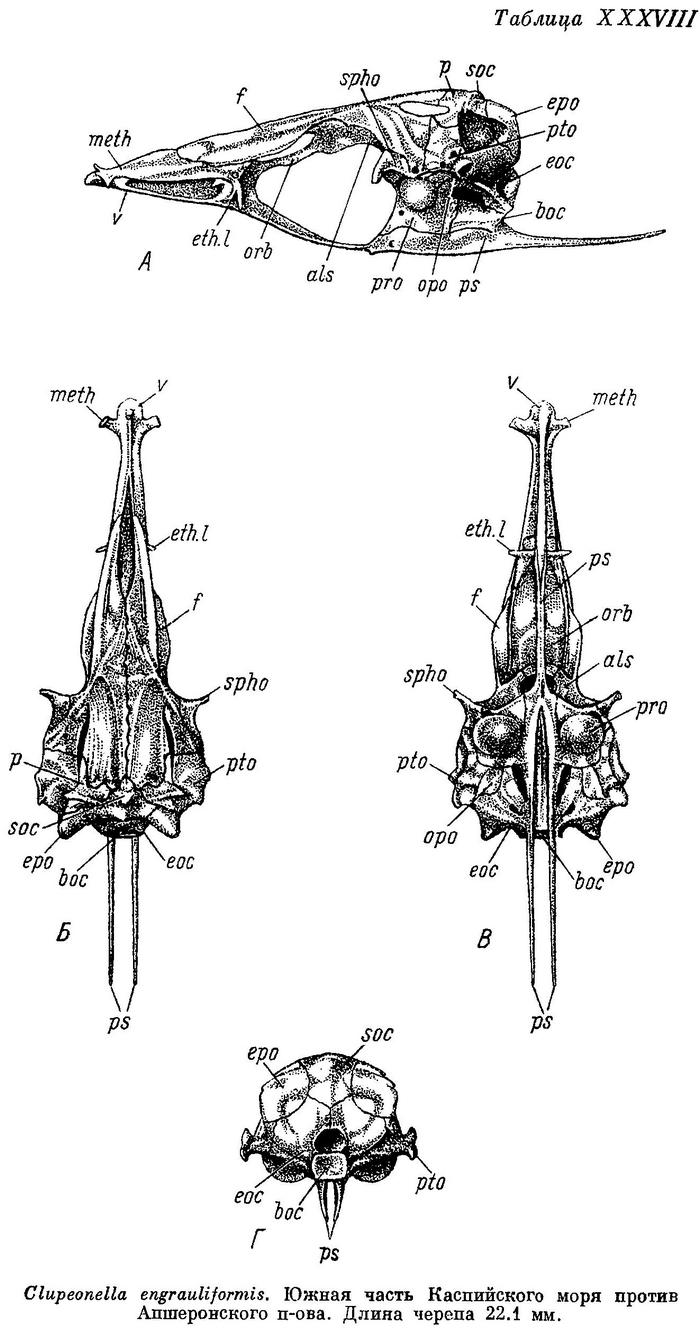
Clupeonella engrauliformis (Borodin, 1904)
Анчоусовидная тюлька, анчоусовидная сарделька, анчоусовидная килька (Rus),
Anchovy sprat
(Eng)
Синонимы:
Clupea engrauliformis Borodin, 1904
Harengula engrauliformis Berg, 1913
Harengula grimmi (non Kessler) Berg, 1923
Атлас-определитель рыб: Книга для учащихся. Н.А.Мягков 1994 г.
Килька анчоусовидная — Clupeonella engrauliformis (Borodin, 1904)
Тело вальковатое, весьма низкое, в поперечном сечении округлое. Достигает длины 15 см. Типично морской вид, не выносит солености ниже 8%. Предпочитает сравнительно большие глубины (свыше 10 м) средней и южной частей Каспийского моря. Объект промышленного лова.
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
Анчоусовидная килька (Clupeonella engrauliformis) обитает в Среднем и Южном Каспии,
заходя и в южную часть Северного
Каспия. В отличие от обыкновенной кильки
никогда не встречается при солености
ниже 8‰, будучи обитательницей
открытых районов моря и избегая глубины
менее 10 м. Анчоусовидная килька имеет
более прогонистое тело, чем
обыкновенная каспийская килька, живет до 7 лет
и растет быстрее. Достигает длины до
15,5 см, обычная длина до 11,5—12,5 см.
Позвонков у нее 44—48. Зимой
анчоусовидная килька держится главным
образом в Южном Каспии, преимущественно
в области глубин от 50 до 750 м. Весной
и летом идет на север и сосредоточивается
в огромном количестве в Среднем Каспии,
придерживаясь зоны температурного
скачка на глубине от 15 до 60 м. Нерестится
главным образом в августе — октябре,
в открытом море, преимущественно над
глубиной от 40 до 200 м, при температуре
воды от 13 до 24°С и солености от 8 до 12‰.
Совершает суточные вертикальные
миграции, поднимаясь ночью к поверхности
и опускаясь вглубь днем. Основной объект
питания анчоусовидной кильки —
веслоногий рачок эвритемора. Анчоусовидная
килька далеко не так жирна, как
обыкновенная: содержание жира в ее теле не
превышает 6,4%.
Фауна СССР. Рыбы. Том II, вып.1. Сельдевые (Clupeidae). А.Н.Световидов 1952 г.
*2. Clupeonella engrauliformis (Borodin) — Анчоусовидная тюлька, или сарделька, анчоусовидная килька (неправ.), у берегов Туркмении сардинка (неправ.) (табл. VII).
— Clupea engrauliformis Бородин, Вестн. рыбопр., XIX, 6, 1904 : 335 (Буйнак (Тип не указан, но экземпляры, по которым дано первоописание, сохранились. За тип следует считать один из этих экземпляров длиной 124 мм, Каспийское м. против Буйнак, 11 IV 1904, № 13860 коллекции Зоологического института Академии Наук СССР.), средняя часть Каспийского м.); там же, XXI, 4, 1906 : 197—203, фиг. 1 на стр. 199 (против Буйнак, а и f табл. измер., против Апшеронского п-ова, b—е табл. измер.); Суворов, Тр. Касп. эксп. 1904 г., II, 1908 : 50, 60, фиг. на стр. 51 (средняя и южная части Каспийского м., по Бородину).
— Harengula engrauliformis Берг, Мат. позн. русск. рыбол., II, 3, 1913 : 46, табл. I, фиг. 3 (средняя часть Каспийского м. от о. Тюленьего до Апшеронского п-ова); Суворов, там же, III, 3, 1914 : 1 (количества позвонков, распространение, биология).
— Harengula grimmi (non Kessler) Берг, Рыбы пресных вод, 1923 : 29; Малятский, Тр. Азовско-черном. научн. рыбохоз. ст., 6, 1930 : 67 (измерения).
— Harengula engrauliformis Сергеева и Ганешина, Бюлл. Всекасп. научн. рыбохоз. эксп., 5—6, 1932 : 43 (количества лучей в плавниках, жаберных тычинок, позвонков).
— Clupeonella engrauliformis Берг, Рыбы пресных вод, I, 1932 : 107; Световидов, Докл. АН СССР, XXXI, 8, 1941 : 806; Зоолог. журн., XXII, 4, 1943 : 231; Докл. АН СССР, XLVI, 5, 1945 : 226; Чугупова, в: Промысловые рыбы СССР, 1949 : 89.
Распространение, биология. Недошивин, Сборн. в честь Книповича, 1927 : 296 (искусственное оплодотворение, развитие); Расс и Халдинова, Сборн., посвящ. деят. Книповича, 1939 : 302 (развитие и распространение икринок и личинок); С. Пробатов, Изв. Азербайджанск. научно-исслед. рыбохоз. ст., 4, 1939 : 3, 32 (размеры, вес, миграция, биология); Пожалуева, Рыбн. хоз., 1940, 1 : 26 (распространение, биология); Ловецкая, там же : 27 (распространение, биология); там же, 1941, 3 : 20 (зрелость, нерест); Каспийские кильки, изд. Азербайджанск. научно-исслед. рыбохоз. ст., 1941 : 6, 13, рис. 2, карта 1—4 (биология, распределение, промысел); Приходько, Тр. Волго-каспийск. научн. рыбохоз. ст., IX, 1947 : 65 (восточные берега средней части моря, летом); Радаков и Токарев, Рыбн. хоз., 1948, 5 : 13 (лов на свет); Токарев, там же, 1949, 6 : 33 (восточное и западное побережья моря, лов на свет); Ловецкая, там же, 1951, 1 : 36 (западное побережье южной части моря; с 4 по 18 июля и с 17 по 22 сентября 1950 г.; лов на свет); Приходько, там же, 1951, 8 : 43 (по обе стороны южной части моря, с 25 февраля по 19 марта 1951; лов на свет); Борисов, там же, 1951, 11 : 9 (северо-восточное побережье средней части моря, с 15 июня по 25 сентября 1951 г.; лов на свет); Ловецкая, Каспийские кильки и их промысел, Пищепромиздат, 1951 : 5, 6, 7, 8 и сл., рис. 2, 4—10 (распределение, миграции, размножение, размеры, возраст, рост, промысел); Н. Танасийчук, Промысл. рыбы Волго-Каспия, Пищепромиздат, 1951 : 50 (размеры, вес).

D III 12—14 (12.8), общим числом 15—17 (15.7), А III 15—19 (16.8), общим числом 18—22 (19.8), sp. br. 56—67 (61.3), килевых чешуй 23—31 (27.6), vert. S. 44—47 (46.07), caud. vert. 25—28 (26.52). Тело вальковатое, брюшко снизу закругленное, килевые чешуи слабые. Голова короткая и широкая, длина ее 20.4—23.1 (21.4)% длины тела, межглазничный промежуток 16.2—18.7 (17.5)% длины головы. В процентах длины тела до конца хвостового плавника: длина головы 18.5— 21.0 (19.5), длина грудного плавника 12.0—14.7 (13.5), длина брюшного плавника 7.4—9.4 (8.4), наибольшая высота тела 14.6—17.5 (16.2), наименьшая высота тела 5.6—6.9 (6.1), высота головы у затылка 10.9—13.1 (12.2). Глаза небольшие, 21.6—27.3 (24.9)% длины головы, заглазничный отдел удлиненный, 39.2—44.8 (42.0)%, рыло довольно укороченное, 30.4—34.8 (32.7)% той же длины. Челюсти короткие, верхняя челюсть 35.8—41.5 (39.0)% длины головы, нижняя челюсть 47.5—53.0 (49.5)% той же длины. Парные плавники короткие, грудные плавники на конце заостренные, составляют 13.2—16.2 (14.9)% длины тела или 40.4—54.1 (48.8)% расстояния между их основанием и основанием брюшных плавников, брюшные плавники 8.1—10.3 (9.2)% длины тела. Тело низкое, наибольшая высота 16.0—19.2 (17.8)% длины его, наименьшая высота 6.2—7.6 (6.7)%, высота головы 12.0—14.4 (13.4)% той же длины. Спина и верхняя часть головы у живых темносиние, с зеленым или оливковым оттенком, у мертвых светлеют или иногда темнеют, становясь почти черными, в особенности в местах соприкосновения с нитями сетей. Все плавники бесцветные, за исключением хвостового, почти черного у основания, и спинного с неясной темной поперечной полосой посредине. Длина до 155, наичаще 115—125 мм, вес до 26.4 г, наичаще 10.5—16.5 г. Описание дано по 64 экз. длиной от 113 до 150, в среднем 122.3 мм, собранным 8—9 XI 1938 в южной части Каспийского моря против Апшеронского п-ова. По данным Сергеевой и Ганешиной (1932), А 15—16 (15.4), жаберных тычинок 57—63 (60), позвонков 44—47 (46). По сообщению Е. В. Пожалуевой, у исследованных ею рыб от берегов Туркмении позвонков 44—48 (46.0).
Солоноватоводная рыба, в пресные воды и в сильно опресненные части моря не заходит.
Распространение. Средняя и южная части Каспийского моря, в северной части встречается не далее границы опреснения. Нерест главным образом в средней и южной частях моря вдоль западных и восточных берегов как в открытом море, так и в прибрежной области, а также и на мелководье в северных районах средней и южных районах северной частей.
Биология. Встречается главным образом в открытом море, но иногда подходит и к берегам. В зимнее время держится в южной части моря, преимущественно над глубинами от 40 до 750 м при температуре воды 8.5—9° и выше, до 13.5°. В средней части моря встречается в незначительных количествах, преимущественно в южной половине. В теплые зимы держится как в западной, так и в восточной половине южной части моря, в холодные отходит в его южные, юго-восточные и восточные районы. Весной, с апреля или мая одни стада начинают мигрировать в среднюю и частично в северную часть моря, подходя к берегам лишь в очень редких случаях, изредка довольно в большом количестве, и придерживается обычно свала и струи постоянного течения вдоль западного и в меньшей степени восточного побережья. В 1946 и 1947 гг. в большом количестве обнаружена летом у восточных берегов от п-ова Мангышлака до зал. Кендырли, где в 1946 г. составляла 32% (8.5 тыс. ц) в уловах обыкновенной тюльки. Держится и здесь на некотором отдалении (1—10 миль) от берегов, в более открытых частях моря (в 10—20 милях от берега) встречались косяки, состоявшие исключительно из этого вида (Приходько, 1947). В северную часть моря дальше границы опреснения не идет. Другие стада остаются в южной части моря, концентрируясь у западного и восточного побережий, точно также не подходя к берегам, за исключением немногих более или менее приглублых районов (над глубинами более 25 м) у берегов северной части Туркмении, где подходит летом и осенью — в 1935 г. в мае—июне и в октябре; в последние годы подходы к берегам здесь незначительны. С конца сентября начинается отход из северной половины средней части моря, но в ее южной половине, преимущественно у западных берегов, держится в массе в течение всей осени и ловится в наибольших количествах над небольшими глубинами (25—100 м), богатыми в это время планктоном. Миграции, как и весной, происходят в области гидрологического течения вдоль западного и отчасти восточного побережий. С охлаждением поверхностных слоев воды до 5—6° опускается в нижние горизонты и держится в более глубоких слоях воды, наичаще от 8 до 32 м; в самых поверхностных слоях воды скопления наблюдаются редко, ниже 32 м, хотя и встречается, но в незначительном количестве. Весной, с началом прогревания воды до 11—15° и выше подымается к поверхности, где преимущественно и держится летом при температуре воды около 16—17°. В июле—сентябре при прогревании воды до 26—28° и выше отходит дальше от берегов и вновь опускается в нижние слои, в октябре—ноябре подымается опять к поверхности (Ловецкая, 1951). Однако ловом на электросвет установлено, что по обе стороны южной части моря с конца февраля по середину марта 1951 г. держалась главным образом на глубине от 30 до 60 м, встречалась также до глубины 78 м (Приходько, 1951). В первой половине июля и во второй половине сентября 1950 г. у западных берегов южной части моря тем же способом лова обнаружено, что в наибольшем количестве держалась в зоне температурного скачка, на глубине 35—42 м. В зависимости от глубины расположения слоя температурного скачка летом и осенью в наибольшем количестве может держаться даже на глубине и более 40 м (Ловецкая, 1951). В период икрометания в большинстве случаев преобладают самцы (73.5%), которые, повидимому, держатся на нерестилищах более долгое время, в то время как самки уходят сразу же после выметывания икры. По окончании нереста при переходе к интенсивному питанию в уловах преобладают самки. Самцы во время нереста, как правило, держатся преимущественно на глубине 10—20 м, самки — на глубине 20—25 м. Самки немного крупнее самцов: средние размеры самок 116 мм, самцов — около 112 мм, средний вес самок 11 г, средний вес самцов — около 10 г. Нерест происходит с конца апреля — начала мая по ноябрь, наиболее интенсивно в июле, при температуре воды 13—24° и солености 8—13‰, в открытом море с глубинами от 10—20 м и выше, в массе над глубинами от 50 до 200 м. Икрометание порционное. Развитие икринок при температуре 17.9° происходит в 30—34 часа. В открытом море, как и Clupeonella delicatula caspia, планктоноядна (Copepoda, пелагические Mysidae и Gammaridae), причем нередко поедает мальков Clupeonella grimmi (?) и даже личинок сельдей. У берегов питается тем же, чем и Clupeonella delicatula caspia (Copepoda, Cumacea, Schizopoda и Amphipoda). Половой зрелости достигает в трехгодовалом возрасте, частично и в двухгодовалом. У берегов Туркмении зимой и весной в уловах дрифтерных сетей преобладают четырех- и трехгодовалые, наибольший возраст — семигодовалый (Ловецкая, 1941, 1951). Обладает наилучшим ростом по сравнению с другими видами рода, (по данным Ловецкой, 1951):
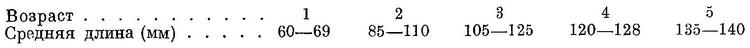
Хозяйственное значение несмотря на большие запасы до последнего времени было ничтожно. Уловы могут быть значительно увеличены путем организации промысла в открытом море, в частности весьма перспективен лов на свет. Ловится прибрежным промыслом и учитывается вместе с Clupeonella delicatula caspia. У восточных берегов средней части моря уловы ее выше последней. Благодаря применению лова на электросвет, уловы в 1951 г. превысили уловы обыкновенной тюльки. Повидимому, большую роль играет в питании хищных сельдей и некоторых других рыб.
Рыбы пресных вод СССР и сопредельных стран. Л.С.Берг 1948 г.
Clupeonella engrauliformis (Borodin). — Анчоусовидная тюлька
Clupea engrauliformis Бородин, Вестн. рыбопр., 1904, стр. 335 (Каспийское море: Буйнак); 1906, стр. 197—203, рис. (против Апшеронского полуострова, Буйнак); Тр. Касп. эксп. 1904 г., II, 1908, стр. 50.
Harengula engrauliformis Берг, Мат. к позн. русск. рыбол., II, вып. 3, 1913, стр. 46, табл. I, фиг. 3 (от о. Тюленьего до Апшеронского полуострова, открытое море в южной части).
Harengula grimmi (non Kessler) Берг, Рыбы пресн. вод России, 1923, стр. 29.
Clupeonella engrauliformis Берг, Рыбы пресных вод СССР, I, 1932, стр. 107. — Расс и Халдинова, Сборник, посвящ. Книповичу, 1939, стр. 302, рис. 2—4, 7—8 (икра, личинки, мальки). — Ловецкая, Касп. кильки, 1941, стр. 6, 13, рис. 2, карты 1—4.
D III 12, А III 15—16, поперечных рядов чешуй 47, брюшных шипов 28. Жаберных тычинок 57—63. Позвонков 44—47. Тело низкое, вальковатое, высота его не более 19% его длины (до конца средних лучей С). Брюшные шипы слабо развиты. Длина до 155—165 мм (абс.). Средний вес 10—18 г, наибольший 26 г. Спина темная.
Каспийское море в средней и южной частях; в пресную воду не входит. Нерест с конца мая по ноябрь. Согласно мнению Т. С. Расса и Н. А. Халдиновой (1939, стр. 309), осенние нерестующие особи либо образуют особую форму, либо эта тюлька нерестится два раза в год: соответственно двукратному наступлению благоприятной для нереста температуры.
Clupeonella engrauliformis. Встречается единично в северном Каспии. При лове на свет летом 1948 г. добывались кильки размерами от 7 до 14 см и весом от 3 до 16 г. Наибольшие уловы были на глубинах от 10 до 20 м при температуре воды 18—20°. В июне в большом количестве приближается к восточным и западным берегам среднего Каспия; с сентября по декабрь отходит в южный Каспий. Нерест продолжается с мая по октябрь. Плодовитость в среднем около 40 тыс. икринок (А. К. Токарев, Рыбн. хоз., 1949, №6).