Clupeonella caspia Svetovidov, 1941
Каспийская обыкновенная килька, каспийская килька, каспийская тюлька, южнокаспийская тюлька, обыкновенная каспийская тюлька, обыкновенная килька (Rus),
Caspian tyulka, Common Caspian kilka
(Eng)
Синонимы:
Clupeonella cultiventris caspia (Svetovidov, 1941) Каспийская килька (Rus),
Clupeonella delicatula caspia Svetovidov, 1941 Каспийская тюлька (Rus),
Атлас-определитель рыб: Книга для учащихся. Н.А.Мягков 1994 г.
Килька каспийская — Clupeonella cultiventris caspia (Svetovidov, 1941)
Внешне очень схожа с подвидом Clupeonella cultiventris cultiventris. Достигает длины 15 см. Солоноватоводный и реже морской подвид. В последние годы в массе проник и размножился в волжских водохранилищах, где образовал пресноводную форму. Подвид распространен в Каспийском бассейне. Объект промысла.
Словарь названий пресноводных рыб СССР. Г.У.Линдберг и А.С.Герд 1972 г.
3.3.2 (2). Clupeonella delicatula caspia Svetovidov, 1941 — Каспийская тюлька (Св., 1952 : 199).
[1] русск. Каспийская килька — Суворов, 1904 : 48. Каспийская тюлька — Св., 1952 : 199. Обыкновенная каспийская тюлька, Обыкновенная килька — Пром. р. : 85. Сарделька Касп. м., Сардинка Касп. м. — Зол. [18] англ. Common Caspian kilka Канада — Ricker : 48.
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
Каспийская обыкновенная килька
(Glupeonella delicatula caspia) представляет
собой подвид черноморско-азовской
тюльки, отличающийся большей величиной, до
14—15 см у длительностью жизни до 6 лет
и несколько меньшей жирностью, до 12%
содержания жира в теле. Позвонков у нее
41—45. Обыкновенная каспийская килька
зимует обычно в Среднем и Южном
Каспии, а в марте идет на север, в Северный
Каспий, подходя к берегам при
температуре воды от 6 до 14°С и частично входя
в дельты Волги и Урала. Разгар нереста
кильки в Северном Каспии в апреле —
мае, при температуре 12—21°С.
Подходящая к берегам килька образует огромные
косяки, подчас заполняющие сплошной
полосой рыбы всю прибрежную отмель.
Появляясь внезапно у берегов, килька
также быстро уходит в открытое море,
где держится преимущественно в слое от
6 до 30 м, опускаясь иногда и до 100 м.
Питается преимущественно веслоногими
рачками каланипедой и гетерокопе.
В затонах и ильменях Волги и в озере
Чархал в бассейне Урала она образует
мелкую пресноводную форму — до 11 см
длины.
Фауна СССР. Рыбы. Том II, вып.1. Сельдевые (Clupeidae). А.Н.Световидов 1952 г.
*1b. Clupeonella delicatula caspia Svetovidov — Каспийская тюлька, или сарделька, обыкновенная килька (неправ.) (табл. VIII).
— Clupea delicatula Кесслер, Рыбы Арало-касп.-понт. обл., 1877 : 276 (частью: Каспийское м. у берегов Мангышлака); Суворов, Тр. Касп. эксп. 1904 г., II, 1908 : 48, 60 (по всему Каспийскому м.; устья рек, описание, биология).
— Harengula delicatula Берг, Мат. позн. русск. рыбол., II, 3, 1913 : 45, табл. I, фиг. 2 (частью: Каспийское м., дельта Волги и Урала); Рыбы пресных вод, 1916 : 31 (частью); Сергеева и Ганешина, Бюлл. Всекасп. научн. рыбохоз. эксп., 5—6, 1932 : 43 (количества лучей в плавниках, жаберных тычинок, позвонков).
— Clupeonella delicatula Берг, Рыбы пресных вод, I, 1932 : 102 (частью).
— Clupeonella delicatula caspia Световидов, Докл. АН СССР, XXXI; 8, 1941 : 806 (отличие от типичной формы вида. Тип — экземпляр длиной 152 мм, дельта Волги, Оранжерейный промысел, 1885 г., Гримм, № 15883 коллекций Зоологического института Академии Наук СССР.); Зоолог. журн., XXII, 4, 1943 : 231; Докл. АН СССР, XLVI, 5, 1945 : 227 (описание); Чугунова, в: Промысловые рыбы СССР, 1949 : 85, табл. 21.
Распространение, биология. Бородин, Ежег. Зоолог. муз. Акад. Наук, I, 1—2, 1896 : 89, фиг. 1 (устье Урала, дельта Волги); Lönnbеrg, Bihag. К. Svensk. Vet.-Akad. Handl., XXVI, afd. IV, 8, 1900 : 28 (Петровск, Мангышлак); Суворов, Мат. позн. русск. рыбол., III, 3, 1914 : 3 (распространение, размеры, нерест, промысловое значение); Киселевич, Промысловые рыбы Волго-каспийского района Астрах., 1926 : 26 (биология); Белоусов, Бюлл. рыбн. хоз., 1926, 6 : 12 (дельта Волги, ход, промысел); Киселевич, «Наш край», 1927, 9—10 : 11 (дельта Волги; биология, промысел); Недошивин, Сборн. в честь Книповича, 1927 : 290 (искусственное оплодотворение, развитие); Тр. Астраханск. научн. рыбохоз. ст., VI, 4, 1928 : 92 (дельта Волги; рост молоди); Дмитриев, Бюлл. рыбн. хоз., 1928, 7 : 17 (у берегов Дагестана; промысел); Изв. Дагестанск. ихт. лаб., I, 1929 : 21 (у берегов Дагестана; биология, промысел); Бюлл. рыбн. хоз., 1930, 7—8 : 13 (у берегов Дагестана; промысел); Беляев, Бюлл. Всекасп. научн. рыбохоз. эксп., 1—2, 1932 : 62 (у берегов Азербайджана; промысел, нерест в зимнее время); Леванидов, там же, 5—6, 1932 : 56 (жирность); Подсевалов, Изв. Астраханск. отд. Научно-исслед. инст. рыбн. промышл., II, 1935 : 256 (иранское побережье; жирность, уловы); Дмитриев, Рыбн. хоз. СССР, 1935, 10 : 16 (у берегов Дагестана; прилов молоди других рыб); Шульман, Рыбн. хоз. СССР, 1936, 3 : 42 (жирность, химический состав жира); Чаянова, Тр. I Всекасп. научн. рыбохоз. конфер., I, 1936 : 51 (питание); Киселевич, Сельди северного Каспия, Сталингр., 1937 : 37 (биология); Клыков, Тр. Касп. ком. АН СССР, II, 1938 : 80 (в громадном количестве заходит в западную часть зал. Мертвый Култук одновременно с долгинкой и немного ранее); Перцева, Рыбн. хоз., 1938, часть Каспийского м.; места и время нереста); Расс и Халдинова, Сборн., посвящ. деят. Книповича, 1939 : 307 (развитие и распространение икринок и личинок); Левоник, Рыбн. хоз., 1939, 5 : 9 (биология, промысел); С. Пробатов, Изв. Азербайджанск. научно-исслед. рыбохоз. ст., 4, 1939 : 3 (биология, промысел); Пожалуева, Рыбн. хоз., 1940, 1 : 26 (у берегов Туркмении, распределение, промысел); Ловецкая, там же : 27 (средняя и южная часть Каспийского м.; распределение, промысел); там же, 1941, 3 : 20 (зрелость, нерест); Каспийские кильки, изд. Азербайджанск. научно-исслед. рыбохоз. ст., 1941 : 4, 15, рис. 1, карты 1—4 (биология, распределение, промысел); Казанчеев, Рыбн. хоз., 1941, 4 : 20 (восточное побережье средней части моря; лов летом и осенью); Шорыгин, Рыбн. промышл. СССР, 1—2, 1944 : 3 (промысел); Борисов, там же, 3, 1945 : 55; Рыбн. хоз., 1946, 2—3 : 32; 10—11 : 38; 1947, 10 : 23 (лов на свет, размеры); Краснова, Докл. Всесоюзн. Научно-исслед. инст. морск. рыбн. хоз. и океаногр., 8, 1947 : 15 (северная часть моря; условия нереста); Казанчеев, Тр. Волго-каспийск. научн. рыбохоз. ст., IX, 1, 1947 : 38 (северная часть моря; биология); Приходько, там же : 63 (восточное побережье средней части моря; летние подходы); Ловецкая, Докл. Всесоюзн. Научно-исслед. инст. морск. рыбн. хоз. и океаногр., 9, 1947 : 9 (прилов молоди рыб в ставных килечных неводах); Радаков и Токарев, Рыбн. хоз., 1948, 5 : 13 (лов на свет); Демин, Тр. Дагестанск. сельскохоз. инст., IV, 1, 1949 : 159 (берега Дагестана; миграции); Приходько, Рыбн. хоз., 1951, 8 : 43 (по обе стороны южной части моря, с 25 февраля по 19 марта; лов на свет); Ловецкая, Каспийские кильки и их промысел, Пищепромиздат, 1951 : 5, 6, 15 и сл., рис. 1, 4—10 (распределение, миграции, размножение, размеры, возраст, рост, промысел); Н. Танасийчук, промысл. рыбы Волго-Каспия, Пищепромиздат, 1951 : 47 (размеры, вес); В. Танасийчук, Тр. Всесоюзн. Научно-исслед. инст. морск. рыбн. хоз. и океаногр., XVIII, 1951 : 21 (северная часть моря; учет молоди, размеры мальков); Перцева-Остроумова, там же: 106, рис. 2 (описание икринок и личинок); Халдинова, там же : 106 (зал. Мертвый Култук и Кайдак и прилегающие районы; нерест, распределение икринок и личинок); Чаянова, там же : 245, рис. 1—3 (северная часть моря; питание).

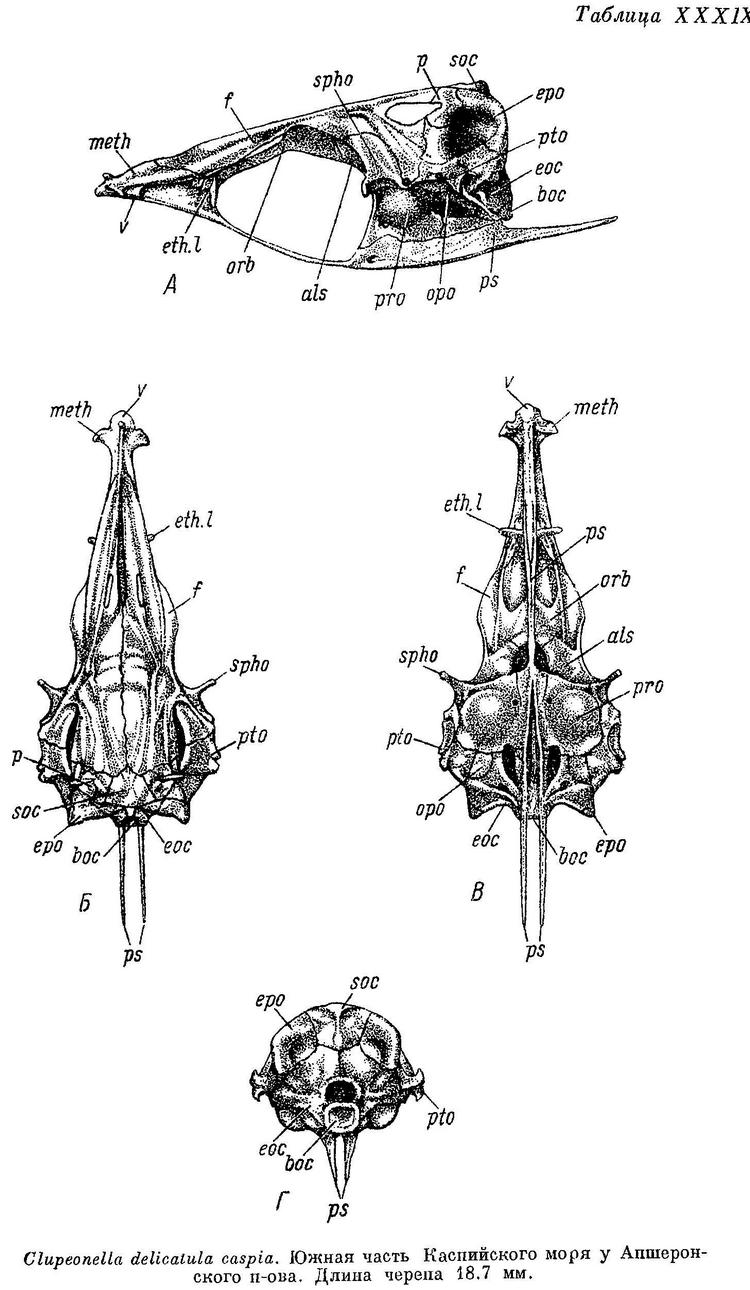
D III—IV 11—13 (11.9), общим числом 14—16 (15.4), А III 14—18 (16.2), общим числом 17—21 (19.1), sp. br. 51—62 (56.6), килевых чешуй 26—30 (27.9), vert. S. 41—44 (42.65), caud. vert. 24—27 (25.70). Тело и в особенности брюшко сжатые с боков, брюшко снизу заостренное, брюшные шипы хорошо развитые. Голова укороченная и широкая, длина ее 20.9—24.2 (22.5)% длины тела, межглазничный промежуток 17.7—21.5 (18.8)% длины головы. В процентах длины тела до конца хвостового плавника: длина головы 18.4—21.3 (19.8), длина грудного плавника 13.7—16.5 (14.9), длина брюшного плавника 7.6—9.2 (8.4), наибольшая высота тела 15.8—21.3 (18.1), наименьшая высота тела 4.9—6.9 (6.2), высота головы у затылка 13.0—15.3 (14.1). Глаза не особенно большие, 22.8—29.1 (25.9)% длины головы, заглазничный отдел удлиненный, 38.2—44.6 (41.8)%, рыло укороченное, 29.2—35.3 (31.9)% той же длины. Грудные и брюшные плавники укороченные, грудные плавники 15.6—18.7 (16.9)% длины тела или 52.3—65.2 (57.8)% расстояния между их основанием и основанием брюшных плавников, брюшные плавники — 8.6—10.5 (9.5)% длины тела. Тело довольно высокое, наибольшая высота 18.0—24.2 (20.6)% длины его, наименьшая 5.6—7.8 (7.0)%, высота головы 14.8—17.4 (16.0)% той же длины. Спина и верхняя часть головы у экземпляров, пойманных в море, светлозеленые. Все плавники бесцветные, за исключением хвостового, обычно темноватого при основании, и спинного с неясной, слабой темной поперечной полосой посредине. Длина до 145 мм, наичаще 80—100 мм, вес до 19 г, наичаще 10—13 г. Описание дано по 50 экз. длиной от 76.0 до 119.5, в среднем 93.4 мм, пойманным 26 II—30 IV 1911 (30 экз.) и в мае 1885 г. (20 экз.) в дельте Волги (Оранжерейный промысел). У 39 экз. длиной от 80.7 до 134.5, в среднем 114.6 мм, собранных 8—9 XI 1939 в южной части Каспийского моря против Апшеронского п-ова, D III 11—14 (12.1), А III 15—19 (16.5), sp. br. 50—63 (57.8), позвонков 42—44 (43.1). По данным Сергеевой и Ганешиной (1932), у экземпляров из южной части моря А 14—17 (15) (Количество лучей в анальном плавнике здесь преуменьшено, повидимому вследствие ошибки в разграничении разветвленных лучей от неразветвленных.), жаберных тычинок 52—64 (58). По сообщению E. В. Пожалуевой, у исследованных ею экземпляров от берегов Туркмении позвонков 41—45 (43.1).
В высокой степени евригалинная форма, встречающаяся, как в воде с соленостью до 15‰ Сl, так и в совершенно пресной.
Биологически крайне изменчивая форма. Часть каспийской тюльки проводит всю жизнь в море (например в южной части Каспийского моря), здесь же и размножается, другая часть является в сущности проходной рыбой, идущей на нерест в Волгу. Возможно, что обе эти группы, тюльки представляют самостоятельные формы.
Распространение. Каспийское море, где встречается почти повсеместно, от сильно осолоненных частей (до 34.15‰ в зал. Кайдак); (Халдинова, 1951), до совершенно опресненных, заходит в низовья рек Волги и Урала, иногда подымаясь высоко по течению: по Волге до Вольска и выше, в притоки среднего течения ее (Еруслан, Б. Иргиз). Икрометание почти повсеместно, как в море, так и в пресной воде предустьевых пространств и низовий рек (Волга, Урал, Терек). В сильно осолоненных частях (например зал. Кайдак) нерест не происходит. В северной части моря нерестует главным образом в юго-западном и западном районах моря (от о. Чечень до Лагани), в предустьевом районе Волги (вдоль свала до Белинской банки), у Забурунья, перед устьем Урала, у берегов пол. Бузачи—Мангышлак. В средней и южной частях моря икрометание происходит вдоль западного и восточного побережий, в заливах (Туркменском, Кирова).
Биология. Держится как в открытом море, так и у берегов, куда в массе подваливает весной. В зимнее время держится вдоль западных берегов в южной части моря в районах с глубинами 35—100 м (от банки Макарова до Куринской) и несколько южнее (Норд-остов Култук—Астара), где встречается в промысловом количестве даже в местах с глубинами 10—20 м; часть зимует на юге южной части моря. В центральной части моря и над глубинами свыше 100—200 м встречается редко. В средней части моря зимой обнаружена в районе Дивичи, есть, повидимому, и в других местах, возможно, и в северной части подо льдом (Ловецкая, 1941, 1951). Зимой в южной части моря держится главным образом при температуре 7—10°, но иногда встречается при 3—4° и даже 2.6°. Весной одни косяки подходят к западным, другие к восточным берегам южной части, третьи вдоль западных берегов направляются в среднюю и далее в северную часть моря, где также подходят к берегам, и в Волгу. Массовые миграции происходят с середины — конца марта, вдоль западного побережья средней части моря в зоне свала над глубинами 35—100 м. Первые подходы к берегам происходят при температуре около 4.5—5.5°, массовые подходы при температуре около 10°, при 13° и выше отходит, предельная температура 23—25° (Казанчеев, 1947). При летних подходах у восточных берегов средней части моря наибольшие скопления образуются при 10—17° (Приходько, 1947). Подходы у берегов Азербайджана начинаются с февраля, массовые подходы в южной н средней частях моря — в марте—апреле. Подходы у берегов Дагестана усиливаются при ветрах восточной четверти, дующих со стороны моря, наиболее благоприятен восточный ветер (Демин, 1949). В северной части моря появляется в начале апреля вместе с долгинской сельдью, главная масса появляется при температуре 10—12°. В Волгу ход начинается в половине апреля, во второй половине апреля при температуре около 10° массовый ход, разгар хода в начале мая; ход очень дружный, продолжается дней 7—10, за 2—3 дня до хода сельди или одновременно с ним. В отличие от сельди идет не серединой протока с наибольшей глубиной, а ближе к берегу, по мелякам, преимущественно утром и в первой половине дня (с 7—8 до 15 час.), затем ход ослабевает, а ночью прекращается совсем. Основная масса заходит в самую нижнюю часть дельты и не подымается вверх. В 1925—1927 гг. ловилась на тоне Первомайской, находившейся в то время в нижней части дельты; в 1942 г. лов на этой тоне был безуспешен ввиду того, что тоня вследствие понижения уровня моря отстояла от выхода реки в море уже на 20—25 км (Казанчеев, 1947). Летом держится у западных и восточных берегов средней и южной частей моря (Махачкала—Каякент, Худат—Дивичи, Апшерон—Астара, зал. Александер-бай, Казахский и др.), в отдалении от берегов, на глубинах от 20—40 до 100 м; у восточных берегов средней части моря временами в зависимости от сгонных ветров и распределения планктона подходит в прибрежную зону. Миграции на зиму в южную часть моря происходят в ноябре. Из северной части моря взрослые уходят сразу после окончания нереста, из Волги и ее дельты — вскоре после нереста или остаются до осени, иногда, будучи запертыми в ильменях и затонах, остаются на зиму, причем молодь превращается в морфу tscharchalensis. После отхода из северной части моря летом и отчасти осенью в массе подходит к восточным берегам от п-ова Мангышлак до зал. Кендырли, где усиленно откармливается (Приходько, 1947). Зимой и в начале весны в большинстве случаев в массе держится на глубине ниже 8 м, в апреле и мае в связи с прогреванием воды в прибрежной области подходит на мелководья до глубин 1—2 м. Летом в наиболее жаркое время (июнь—август) при прогревании поверхностных слоев воды до 25—28° в массе держится ниже этих слоев. В начале осени (в сентябре) с охлаждением поверхностных слоев воды поднимается в верхние горизонты и по мере дальнейшего охлаждения держится на глубине ниже 8 м от поверхности (Ловецкая, 1951). В 1951 г. с конца февраля по середину марта по обе стороны южной части моря в наибольшем количестве при лове на свет обнаружена на глубине от 20 до 40 м, в двух случаях наибольшее количество было на глубине 60 м (Приходько, 1951). Время нереста зависит от температуры воды. В северной части моря перед дельтой Волги в 1940—1941 гг. икрометание происходило со 2—3-й декады апреля, разгар в мае, продолжаясь единично в июле, при температуре воды от 11 до 20°, солености от 0 до 15‰, на глубине от 1 до 9 м. При повышении температуры выше 20° нерест незначителен, при 25° и выше прекращается (Краснова, 1947). На мелководье перед входом в Мертвый Култук и в прилегающих ко входу частях залива икринки встречались при более высокой солености, при 20‰ и выше икринки отмирают (Халдинова, 1951). В южной части моря нерест сильно растянут, происходит с января—февраля; наиболее развитые половые продукты бывают здесь в феврале—марте и вплоть до лета. В промысловых уловах круглый год обычно преобладают более крупные по размерам самки, среди мелких — самцы, которые созревают раньше (Казанчеев, 1947). Плодовитость тюльки от берегов Дагестана исчисляется от 9.5 до 60 тыс., в среднем 31.2 тыс. икринок, в дельте Волги, где тюлька мельче, несколько меньше, от 9.2 до 17.4 тыс., в среднем 14 тыс. икринок. Относительная плодовитость (количество икринок на 1 г веса тела) значительно выше, от 4 до 13 раз, чем у сельдей из рода Alosa (Дмитриев, 1929). Благодаря быстрому созреванию и высокой относительной плодовитости обладает большой способностью к восстановлению своих запасов. Икра выметывается не вся сразу, а несколькими порциями. Развитие икры при 8—9° длится 92 часа, при средней температуре 17.6° — 27—30 часов (Ловецкая, 1951). Состав пищи в основном зависит от состава зоопланктона. В северной части моря пища состоит главным образом из Copepoda (56%) — Calanipeda aquae-dulcis и Heterocope caspia, Cladocera (20%), Evadne trigone, Evadne camptonyx, Evadne hircus, Cercopages gracillina, временами планктоно-бентические ракообразные — Mysidae, Amphipoda, Cumacea, на местах икрометания икра этого же вида. Питается в течение всего светлого времени суток, ночью не питается или питается в незначительной степени. Питается и в период икрометания, но в незначительной степени (Чаянова, 1951). Половой зрелости особи, нерестящиеся в Волге и в северной и средней частях моря, достигают на втором году жизни, в южной части частично в возрасте даже менее одного года. Это, повидимому, будут рыбы, выведшиеся из икры поздней осенью и зимой и поэтому не имеющие на чешуе первого зимнего кольца. Тюльки из северной и средней частей моря (от берегов Дагестана) отличаются от тюлек из южной части, главным образом, меньшими приростами за первый год, достигая большего возраста. Кривая прироста, за первый год у килек из южной части моря двухвершинна, первый максимум (около 7 мм) соответствует приросту особей от весеннего нереста, второй (около 10 мм) — приросту особей от осеннего нереста. Нерестовые косяки в северной части моря в 1941 г. состояли из рыб от одно- до шестигодовалого возраста, преобладали одногодовалые и четырех- и трехгодовалые, что, повидимому, было обусловлено различной урожайностью отдельных поколений (Казанчеев, 1947). В средней части моря в уловах преобладают двух- и трехгодовалые особи, встречаются одно-, четырех- и пятигодовалые. В дельте Волги преобладают двух- и трехгодовалые, четырехгодовалых мало, пятигодовалых нет, есть годовики. В южной части моря в уловах преобладают двух- и трехгодовалые, шестигодовалые не встречаются (Ловецкая, 1951). Самки немного крупнее самцов, самки от берегов Азербайджана имеют в среднем длину 102— 107 мм, вес 8.5—10.0 г, самцы длину 93—101 мм, вес 6.5—8.5 г. В дельте Волги ловится несколько меньшего размера, что обусловлено, повидимому, отбирающим влиянием орудий лова. Растет значительно быстрее, чем черноморско-азовская форма (по данным обратного расчисления Киселевича, 1926 и Дмитриева, 1929):

Менее жирна, чем черноморско-азовская форма. В апреле у берегов Дагестана количество жира довольно значительно (10.84%), к началу мая в северной части моря значительно уменьшается (до 4.27—6.37%; Леванидов, 1932), у берегов Ирана (Ашур-аде) — 5.86% (Подсевалов, 1935).
Хозяйственное значение довольно существенное. Уловы за последние 15 лет возросли с 25 тыс. ц в 1929 г. до 169 тыс. ц. В Горганском районе в 1931—1932 г. поймано 240 ц, т. е: 0.2% всего улова (Подсевалов, 1935). Основная масса улова добывается у берегов Дагестана, Казахстана и в южном районе Азербайджана; в дельте Волги и у берегов Туркмении промысел развития не получил. Уловы могут быть значительно увеличены, так как запасы велики. Лов производится весной во время массовых подходов к берегам, главным образом в марте—апреле; в дельте Волги лов производился во время хода, в начале мая. К концу массовых подходов к берегам вместе с тюлькой начинает ловиться молодь сельдей, вследствие чего по существующим правилам лов должен прекращаться с 16 апреля и ранее, если в улове более двух дней подряд будет свыше 5% молоди других рыб. Главное орудие лова у берегов скипасти, в море также эломаны, в южной части моря ставные сети, в дельте Волги ловилась волокушами. Лов с помощью сильных источников света оказался также эффективным.
Одной из особенностей развития личинок и строения взрослых сельдевых следует считать различие в положении спинного плавника. Как известно, личинки сельдевых по сравнению со взрослыми, подобно личинкам и малькам многих костистых рыб, характеризуются сильно удлиненной передней частью тела, вследствие чего брюшные, анальный и спинной плавники у них отнесены далеко назад и, что самое существенное и свойственно только сельдевым и некоторым близким к ним семействам отр. Clupeiformes, спинной плавник у них расположен не над брюшными плавниками, как у взрослых, а значительно далее назади, между брюшными и анальным или почти над анальным (у Engraulidae). В процессе метаморфоза личинок передняя часть тела у них укорачивается вследствие сложного перемещения плавников. Спинной плавник при этом перемещается над миотомами вперед, а брюшные плавники назад, поэтому взаимное расположение обоих плавников становится таким, как у взрослых. Одновременно с этим анальный плавник перемещается вперед (Lebour, 1921; Ford, 1930). У взрослых сельдевых спинной плавник расположен над или несколько позади от брюшных в непосредственной к ним близости. При этом у разных видов семейства и видов одного и того же рода и даже форм вида положение спинного, брюшных и анального плавников неодинаково: у одних они расположены ближе к переднему концу тела, у других — дальше. Обусловлено это тем, что у разных родов и видов плавники закладываются в различном удалении от переднего конца тела, не над одними и теми же миотомами, а в дальнейшем степень перемещения плавников у разных родов и видов неодинакова (Lebour, 1921; Ford, 1930). Перемещение спинного плавника заканчивается к концу метаморфоза личинок и неоформившихся мальков, и у вполне сформировавшихся мальков и у молоди он расположен уже, как у взрослых. В дальнейшем, по мере роста молоди и взрослых рыб, как это видно на примере каспийской тюльки (Clupeonella delicatula caspia), положение анального и брюшного плавников несколько изменяется. (На каждый класс размеров приходится по 10 рыб. Как молодь, так и взрослые рыбы происходят из одного места (от западных берегов средней части Каспийского моря) и пойманы приблизительно в одно время. Такие же цифры рассматриваемых признаков получены автором и для тюльки из северной части моря перед дельтой Волги.)

Спинной плавник, как видно из приведенных примеров, занимает постоянное положение как у взрослых, половозрелых рыб, так и у рыб, еще не достигших половой зрелости. Что касается анального и брюшного плавников, то положение их на протяжении жизни рыбы претерпевает перемещение, судя по всему, в связи с половым созреванием вследствие растяжения брюшной стенки развивающимися половыми продуктами. Увеличение антеанального и антевентрального расстояний происходит по достижении рыбами размеров 70—80 мм, что приблизительно соответствует половому созреванию. В дальнейшем антеанальное расстояние остается примерно постоянным, увеличиваясь в незначительной степени, на доли процента. Антевентральное расстояние изменяется несколько в большей степени, повидимому, в связи с различным растяжением брюшной стенки вследствие разной степени наполнения полости тела половыми продуктами, у исследованных крупных рыб отчасти уже выметанными. К такому же выводу о постоянстве антедорсального расстояния, характеризующего положение спинного плавника у взрослых черноморско-азовских проходных сельдей (Alosa kessleri pontica), пришла Майорова (1939 : 7), а для черноморского анчоуса (Engraulis encrasicholus ponticus), относящегося к близкому сельдевым семейству того же подотряда, С. М. Малятский и отчасти Александров (1927 : 45, 46). Точно так же и относительно атлантической сельди (Clupea harengus harengus) установлено, что у взрослых рыб длиной свыше 10 см положение спинного, брюшных и анального плавников остается с возрастом неизменным (Heincke, 1898 : 113). Таким образом, у взрослых сельдевых положение спинного, а в известной мере также анального и брюшного плавников следует считать практически неизменным.
Рыбы пресных вод СССР и сопредельных стран. Л.С.Берг 1948 г.
Clupeonella delicatula (Nordmann). — Тюлька, обыкновенная тюлька
(На Каспийском море неправильно — килька, на Азовском и в Очакове — тюлька, в Ростове — сарделька.)
Clupea delicatula Nordmann, Faune pontique, 1840, p. 524 (Одесский рынок). — Кeсслep, Тр. СПб. общ. ест., V, 1874, стр. 309 (Черное море). — Antipa, Denkschr. Akad. Wien, math.-naturw. Kl., LXXVIII, 1905, p. 32—35, табл. II, фиг. 11—14 (дельта Дуная до Браилова, икрометание март—апрель ст. ст.). — Браунер, Тр. Бессарабcк. общ. ест., II, вып. 2, 1912, стр. 12 (Севастополь, Днестровский лиман, устье Днестра; жаберных тычинок 47—56).
Harengula delicatula Берг, Мат. к позн. русск. рыбол., II, вып. 3, 1913, стр. 45, табл. I, фиг. 2 (Каспийское море). — Суворов, там же, III, вып. 3, 1914 (Каспийское море; образ жизни). — А. Никольский, Бюлл. Черном.-азовск. ст., № 6—7, 1923, стр. 4 (Очаков). — Белинг, Тр. Черном.-азовск. научно-пром. ст., I, 1925, стр. 57 (Днепр до Берислава и Никополя); Збiрн. праць Днiпр. ст., № 2, Киев, 1927, стр. 338 (Бугский лиман; весной доходит по Бугу до порогов). — Тихонов, Бюлл. Черном.-азовск. ст., № 19—20, 1927, стр. 59 (Азовское море, северный берег). — Киселевич, Промысл, рыбы Волго-касп. района, 1926, стр. 26; Наш Край, 1927, № 9—10, стр. 12—14 (дельта Волги). — Чугунов, Тр. Астраханск. рыбохоз. ст., VI, вып. 4, 1928, стр. 92 (рост молоди в дельте Волги). — Н. Дмитриев, Изв. Дагестанск. ихтиол, лабор., I, 1929, стр. 21 (Каспийское море у берегов Дагестана; биология). — Недошивин, Тр. Азов.- черном. эксп., IV, 1929, стр. 36 (заходит весной в низовья Дона на нерест: Мертвый Донец, ст. Елизаветовская). — Телегин, там же, стр. 211 (Таганрогский залив).— Сергеева и Ганешина, Бюлл. Всекасп. рыбохоз. эксп., № 5—6, 1932, стр. 43 (Каспийское море).
Clupeonella delicatula Берг, Рыбы пресн. вод СССР, I, 1932, стр. 102. — Киселевич, Сельди сев. Каспия, 1937, стр. 37. — С. Троицкий, Природа, 1939, № 10, стр. 71 (дельта Кубани). — С. Пробатов, Изв. Азербайджанск. рыбохоз. ст., вып. 4, 1939, стр. 3—44 (от Зюдостова култука до Астары). — Расс и Халдинова, Сборник, посвящ. Книповичу, 1939, стр. 307, рис. 5—7 (Касп. м.; мальки). — Пожалуева, Рыбн. хоз., 1941, № 1, стр. 26 (юго-вост. часть Каспия). — Ловецкая, Рыбн. хоз., 1941, № 1, стр. 37 (Каспий; миграции); Рыбн. хоз., 1941, № 3, стр. 20 (Каспий; нерест); Каспийские кильки, Баку, 1941, стр. 4, 15, карты 1—4.
D III—IV 11—13, А III 14—18, V I (6) 7, брюшных шипов 24—30, жаберных тычинок 52—64, в среднем 58, позвонков 42—43, высота тела 20.7—24.7% его длины (до конца средних лучей С), длина головы 21.5— 20.7%, диаметр глаза 5.6—6.0% (предыдущее описание по экземплярам из южн. Каспия, Куринская коса, абсолютная длина 90—125 мм). Спина светлозеленая. Описание 2 экз. из устья Волги (Забурунье, № 15884) длиной (абс.) 114—115 мм. D IV 11—12, A III 16—18, V I 7, жаберных тычинок 53, высота тела в процентах его длины (до конца средних лучей С) 22.1—23.8, длина головы 21.8—21.6, диаметр глаза 5.3—6.0; более высокий экземпляр — самка с незрелой икрой. Описание 3 экз. из Ростова-на-Дону (№ 16117, «сарделька») длиной (абс.) 77—78 мм. D III 12—13, А III 15—16, V I 6—7, брюшных шипов 27—29, высота тела в процентах его длины (до конца средних лучей С) 23.5—24.2, длина головы 23.5—24.2, диаметр глаза 6.3—7.0. Экземпляр из Станислава, низовья Днепра (№ 21060), высота тела 23.5%.

Черное и Каспийское море, входит в реки. Дельта Дуная (вверх до Браилова и выше, до оз. Калараш), устье Днестра (Маяки), Буг до порогов, Днепровский лиман, Днепр до Никополя, дельта Дона, дельта Кубани (С. К. Троицкий), оз. Палеостом. Все берега Каспийского моря (есть и в Мертвом култуке), дельты Волги, Урала и Терека; в средн. и южн. Каспии в устья рек не входит; по Волге встречается вверх до Вольска и выше. Есть и в притоках сред. Волги: Еруслан, Б. Иргиз (Мною просмотрены многочисленные экземпляры, собранные летом 1930 г. Средневолжской экспедицией Института рыбного хозяйства и добытые в самой Волге между Песковаткой и Балаковым в затонах и в притоках; длина (абс.) до 107 мм.).
В дельте Волги косячный ход обычно начинается во второй половине апреля. В 1926 г. у с. Оля первые экземпляры тюльки появились 12 апреля, косячный ход начался 24 апреля, разгар хода был с 3 по 8 мая. Тюлька мечет икру частью в море перед дельтой Волги (См. Т. Перцева, Рыбн. хоз., 1938, № 7, стр. 31, 32.), частью в нижней и в средней части дельты. В 1927 г. в середине мая попадались тюльки с выметанной икрой. Средние размеры (до конца средних лучей С) в дельте Волги в миллиметрах: 1926 г. Самцы — 76, Самки — 84; 1927 г. Самцы — 78, Самки — 86.
Средний вес тысячи самцов 3.4 кг, самок 5.5 кг. У берегов Туркмения попадаются длиной до 15 см. Входящие в дельту Волги бывают возрастом от 1 до 4 лет; самцы половозрелы в возрасте уже одного года, самки — в возрасте двух лет; преобладают трехгодовалые и четырехгодовалые. Средние размеры годовиков 57—60 мм, двухгодовалых 79—80 мм, трехгодовалых 91—95 мм, четырехгодовалых 110 мм (Киселевич, 1937). Икрометание порционное; в среднем самка выметывает около 30 тыс. икринок (Ловецкая). После икрометания часть взрослых скатывается в море, часть остается в ильменях, откуда уходит осенью; иногда, по словам Киселевича (1926), они остаются в ильменях до весны, причем приобретают облик чархальской тюльки (Clupeonella tscharchalensis). Е. К. Суворов находил в 1913 г. в северной части Каспия половозрелых тюлек с 8 апреля (н. ст.) и до 19 июня; у Челекена и у Астары половозрелые самцы и самки встречены 14 октября и 2 ноября 1913 г. н. ст. По словам же Сергеевой и Ганешиной, пелагическая икра и личинки этой сельди встречаются в южн. Каспии, а также частью и в восточной половине южного Каспия в течение осени, всей зимы и весны. Возможно, то мы имеем здесь дело с весенними и осенними формами Harengula delicatula, подобно тому как это известно для Clupeonella harengus.
Итак, каспийская тюлька может метать икру и в пресной воде, и в соленой (до 15 pro mille).
Южнокаспийские тюльки становятся половозрелыми или в возрасте меньше года или в начале второго года; самцы длиной около 5 см уже нерестятся (Сергеева и Ганешина). 9 апреля было произведено на Брянской косе искусственное оплодотворение; мальки длиной 1.25 мм выклюнулись спустя 92 часа по оплодотворении при температуре воды в 8—10°.
Каспийская тюлька держится обычно на глубине от 6 до 30 м, иногда глубже. Зимою и летом собирается на глубинах от 8 до 30 м. Весною поднимается выше, иногда почти до поверхности. Из всех тюлек, этот вид наиболее приспособлен к перенесению низких температур: он встречается при 3°, а возможно и при более низких. Осенью, в связй с похолоданием, косяки каспийской тюльки передвигаются на юг.
По мнению А. А. Ловецкой (1940, стр. 29), обыкновенная каспийская тюлька состоит не менее чем из двух стад. Одно из них, проведя зиму в южном Каспии (в его северной половине, а возможно, в южной части среднего Каспия), в начале весны начинает двигаться на север, преимущественно вдоль западного побережья среднего Каспия, направляясь для нереста в северный Каспий, откуда часть этого стада заходит в низовья Волги и других рек, где и происходит икрометание; частью же нерест совершается в самом море, в опресненных, предустьевых частях. Второе стадо, повидимому, всю жизнь проводит в южном Каспии, совершая сезонные миграции в его пределах; в конце зимы и весною эта южная тюлька подходит к западным и восточным берегам южного Каспия.
В Каспийском море тюлька весною громадными косяками приваливает к берегу. У берегов Дагестана весенний ход обычно начинается в конце марта — начале апреля, при температуре воды 5—8°, и продолжается около месяца. В 1927 г. максимальный ход был в двадцатых числах апреля. Наибольшее количество вылавливаемой у берегов Дагестана тюльки приходится на долю трехгодовалых; предельный возраст 5 лет; длина улавливаемой здесь тюльки 53—133 мм. Средний размер самцов 91 мм, самок 103 мм (до конца средних лучей С). Средний вес тысячи самцов 7290 г, тысячи самок 9030 г, тысячи особей обоего пола 8720 г, максимальный вес одной особи 15.5 г (весна 1928 г. у берегов Дагестана). Таким образом, в уловах у берегов Дагестана тюльки попадаются крупнее и старше ловимых в дельте Волги, что зависит от величины ячей орудий лова. Плодовитость 9.5—60 тыс. икринок, в среднем 31.2 тыс. (Дмитриев).
Как правило, в сев. Каспии не зимует, появляясь здесь лишь в начале апреля.
Промысловая рыба, идущая на приготовление консервов.
В 1940 г. в Каспийском море добыто 80.7 тыс. ц тюльки (В уловы Clupeonella delicatula, очевидно, включены и уловы Clupeonella engrauliformis.), из них у берегов Дагестана 50.6 тыс., Азербайджана 15.2 тыс. и Казахстана 11.3 тыс. (Ловецкая, 1941).
В Таганрогском заливе главный лов тюльки, как здесь называют Clupeonella delicatula, происходит с апреля по июнь, когда она в большом количестве подходит к берегам для нереста и частью входит в дельту Дона.
Нерестует она также в лиманах Ейском и Миусском. Особи с текучими половыми продуктами попадаются с конца апреля до августа (Телегин). Весною 1930 г. в Азовском море выловлено 40000 ц тюльки размерами 6—7 см. Экземпляры из Бердянска (апрель—июнь 1923 г.) имели в длину: самцы до 80 мм (в среднем 64), самки до 90 мм (в среднем 72) (как измерялось, не указано; повидимому, до конца средних лучей С). У Белосарайской косы икра и личинки тюльки встречаются с мая по август; повидимому, нерест продолжается и в сентябре. К северным берегам Азовского моря тюлька подходит с апреля по середину мая, когда ее ловят массами; главный лов происходит на Бердянской косе (Тихонов). В Днепровском лимане встречается массами и под Очаковым составляет предмет промысла. В 1938 г. в Азовском море добыто 793.8 тыс. ц тюльки, в Черном 22.5 тыс. ц.
Необходимы биометрическое изучение и сравнение каспийской и черноморской тюлек.
Прибавление. А. Н. Световидов (Зоол. журн., 1943, стр. 231; Докл. Акад. Наук, XLVI, № 5, 1945, стр. 227) выделяет каспийскую тюльку в особый подвид caspia Svetovidov, который отличается большей величиной (обычно 80—100 мм до конца средних лучей С, вместо
40— 70 мм, как обычно в Азовском море), более короткими грудными (в среднем 17%, вместо 19%) и брюшными (9.5% вместо 12.7%) плавниками, менее высоким телом (20.6% вместо 24.4%). О миграции каспийской тюльки у берегов Мангышлака и в сев. Каспии сообщает Е. Н. Казанчеев в Тр. Волго-касп. ст., IX, вып. 1, 1947, стр. 38—62.