Limanda aspera (Pallas, 1814)
Желтопёрая камбала, желтопёрая лиманда, колючая лиманда (Rus),
Yellowfin sole
(Eng)
Синонимы:
Limanda asprella Hubbs, 1915
Pleuronectes asper Pallas, 1814
Атлас-определитель рыб: Книга для учащихся. Н.А.Мягков 1994 г.
Камбала желтоперая — Limanda aspera (Pallas, 1811)
Тело высокое. Боковая линия сильно изогнута над грудными плавниками. Рыло очень короткое, его длина не превышает диаметра глаза. D 66—75. А 50—61. В боковой линии 80—89 чешуй. Окраска похожа на таковую Limanda limanda, но спинной и анальный плавники со слепой стороны интенсивно-желтые. Длина тела не превышает 45 см. Ареал охватывает воды Дальнего Востока от Берингова пролива до Кореи.
Рыбы Японского моря и сопредельных частей Охотского и Желтого морей. Часть 6. (Pleuronectiformes). Г.У.Линдберг, В.В.Федоров 1993 г.
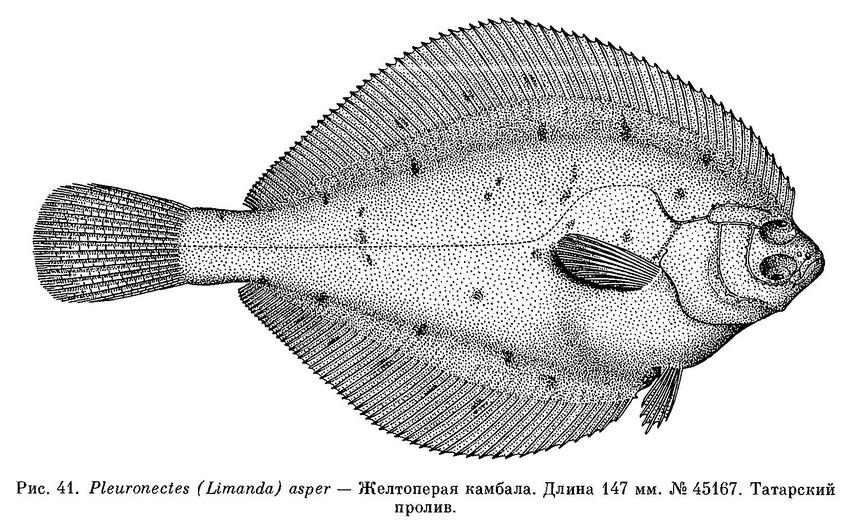
2. Pleuronectes (Limanda) asper Pallas, 1814 — Желтоперая камбала (лиманда) (рис. 41).
Pleuronectes asper Pallas, Zoogr. Rosso-Asiat, 3; [1814] : 425 (Восточный океан). — Günther, Cat. Fish, 4, 1862 : 454 (по Pallas, 1814). — Steindachner, Kner, Sitz. Ber. Akad. Wiss. Wien, 61, 1, 1870 : 425. — Sakamoto, Mem. Fac. Fish. Hokkaido Univ., 31, 1—2, 1984 : 208 (положение в системе).
Limanda aspera, Bean, Proc. U. S. Nat. Mus., 4, 1881 : 242. — Jordan, Goss. Rep. U. S. Comm. Fish., 14, 1889 : 288, pl. 5, fig. 13. — Jordan, Evermann, Bull. U. S. Nat. Mus., 47, 3, 1898 : 2645, pl. 377, fig. 930 (синонимия, описание). — Шмидт, Рыбы восточных морей..., 1904 : 233, рис. 16 (ЗИН 12356, 12357, 13058—13062, 8731). — Jordan, Starks, Proc. U. S. Nat. Mus., 31, 1906a : 209 (синонимия, описание). — Грацианов, Опыт обзора рыб..., 1907 : 447 (синонимия). — Tanaka, Annot. Zool. Jap., 6, 4, 1908а : 254 (Южный Сахалин). — Павленко, Рыбы залива Петр Великий..., 1910 : 57. — Gilbert, Burke, Bull. U. S. Bur. Fish., 30, 1912 : 15. — Jordan, Tanaka, Snyder, J. Coll. Sci. Imp. Univ. Tokyo, 33, 1, 1913 : 328 (Берингово и Охотское моря). — Hubbs, Proc. U. S. Nat. Mus., 48, 1915 : 478 (описание). — Mori, J. Pan-Pacif. Res. Inst., 3, 3, 1928 : 8. — Солдатов , Линдберг, Обзор рыб..., 1930 : 401 (частью: исключая Pleuronectes sakhalinensis. Охотское море, Татарский пролив, зал. Петра Великого). — Попов, Иссл. морей СССР, 14, 1931 : 146. — Шмидт, Изв. АН СССР, 12, 1931а : 319 (Пусань). — Rendahl, Ark. Zool., 22, 18, 1931b : 65 (частью: исключая Pleuronectes sakhalinensis). — Norman, Flatfishes (Heterosomata)..., 1934 : 336, fig. 247 (Берингово море, Аляска, Камчатка, Де-Кастри). — Тоwnsеnd, Rep. Internat. Fish. Comm., 11, 1936 : 10 (меристические признаки). — Андрияшев, Исслед. дальневост. морей, 5, 1937 : 325. — Моисеев, Изв. ТИНРО, 12, 1937б : 114 (меристические признаки). — Таранец, Краткий определитель..., 1937 : 144, рис. 87 (северная часть Тихого океана на юг до Ванкувера и п-ова Корея). — Okada, Matsubara, Keys Fishes Jap., 1938 : 430. — Kuronuma, Bull. Biogeogr. Soc. Jap., 13, 16, 1943 : 116 (сравнение с Pleuronectes sakhalinensis). — Линдберг, Изв. ТИНРО, 25, 1947 : 198 (частью: исключая Pleuronectes sakhalinensis). — Шмидт, Желтоперая камбала в: «Промысловые рыбы СССР», 1949 : 714, табл. 222. — Шмидт, Рыбы Охотского моря, 1950 : 231, рис. 24 (Охотское и Японское моря). — Андрияшев, Рыбы северных морей СССР, 1954 : 490, рис. 284 (синонимия, описание). — Matsubara, Fish morph. a. hier., 2, 1955 : 1271 (Хоккайдо, Сахалин, Охотское море, Камчатка, Аляска, Корея). — Walters, Bull. Amer. Mus. Nat. Hist., 106, 5, 1955 : 322. — Сhyung Moon Ki, Fishes Korea, 1977 : 573, col. pl. 128, 3. — Masuda et al., Fish. Jap. Arch., 1984 : 337, pl. 316—A,B.
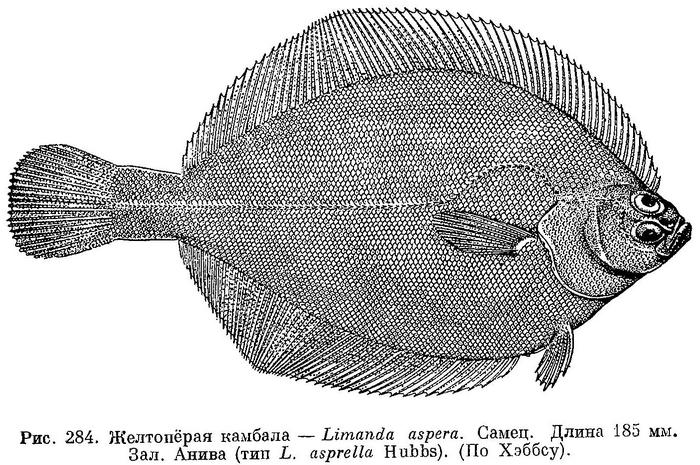
Limanda asprella Hubbs, Proc. U. S. Nat. Mus., 48, 1915 : 482, pl. 27, fig. 7 (зал. Анива, Корсаков).
12356. Японское море, Холмск. 1 VI 1901. П. Ю. Шмидт. 1 экз.
12357. Японское море, р. Лютога, Углегорск. 2 VII 1901. П. Ю. Шмидт. 1 экз.
17817. Японское море, зал. Петра Великого, Уссурийский зал. 3 X 1912. ДВЭ. 1 экз.
17818. Японское море, Татарский прол., 51 °41'30" с. ш., 171 °16'00" в. д. 4 VII 1912. 6 экз.
17820. Японское море, зал. Петра Великого, бухта Патрокл. 3 X 1912. ДВЭ. 4 экз..
17833. Японское море, зал. Де-Кастри. 30 VIII 1913. ДВЭ. 6 экз.
17837. Японское море, Татарский прол., мыс Пильво, Западный Сахалин. 4 V 1913. ДВЭ. 1 экз.
17838. Японское море, Татарский прол., Ванино. 24 VIII 1911. ДВЭ. 2 экз.
17840. Японское море, Татарский прол., Ванино. 6 V 1913. ДВЭ. 1 экз.
17841. Японское море, Татарский прол., Советская гавань. 23 VIII 1911. ДВЭ. 2 экз.
23847. Японское море, Пусань. 1930. 4 экз.
25124. Японское море, 51 °28' с. ш, 140 °58' в. д. Д. И. Охрямкин. 2 экз.
26219. Японское море, бухта Киевка. 17 IX 1934. Эксп. ЗИН в Японское море. 1 экз.
43699. Японское море, Татарский прол. 18 VI 1910. Ф. А. Дербек. 2 экз.
45167. Японское море, Татарский прол. 29 VIII 1915. М. Н. Павленко. 1 экз.
45169. Японское море, зал. Петра Великого. Широкая Падь. 20 VIII 1933. А. Кузнецов. 1 экз.
Охотское море: 13058—13062, 17815, 17819, 17825, 17827—17829, 17831, 17832, 17834, 17836, 17939, 19076, 19080, 19887, 25123, 31659, 41268, 44672, 44673, 46888, 46890—46895, 46900, 46997, 46998, 47000—47003, 47058.
Курильские о-ва: 45170, 46889, 46897—46899, 46996.
Юго-восточная Камчатка: 19075, 19078, 20687, 33505, 34973, 43745, 45166, 45168, 45171, 46896, 46999, 47004.
Берингово море: 19079, 33407, 33503, 33504.
Северная Америка: 8731, 23701.
D 67; А 48; Р 12; V 6; С 18 (12 ветвистых) (Pallas, 1814 : 425).
D 68—72; А 53—55; l.l. 83—85 (Шмидт, 1904 : 233).
D 68—77; А 53—58; Р 11; V 6; l.l. 80—90; пор в подглазничном канале ниже нижнего глаза 18—20; жаберных тычинок 5—7 + 8—10 = 13—17 (Hubbs, 1915 : 478).
D 68—74; А 48—56; Р 10—12 (6—8 ветвистых); l.l. 74—81, выше боковой линии 25—31 ряд чешуи; жаберных тычинок ? + 8—10; зубов 7—12 + 17—29 / 9—14 + 18—25 (Norman, 1934 : 336).
D (61) 66—71 (74), ср. 68.4; А (49) 50—54 (58), ср. 52.6; С (17) 18 (19); позвонков 10—11 + 28—29 = 39—40, ср. 39.4 (Townsend, 1936 : 10).
D (65) 67—72 (75), ср. 70.01; А (50) 51—56 (58), ср. 53.77; Р (10) 11 (12), ср. 10.99; l.l. (80) 81—85 (87), ср. 83.00; жаберных тычинок (12) 14—15 (18), ср. 14.86; позвонков 10 + (29) 30—31, ср. 30.37 = (39) 40—41, ср. 40.37 (Моисеев, 1937б : 144).
По нашим материалам (82 экз. длиной 59—420 мм): D (63) 65—75 (78), ср. 69.76; А (48) 50—59 (61), ср. 55.23; Р (10) 11—12 (13), ср. 11.45 / (9) 11—12, ср. 11.21; С (2) 3 (4) + (5) 6 (7) + (5) 6 + (2) 3 (4) = (17) 18 (19), ср. 17.97; l.l. (72) 78—87 (95), ср. 80.28, выше боковой линии (23) 25—29 (33), ср. 28.73, ниже — (26) 28—31 (38), ср. 30.0 рядов чешуи; пор в подглазничном канале ниже нижнего глаза 10—17, ср. 14.0, выше верхнего глаза — 6—12, ср. 8.9; жаберных тычинок (4) 5—7 (9), ср. 6.15 + (7) 8—10, ср. 8.94 = (12) 14—17 (18), ср. 15.09; зубов (4) 7—11 (16), ср. 8.92 + (15) 17—24 (30), ср. 21.17 = (22) 27—33 (44), ср. 30.08 / (5) 8—14 (22), ср. 12.50 + 15—24, ср. 20.00 = 23—36 (44), ср. 32.01.
Длина головы 24.66, антеанальное расстояние 27.51, длина хвостового плавника 23.29 % SL (Pallas, 1814 : 425).
Длина головы 22.73—25.0, высота тела 43.48—47.62 % SL, диаметр глаза 16.67, межглазничное расстояние 8.33 % lс (Шмидт, 1904 : 233).
По нашим материалам (25 экз. длиной 59—247 мм) в процентах стандартной длины: длина головы 23.9—27.6, ср. 26.20; наибольшая высота тела 40.7—50.0, ср. 45.82; высота хвостового стебля 9.3—12.2, ср. 11.06; длина хвостового стебля 6.6—10.6, ср. 8.46.
Pleuronectes asper морфологически очень близок к Pleuronectes sakhalinensis: все пластические и счетные признаки у этих видов перекрываются в той или иной степени. Наиболее надежный и пока единственный признак, по которому можно сразу же различить эти виды — число ктений на чешуе. У Pleuronectes asper на каждой чешуе имеется от 1 до 3 грубых, торчащих в сторону ктений; у Pleuronectes sakhalinensis — 4—10 утонченных, направленных назад ктений. Поэтому Pleuronectes asper на ощупь более грубый, чем Pleuronectes sakhalinensis.
Проведено сравнение электрофоретических спектров белков и выявлен биохимический полиморфизм популяции Pleuronectes asper в зал. Петра Великого (Коваль, Богданов, 1978, 1979, 1982). Исследованы биохимические варианты изоцитратдегидрогеназы, лактатдегидрогеназы, а-глицерофосфатдегидрогеназы, тетразолиумоксидазы, фосфоглюкомутазы, аспартатаминотрансферазы, глицеральдегид-3-фосфатдегидрогеназы, фосфогексоизомеразы, малатдегидрогеназы, пептидазы и общего белка мышц, которые могут быть полезны при решении вопросов о разделении различных линий рыб, их гибридизации и происхождении (Johnson, 1977). Биохимические показатели даны для технологических характеристик (Кривобок, Тарковская, 1964), обмена веществ (Рассказов и др., 1967) и биологически активных веществ (Степанова, 1971).
Мускулатура интенсивно заражена личинками трематод рода Stephanostomum (Малышева, Коротаева, 1987).
Распределение желтоперой камбалы в промысловых количествах носит прерывистый, мозаичный характер. Существует несколько географически изолированных, локальных стад различной численности. Сопоставление распределения Pleuronectes asper с рельефом дна, наличием подводных долин и каньонов свидетельствует, что промысловые скопления приурочены к хорошо развитым системам подводных долин и каньонов. Наиболее многочисленные популяции и наиболее интенсивно эксплуатируемые промыслом располагаются в Японском море в зал. Петра Великого, у берегов Приморья и о. Сахалин в Татарском прол., в Охотском море в зал. Терпения и у берегов Юго-Западной Камчатки, в Тихом океане в Кроноцком и Камчатском заливах у берегов Юго-Восточной Камчатки, в Беринговом море в юго-восточной части. Во всех этих районах в результате увеличения интенсивности промысла произошли значительные изменения в численности, распределении, размерно-возрастном составе, сроках миграции и поведении отдельных видов камбал и соотношения их в общих уловах. Отмечено появление вертикальных суточных миграций, начала активного питания зимой. Все это сказалось на эффективности промысла, что привело к снижению его интенсивности. Однако резкое снижение вылова, а в некоторых районах полное его прекращение, к восстановлению прежних запасов камбал пока не привело.
Дополнительные сведения о промысле и промысловых запасах можно найти преимущественно в работах отечественных авторов (Павленко, 1920; Линдберг, 1927; Охрямкин, 1930; Кривобок, 1931; Охрямкин и др., 1936; Моисеев, 1937а, 1937б, 1940, 1946а, 1946б, 1946в, 1953а; Таранец, 1938; Гавриленко, Моисеев, 1939; Тычкова, 1946; Гордеев, 1949; Суворов, 1951; Полутов, Пашкеев, 1955, 1967; Фадеев, 1957, 1960, 1961а, 1961б, 1962, 1963а, 1963б, 1965а, 1965б, 1965в, 1965г, 1968, 1970а, 1970б, 1970в, 1970г, 1970д, 1970е, 1971а, 1971б, 1971в, 1984, 1987; Ломаев, 1958; Полутов, 1958, 1960, 1967; Полутов, Васильев, 1959; Полутов, Тихонов, 1959; Меркулов, 1962; Минева, 1964, 1967, 1970, 1971а, 1971б, 1972; Тихонов, 1965, 1966, 1968а, 1968б, 1969, 1970, 1975, 1976, 1977, 1981, 1984; Полутов и др., 1966; Власова, 1967а, 1967б; Иванкова, 1967, 1968; Ковтун, 1968; Alverson, Pereyra, 1969; Власова и др., 1971; Smith, 1974; Власова, 1979; Baccala, 1979; Baccala et al., 1980; Wakabayashi, 1984).
Длина 490 мм, масса 1.79 кг (Фадеев, 1987 : 115).
Распространение. В Японском море известен вдоль материковых берегов у Пусаня (Mori, 1928 : 9; Шмидт, 1931а : 317), Сончжинь (Mori, Uchida, 1938 : 23), Чхончжинь (Mori, 1952 : 181), в зал. Петра Великого (Павленко, 1910 : 57; Линдберг, 1947 : 198), вдоль Приморья (Охрямкин и др., 1936 : 16) и далее до северной части Татарского прол. (Солдатов, Линдберг, 1930 : 401); вдоль островных берегов у о-вов Хоккайдо (Ueno, 1971 : 99) и Сахалин (Николаев и др., 1958 : 5). В Охотском море повсеместно на материковом шельфе (Шмидт, 1904 : 233, 1950 : 231; Попов, 1931 : 146; Таранец, 1937а : 144), заходит в Амурский лиман (№№ 13060, 17891) и ловился в Сахалинском зал. (№№ 17819, 17825, 17832, 17834, 17836, 17839) и у Шантарских о-вов (Н. С. Фадеев (1987 : 56) сообщает, что Pleuronectes asper отсутствует у Шантарских о-вов.) (№№ 17815, 17828, 17829, 44672, 44673), известен у берегов о. Хоккайдо (Ueno, 1971 : 99), в том числе в оз. Ноторо (Hikita, 1952а : 1,5). До сих пор не пойман в средней части Курильской гряды (Фадеев, 1987 : 56). У южных Курильских о-вов малочислен (Линдберг, 1959 : 256) и является объектом промысла у северных (Легеза, 1959 : 280). В Желтом море неизвестен. У тихоокеанских берегов Японии только вдоль о. Хоккайдо (Ueno, 1971 : 99). Многочислен вдоль берегов Юго-Восточной Камчатки (Полутов, 1960 : 31) и в Беринговом море (Андрияшев, 1937 : 325, 1952 : 320, 1954 : 490), у Командорских о-вов (Гребницкий, 1897 : 324). В Чукотском море (Андрияшев, 1954 : 491; Pruter, Alverson, 1962 : 82; Quast, 1972 : 205; Quast, Hall, 1973 : 38; Wolotira et al., 1977 : 250). У берегов Северной Америки от зал. Аляска (Wilimovsky, 1954 : 27) на юг до зал. Беркли в Британской Колумбии (Eschmeyer et al., 1983 : 290).
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
Особенно многочисленна и широко
распространена тихоокеанская желтоперая
камбала (Limanda aspera), встречающаяся от
берегов Кореи до Чукотского моря и
по тихоокеанскому побережью Северной
Америки на юг до о-ва Королевы
Шарлотты. Желтоперая камбала может
достигать длины 48 см, средняя длина ее 28—35 см. На глазной стороне у нее вдоль
оснований спинного и анального
плавников проходит узкая черная полоска.
Слепая сторона белая, спинной и
анальный плавники снизу желтые с
голубовато-серым краем и белыми концами лучей.
Держится у берегов на глубине от 15
до 80 м.
Это одна из важнейших промысловых
камбал Дальнего Востока. Она является
основой наиболее крупных скоплений
камбал у берегов Западной Камчатки,
в Олюторском заливе, Татарском проливе
и заливе Петра Великого. Большие
скопления желтоперой камбалы имеются и в
восточной части Берингова моря.
По вкусовым качествам это одна из
лучших тихоокеанских камбал.
Рыбы северных морей СССР. А.П.Андрияшев 1954 г.
2. Limanda aspera (Pallas) — Желтопёрая камбала, колючая лиманда (рис. 284).
Pleuronectes asper Pallas, Zoogr. Rosso-Asiat., III, 1811 : 425 («ex oceano orientali»; повидимому, из сборов д-ра Мерка в Беринг. м.).
Limanda aspera Шмидт, Рыбы вост. морей, 1904 : 233, фиг. 16 (нижнеглоточные зубы). — Evermann a. Goldsborough, Fishes Alaska, 1907 : 354, fig. 138 (на север до зал. Коцебу). — Hubbs, Proc. U. S. Nat. Mus., LXVIII, 1915 : 478 (жаб. тычинок 13—17; близкие, если не тождественные, формы описаны здесь же как Limanda sakhalinenisis, Limanda asprella и Limanda korigarei). — Rendahl, Ark. Zool., Stockh., XXIIA, 18, 1931 : 65—74 (сравнение с другими видами). — Norman, 1934 : 336, fig. 247 (синонимия). — Охрямкин, Моисеев и Таранец, Промысл. камбалы Приморья, 1936 : 16, фиг. 7 (зал. Петра Великого, Татарск. прол.; биология). — Townsend, Rep. Int. Fisher. Comm. Seattle, 11, 1936 : 10 и сл. (меристические признаки). — Андрияшев, Иссл. морей СССР, 25, 1937 : 325 (Анадырск. зал., устье р. Юкон, к северо-востоку от о. Св. Лаврентия). — Моисеев, Изв. Тихоок. научн. инст. рыбн. хоз. океаногр., XII, 1937 : 144 (Японск. м.; меристические признаки). — Суворов и др., Учен. зап. Ленингр. Гос. унив., 15, 1937 : 241 (зап. Камчатка; размеры, возрастные группы). — Мищенко, Изв. Тихоок. научно-иссл. инст. рыбн. хоз. океаногр., XIV, 1938 : 171, фиг. 1 (развитие). — Моисеев, там же, XXII, 1946 : 75—184 (миграции, рост, плодовитость, промысел). — Моисеев, Промысл. камбалы Дальн. Вост., 1946 : 13 и сл. — Сlеmens a. Wilby, Fish. Pacif. Coast Canada, 1946 : 323, fig. 242 (тихоокеанск. Канада). — Расс и Желтенкова, Изв. Тихоок. научно-иссл. инст. рыбн. хоз. океаногр., XXVIII, 1948 : 146 (зап. Камчатка; икра). — Шмидт в изд.: Промысл. рыбы СССР, 1949 : 714, атлас табл. 222 (сводка, цветн. рис.).
Limanda aspera. Новые данные о темпе роста желтопёрой камбалы сообщает М. А. Петрова-Тычкова (Изв. Тихоок. инст. рыбн. хоз. океаногр., XXXVII, 1952 : 254).
D 66—75, А 50—61, l.l. 80—89, жаберных тычинок 14—15 (в северной части Берингова моря и у вост. Камчатки); для 153 аляскинских экземпляров Тоунсенд приводит следующие данные: D 61—74, чаще 66—71 (среднее 68.4), А 49—58, чаще 50—54 (ср. 52.6), позвонков у 23 экз. 39—40. У 73 экз. из зал. Петра Великого по данным Моисеева (1937): D 65—75 (ср. 70.0), А 50—58 (ср. 53.8), пор в боковой линии 80—87 (ср. 83.0), позвонков (39) 40—41 (ср. 40.4).
Близка к Limanda limanda. Между боковой линией и краем середины спины 25—31 чешуя (Норман). Рыло короткое, не превышает диаметра верхнего глаза. Заглазничный гребень не развит. На голове простая надвисочная ветвь боковой линии. Узкая черная полоска, проходящая на глазной стороне вдоль основания спинного и анального плавников, выражена отчетливо. Слепая сторона тела белая, спинной и анальный плавники снизу желтые, с голубовато-серым краем и белыми кончиками лучей. Длина до 48 см; длина наибольшего из наших экземпляров 433 мм (бухта Моржовая на восточном берегу Камчатки). У западных берегов Камчатки средний размер траловой желтопёрой камбалы 34 см, средний вес 420 г (Моисеев, 1946).
Распространение. Северная часть Тихого океана. Японское (на юг до Пусана), Охотское и Берингово моря; на север доходит до Берингова пролива, единично до зал. Коцебу в юго-восточной части Чукотского моря. По американскому побережью спускается на юг до о. Королевы Шарлотты.
Биология. Зимует на глубине 150—250 м, но в апреле вслед за хоботной камбалой подходит для нереста к берегам, где держится до сентября. В Беринговом море желтопёрая камбала встречается в летнее время на глубине от 10 до 130 м, преимущественно при положительных придонных температурах. Основную пищу составляют полихеты, моллюски и офиуры.
Биология желтопёрой камбалы Берингова моря не изучена; довольно полные биологические данные имеются для желтопёрой камбалы Японского моря (зал. Петра Великого и Татарский пролив) и западного побережья Камчатки (Моисеев, Суворов, Мищенко, Демидова и др. — см. у Моисеева, 1946).
Хозяйственное значение. Одна из основных промысловых камбал дальневосточных морей; уловы ее составляют около 60% улова всех камбаловых у западных берегов Камчатки и около 90% в Татарском проливе. Общий улов всех камбал на Дальнем Востоке составляет, по Моисееву (1946), в послевоенные годы, около 200—220 тыс. ц (1943—1944 гг.).