(Osteichthyes (Huxley, 1880)) Надкласс Остеихтиесы, Костные рыбы, Superclass Osteichthyes (Huxley, 1880) (Bony fishes) 2 класса
Материал подготовил Фактистов И. Е., 2020 г. Сайт www.fishbiosystem.ru
Костные рыбы (лат. Osteichthyes) — группа классов в надклассе Рыбы (Pisces). Ранее считался классом, в современной систематике обычно трактуется как объединение двух сестринских групп: классов Лучепёрые (Actinopterygii Klein, 1885) и Лопастепёрые (Sarcopterygii Romer, 1955).
Древнейшие известные окаменелости костных рыб относятся к лудловской эпохе силурийского периода (около 425 млн лет назад). Сейчас спорно, от кого они вместе с хрящевыми произошли: от плакодерм или акантод. После девонского вымирания, когда вымерли плакодермы, в морях стали господствовать костные и хрящевые рыбы, не имея конкурентов.
В зоологической литературе XX века группа костных рыб обычно рассматривалась как класс, состоящий из двух подклассов: Лучепёрые (Actinopterygii) и Лопастепёрые (Sarcopterygii). Такая трактовка сохраняется и в некоторых учебниках начала XXI века, однако всё большее распространение получают классификации, в которых ранг упомянутых подклассов повышен до ранга класса. Например, в классификации канадского зоолога Дж. С. Нельсона (2006) группа костных рыб (выступающая под названием Euteleostomi и не имеющая ранга) трактуется как объединение двух сестринских групп — классов Лучепёрые и Лопастепёрые. При этом если лучепёрые являются монофилетической группой, то лопастепёрые — группа заведомо парафилетическая (поскольку ближайший общий предок последней группы также является и предком современных четвероногих); в результате парафилетической оказывается и вся группа костных рыб. Следуя правилам кладистики, не допускающим использования парафилетических таксонов, Нельсон переопределяет объём лопастепёрых, включая в него и всех четвероногих (получивших в его классификации ранг инфракласса).
Рыбы открытого океана. Н.В.Парин 1998 г.
Рыбы — наиболее многообразная группа позвоночных животных, которая насчитывает более 20 тыс. видов, в большинстве своем относящихся к классу костных рыб, самых высокоорганизованных первичноводных позвоночных. Эта цифра превышает общее количество земноводных, пресмыкающихся, птиц и млекопитающих: их известно только 18 тыс. видов. При этом следует иметь в виду, что «инвентаризация» рыб еще далеко не закончена и ежегодно в специальной литературе публикуются десятки описаний новых видов, главным образом населяющих большие глубины Мирового океана и малоизученные реки в сельве и джунглях тропической зоны.
Такое многообразие форм — результат длительного эволюционного процесса, позволившего рыбам освоить почти все участки водной среды обитания в океанах, морях и внутренних водоемах нашей планеты. Действительно, рыб можно встретить в прозрачных ключах и в мутных болотах, в горных озерах на высоте 6000 м над уровнем моря и в мрачных подземных пещерах. В морских водах они обитают от зоны волнового заплеска до глубoчайших океанических впадин. Рекордные по глубине нахождения принадлежат семействам морских слизней (Liparididae) и ошибневых (Ophidiidae), которые были обнаружены на глубине около 7600 и 8300 м, и есть все основания считать, что это еще не предел — рыбы, по-видимому, могут встречаться и на дне более глубоких желобов (при спуске на батискафе «Триест» в Марианском желобе Ж. Пикар и Д. Уолш наблюдали и фотографировали какое-то камбалоподобное живое существо на глубине более 10000 м, но анализ их снимков специалистами к сожалению, не подтвердил принадлежности этого объекта к рыбам).
Представители этой группы животных живут в толще воды, на дне и даже в верхнем слое грунта, а отдельные виды частично используют и воздушную среду. В Арктике и Антарктике рыбы населяют воды с отрицательной температурой (до —2°С), близкой к точке замерзания соленой воды, а в горячих термальных источниках Калифорнии благоденствуют даже при 50-градусной температуре. Максимальная соленость воды, которую переносят некоторые виды, достигает 60—80‰ — вдвое выше «нормы» для открытого океана.
Внешний облик рыб прекрасно соответствует их образу жизни. Как разительно непохожи живущая среди залитых тропическим солнцем коралловых рифов яркая и пестро окрашенная рыба-бабочка и обитатель мрачных океанических глубин черный и зубастый, похожий на надутый пузырь удильщик или змееподобный угорь и дисковидная, состоящая как бы из одной только головы рыба-луна! Тонкие приспособления рыб к условиям среды их обитания не могут не вызвать естественного изумления у человека, который знакомится с поразительным многообразием этой группы животных.
С давних времен и до наших дней лов рыбы имеет первостепенное хозяйственное значение в жизни и экономике многих народов. Биологические (и прежде всего рыбные) ресурсы морей и внутренних водоемов все более полно используются человеком для удовлетворения растущих потребностей в белковой пище. Во многих странах, расположенных на берегах океанов (Японии, Вьетнаме, Индонезии и др.), рыба и другие продукты моря составляют основу белкового рациона населения. Нынешний мировой улов выражается огромной цифрой — около 85 млн т в год, т. е. примерно 17 кг в пересчете на одного жителя Земли. Список промысловых объектов включает сотни названий, но особенно большое место в рыболовстве занимают морские рыбы шести семейств — сельдевые, тресковые, анчоусовые, скумбриевые, ставридовые и камбаловые, на долю которых приходится около половины общего вылова.
В этой книге речь пойдет о рыбах, населяющих только воды открытого океана, той части водной оболочки Земли, которая находится вне границ прибрежной зоны. Естественно, сразу возникает вопрос, где именно начинается этот самый «открытый океан». В современной биологической океанологии и рыбохозяйственной науке это словосочетание употребляется сейчас в двух значениях. Во-первых, так обозначается часть Мирового океана, находящаяся за пределами 200-мильных экономических зон, повсеместно введенных прибрежными государствами в 1976—1978 гг. и находящихся под их юрисдикцией, во-вторых, воды, лежащие мористее внешнего края материковой отмели (шельфа).
Таким образом, океанические рыбы обитают вне шельфов и надшельфовой (иначе говоря, неритической) пелагиали. По такому определению в эту категорию попадают как пелагические (т. е. живущие в толще воды), так и бентические (донные) рыбы, а также бенто-пелагические (придонные и наддонные) рыбы, занимающие промежуточное положение между первыми двумя группами. Вертикальное распределение океанических рыб охватывает весь диапазон глубин Мирового океана — от поверхности до дна самых глубоких желобов.
Впервые океанические рыбы (среди них летучие рыбы, тунцы, меч-рыба, рыба-луна и др.) были описаны в 1758 г. основоположником систематической зоологии Карлом Линнеем. В конце XVIII — начале XIX в. состоялось несколько хорошо известных кругосветных экспедиций — плавания капитанов Дж. Кука, Ж. Ф. Лаперуза, И. Ф. Крузенштерна и др., в ходе которых были собраны значительные биологические коллекции, включавшие и рыб приповерхностного слоя. Начало изучению глубоководных рыб было положено в первой половине XIX столетия трудами француза Антуана Риссо, итальянца Анастасио Кокко, англичанина Ричарда Лоу, которые дали описания ряда видов по экземплярам, обнаруженным мертвыми на поверхности воды, выброшенным на пляжи или пойманным крючковой снастью, опущенной на большую глубину.
Новую эпоху в изучении рыб открытого океана — период комплексных океанографических экспедиций — открыло в 1872 г. плавание английского корвета «Челленджер», специально переоборудованного для такого рода исследований. Ихтиологические сборы экспедиции (около 280 видов океанических рыб) были обработаны Альбертом Гюнтером, опубликовавшим специальные монографии по глубоководным и пелагическим рыбам, не утратившие своего значения и в настоящее время. В период, предшествовавший первой мировой войне, значительный вклад в изучение рыб открытого океана внесли также экспедиции на судах «Блейк», «Альбатрос», «Травайер», «Талисман», «Инвестигейтор», «Ирондель», «Принцесс Алис», «Вальдивия», «Сибога», «Михаэль Саре», в обработке материалов которых участвовали известные ихтиологи Д. Гуд и Т. Вин (первыми употребившие в 1896 г. термин «океаническая ихтиология»), Ч. Гилберт, А. Элкок, Л. Вайян, Р. Коллетт, С. Гарман, Д. С. Джордан, А. Брауэр, М. Вебер и др. Как ни прискорбно, в приведенных списках нет ни русских названий судов, ни русских фамилий — в те годы Россия не участвовала в накоплении сведений по океаническим рыбам. Итоги этого этапа исследований были подведены Джоном Мюрреем и Юханом Йортом в увидевшем свет в 1912 г. очень солидном труде «Глубины океана», в котором рассматривается и дно и вся толща вод океана от поверхности до грунта с ее характерным населением.
После перерыва, вызванного военными действиями, биологические изыскания в Мировом океане получили новое развитие. Среди многих исследовательских плаваний того времени следует особо выделить датские экспедиции на «Дане» и английские на «Дискавери» — под каждым из этих названий фактически существовало по два экспедиционных судна. На «Дане I» в 1920—1922 гг. были проведены под руководством И. Шмидта работы в Атлантическом океане, приведшие к обнаружению нерестилищ европейского угря в Саргассовом море. Кругосветное плавание «Даны II» (1928—1930 гг.), маршрут которой был проложен через тропические воды всех океанов, позволило получить огромные ихтиологические коллекции. Экспедиции «Дискавери I» (1929—1931 гг.) и «Дискавери II» (1932—1933 гг.) проходили преимущественно в Южном океане (вплоть до 60° ю. ш.) и много способствовали познанию природы и живого населения этого малоизученного региона. Большой вклад в изучение океанических рыб был внесен в рассматриваемый период такими учеными, как Т. Ригэн, Д. Нормен, В. Тонинг, В. Эге, А. Парр, Р. Болин, А. Бруун, Ч. Бредер, К. Хаббз, К. Матсубара, У. Чепмен, У. Данкона. К этому же времени относятся первые наблюдения океанических рыб непосредственно в среде их обитания: в 1934 г. американский ихтиолог У. Биб опустился в батисфере «Век прогресса» — стальном шаре диаметром 1,3 м с небольшим смотровым иллюминатором — на глубину более 900 м в районе Бермудских островов. Он и его спутник О. Бартон были первыми людьми, которым удалось увидеть довольно разнообразных глубоководных рыб (в том числе и поныне не пойманных) в их родной стихии.
В 30-е годы в дальневосточных морях и морях Северного Ледовитого океана были начаты и советские исследования глубоководных рыб. Основоположником отечественной океанической ихтиологии с полным основанием можно считать А. П. Андрияшева, которым еще в довоенные годы были сделаны очень важные обобщения, касающиеся происхождения глубоководной ихтиофауны.
В изучении рыб, обитающих за пределами шельфа, участвовали также Н. М. Книпович, П. Ю. Шмидт, В. К. Солдатов, А. Я. Таранец, А. Н. Световидов, Г. У. Линдберг.
С началом второй мировой войны биоокеанологические и ихтиологические исследования во всех океанах почти полностью прекратились, но вскоре после ее окончания были продолжены на новом, более высоком техническом уровне. Важный вклад в эти работы был сделан научно-исследовательским судном (сокращенно НИС) «Витязь», принадлежавшим Институту океанологии им. П. П. Ширшова АН СССР и совершившим в 1949— 1979 гг. 65 экспедиционных рейсов в Тихий, Индийский и Атлантический океаны. Ихтиологические исследования входили в программу 28 его плаваний. Сначала они были направлены на изучение дальневосточных морей и прилегающих районов Тихого океана, а с 1955 г. основное внимание было перенесено на исследование глубин и пеолагиали открытых вод. Работы, выполненные в плаваниях «Витязя», обеспечили быстрое развитие советской океанической ихтиологии, завоевавшей широкое международное признание. Они были начаты под руководством Т. С. Расса и осуществлялись в основном силами сотрудников Института океанологии (II. В. Парии, В. А. Мухачева, Н. Н. Горбунова, В. Э. Беккер, В. М. Макушок, Н. В. Ковалевская, Ю. Н. Щербачев и др.).
Сейчас исследовательский флот нашей страны насчитывает десятки разнотипных современных судов. В его состав входит и новый «Витязь», имеющий намного лучшие условия для работы и отдыха, чем на старом «Витязе». И тем не менее те, кому выпало счастье начать на этом судне свой путь в океанологию, всегда будут вспоминать его как «праздник, который всегда с тобой». Все рейсы «Витязя» (а мне довелось участвовать в его первом выходе в открытый океан, состоявшемся в 1955 г.) кажутся сейчас единственными и неповторимыми. Невозможно забыть радостный энтузиазм труда, царивший на борту и делавший любого участника комплексных экспедиций сопричастным к большим и маленьким открытиям, которые приносило каждое плавание. Именно поэтому все мы, «старые витязяне», так едины в своей привязанности к бывшему флагману советской научной флотилии и так близко принимаем к сердцу судьбу заслуженного ветерана, «временная» стоянка которого в калининградском порту слишком затянулась, так как планы его превращения в постоянный океанологический музей никак не воплотятся в жизнь.
Вклад экспедиций на «Витязе» в мировую ихтиологическую науку вряд ли может быть переоценен, и имя этого корабля, закрепленное в названиях одного нового рода рыб (Vitiaziella Rass, 1955) и девяти новых видов (Gonostoma vitiazi Rass, 1950; Lycenchelys vitiazi Andriashev, 1955; Pelagocyclus vitiazi Lindberg et Legesa, 1955; Cypselurus vitjazi Parin, 1958; Diaphus vitiazi Kulikova, 1961; Benthodesmus vitiazi Parin et Becker, 1970; Monomeropus vitiazi Nielsen, 1971; Eustomias vitiazi Parin et Pokhilskaya. 1974; Melanostigma vitiazi Parin, 1980), навсегда останется в анналах истории ихтиологии.
Одновременно с НИС «Витязь» проводили океанологические исследования, включавшие и сбор материалов по океаническим рыбам, другие советские исследовательские суда: «Обь» (1955—1958 гг.), «Академик Курчатов» (с 1967 г.), «Дмитрий Менделеев» (с 1969 г.). Продолжались экспедиционные работы иностранных государств, среди которых нельзя не упомянуть датскую глубоководную экспедицию на «Галатее» (1950—1952 гг.), американские исследования в Мексиканском заливе на «Орегоне» (1955 г.) и в Южном океане на «Элтенине» (с 1962 г.) и планомерные изыскания ученых ФРГ в Атлантическом океане на «Вальтере Гервиге I» (1964—1971 гг.) и «Вальтере Гервиге II» (с 1971 г.). Полный перечень ихтиологов, участвовавших в обработке собранных материалов, занял бы слишком много места — их число многократно увеличилось по сравнению с предвоенным временем. Особенно существенно продвинулось изучение фаунистического состава и систематики океанических рыб, в разработку которой внесли свой вклад ученые многих стран — СССР (А. П. Андрияшев, А. В. Балушкин, В. В. Барсуков, В. Э. Беккер, О. Д. Бородулина, В. Н. Долганов, А. Н. Котляр, В. А. Мухачева, А. В. Неелов, Н. В. Парии, Т. С. Расе, Ю. И. Сазонов, А. Н. Световидов, И. А. Трунов, В. В. Федоров, Ю. Н. Щербачев и др.), США (Р. Гиббс, Р. Джонсон, Т. Ивамото, Б. Коллетт, Л. Компаньо, Д. Коэн, Г. Мид, Б. Нафпактитис, Т. Питч, Р. Хэдрич, Д. Штейн, А. Эбелинг и др.), Японии (Т. Абэ, К. Амаока, Р. Исияма, И. Накамура, О. Окамура и др.), ФРГ (А. Коттхауз, Г. Креффт, А. Пост, М. Штемани), Франции (М. Л. Бошо, Ж.-К. Керо, П. Фурмануар, Ж.-К. Юро), Дании (Э. Бертельсеи, И. Нильсен), Великобритании (Д. Бедкок, Н. Меррит, Н. Маршалл), Новой Зеландии (Д. Гаррик, П. Касл), Португалии (Г. Мауль, Л. Салданья), Австралии (Д. Пэкстон), Норвегии (Е. Куфуд), Кубы (Д. Гитарт), Канады (Д. Нелсон). Некоторые из этих ихтиологов, а также многие не названные здесь внесли свой вклад и в изучение экологии рыб открытого океана.
Этот перечень разноязычных имен, без сомнения, покажется читателю малоинтересным, но я никак не могу обойтись без него в этой книге. Я смотрю на этот список, и перед моим мысленным взором предстают вполне конкретные результаты деятельности «поименованных лиц» - статьи и монографии, с которыми постоянно приходится иметь дело в повседневной работе на борту судна или в институтской лаборатории.
Помимо исследований рыб в океанологических экспедициях, не имевших, казалось бы, непосредственного отношения к промысловой тематике, в 50-е годы начались и быстро прогрессировали рыбохозяйственные исследования открытых вод Мирового океана. В нашей стране первым объектом этих изысканий стали «океаническая» сельди Северной Атлантики и сайра северо-западной части Тихого океана. В 60-е годы проводятся и научно-промысловыа исследования «полуглубоководных» рыб, живущих на материковом склоне. Эти работы (в их развитие вложили много труда П. А. Моисеев, Ю. Ю. Марти, В. П. Шунтов, Д. А. Шубников, Н. П. Новиков, Ю. В. Новиков, В. Л. Жаров и др.) выполняются в основном экспедициями бассейновых институтов и промысловых разведок Минрыбхоза СССР и характеризуются очень широким размахом. Суммарное число советских экспедиций в открытый океан уже превысило 2000.
Ихтиопланктон открытого океана включает не только ранние стадии развития океанических рыб (естественно, исключая щележаберных — акулы и скаты с самого начала жизни принадлежат к нектону), но и личинок и мальков множества шельфо-неритических видов, которые попадают в эпипелагиаль в результате пассивного выноса. В тропических водах его характеризует эфемерность существования — ведь развитие икры продолжается, как правило, не более 2 суток (правда, у макрелещуковых и летучих рыб — 2 недели, у вогмеровых рыб — до 2 месяцев), а личинки находятся в составе планктона в среднем около 8 недель. В более холодноводных районах эти сроки удлиняются, и длительность предмалькового периода жизни достигает у большинства видов 4—6 месяцев. Сильно растянуто личиночное развитие и у некоторых тепловодных форм, в частности у многих камбаловых и угреобразных рыб (личинки европейского угря, например, проводят в ихтиопланктоне до 3 лет), и это сильно способствует их дальнему разносу и широкому pacnpoстранению.
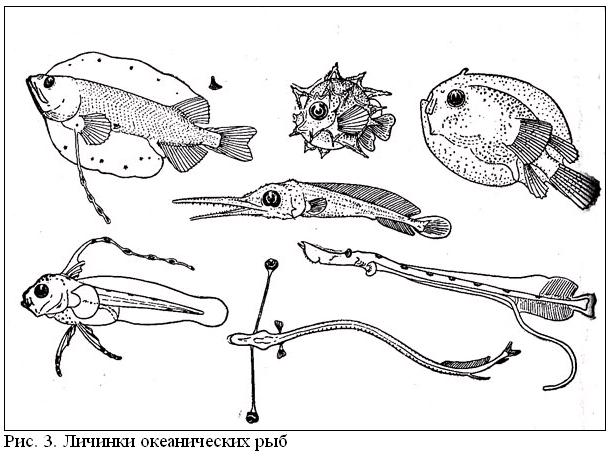
Икринки и личинки имеют приспособления к планктонному существованию, способствующие парению в толще воды: разного рода выросты и придатки на теле, удлиненные лучи плавников, а также жировые включения и газовые пузырьки (рисунок 3). Как правило, они сильно отличаются по всему своему облику от взрослых особей и в ряде групп претерпевают резкий метаморфоз (изменение строения) в ходе индивидуального развития. Поэтому установить принадлежность раннего малька, а тем более личинки (не говоря уже об икринке) к тому или иному виду, роду и даже семейству во многих случаях совсем непросто.
Особенно своеобразны прозрачные листовидные личинки угрей и близких к ним групп рыб. В прошлом столетии этих «рыбок» относили к особому роду Leptocephalus, и это название конечно, не в качестве родового осталось за ними до сих пор. Лептоцефалы некоторых видов достигают очень солидных размеров - рекорд (184 см) принадлежит L. giganteus (эта личинка относится к отряду спиношипообразных). Чтобы вырасти до такого размера, нужно, казалось бы, неплохо подкормиться, но, как ни странно, в кишечниках у этого гиганта или каких-либо других лептоцефалов никогда не находили даже следов пищи. Предполагают поэтому, что они могут усваивать растворенное в воде органическое вещество непосредственно через эпидермис. Согласно другому предположению, лептоцефалы высасывают протоплазматические соки из планктонных организмов, протыкая их покровы острыми зубами.
Мне никогда не приходилось всерьез заниматься изучением пхтиопланктопа (для этой исключительно тонкой работы нужны особые качества, которыми я не обладаю), хотя не раз доводилось иметь с ним дело, особенно в первых экспедициях — в те давно прошедшие времена одна из моих лаборантских обязанностей состояла в отборе икры и личинок рыб из планктонных проб. Эта нудная, как тогда казалось, работа (она и вправду очень однообразна) дала мне, однако, возможность воочию познакомиться с изумительным многообразием строения рыб на ранних стадиях развития, когда они так разительно непохожи на своих родителей. Я не могу забыть изумления и восхищения, которые мне доставляли тогда первые встречи с личинками листовидных совершенно прозрачных угрей, шарообразных колючих рыб-лун, стебельчато-глазых идиакантов, исключительно разнообразных по форме тела миктофид (взрослея, они становятся гораздо более схожими между собой), камбал, тунцов, рыб-топориков и прочими удивительными существами, которых так занимательно разглядывать под увеличением. С тех пор я сохраняю интерес к ихтиопланктону (мне даже удалось впервые определить видовую принадлежность нескольких неописанных личинок). Как и многие другие ихтиологи, я твердо убежден, что признаки ранних стадий рыб должны обязательно приниматься в расчет при изучении их систематики и филогении.
Итак, реальность существования и относительная обособленность планктона и нектона, несмотря на их не всегда четкое разграничение, не вызывают сомнений. Эти группировки представляют собой две крупные жизненные формы пелагического населения — пассивную и активную, из которых каждая по-своему хорошо приспособлена к обитанию в толще воды.
Форма тела нектонных рыб соответствует их активному образу жизни. Наиболее быстрым и подвижным хищникам — тунцам, акулам и др. — свойственна возникшая конвергентным путем торпедообразная форма тела. У некоторых рыб (меч-рыба, марлины) оно снабжено; спереди еще особым рострумом-обтекателем, который, как предполагает В. В. Овчинников, турбулизирует набегающий поток и, подобно выступу в подводной части носа современных быстроходных судов, сильно уменьшает лобовое сопротивление. Важную роль в обеспечении возможности быстрого движения имеют и хвостовые кили, увеличивающие поперечную жесткость хвостового стебля и выполняющие роль горизонтальных стабилизаторов. Для нектоирых рыб довольно обычно также симметрично сжатое с боков тело, в той или иной степени вытянутое в длину, или стреловидное тело с отставленными кзади спинным и анальным плавниками. В открытом океане минимальные размеры нектонных рыб могут, по-видимому, находиться в пределах от 15 до 20—30 см.
Планктонные рыбы обладают весьма ограниченными; возможностями для активного плавания на всех стадиях жизненного цикла. К этой группе принадлежат не только карликовые (длина 2—5 см) рыбки, населяющие глубокие слои пелагиали — циклотоны и другие представители семейства гоностомовых (Gonostomatidae), меламфайя мелкие светящиеся анчоусы и т. д., но и многие более крупные (длина до 30—50 см) малоактивные обитателя этого биотопа — нитехвостые угри, мешкороты, хаулиоды и др. К планктону (частично к нектопланктону) относятся и все рыбы, движущиеся при помощи ундулирующих, (колебательных) движений плавников — удильщики, рыбы-луны, сельдяные короли. А ведь некоторые из них; достигают очень внушительных размеров — нескольким метров. Такие рыбы почти не способны противостоять встречному движению воды, и это делает их объектом пассивного горизонтального переноса течениями.
В пространственном распределении всех организмов планктона (в том числе, естественно, и планктонных рыб) решающую роль играет пассивный перенос течениями. Нектонные животные активно перемещаются в продолжение своего жизненного цикла, мигрируя в пределах районов, характеризующихся необходимыми для вида в тот или иной период условиями существования. Это обстоятельство приводит к довольно важным различиям в функциональной структуре ареалов планктонных и нектонных рыб.
Ареалы нектона представляют собой районы, границы которых определяются активными перемещениями caмих рыб, не выходящих за пределы вод с благоприятными для них условиями. В то же время способность к продолжительному плаванию дает этим рыбам возможность дифференцированно использовать разные части ареала в продолжение жизненного цикла. В соответствии с этим в ареалах нектонных рыб различают 1) область размножения (нереста), или репродуктивную часть ареала; 2) нагульную, или вегетативную, часть ареала, используемую видом для откорма путем двусторонних активных (иногда — за счет разноса молоди — пассивных в начальной фазе) миграций, нередко имеющих сезонный характер; 3) область пассивного выселения планктонных икринок, личинок и мальков. Структура ареала нектобентических рыб в принципе та же, но их миграции не имеют значительной протяженности.
Атлас-определитель рыб: Книга для учащихся. Н.А.Мягков 1994 г.
КЛАСС КОСТНЫЕ РЫБЫ (OSTEICHTHYES)
Первичноводные позвоночные животные с окостеневшим или полностью костным скелетом. Жабры прикрыты костной жаберной крышкой. В водах нашей страны обитают представители одного подкласса — Лучеперые рыбы (Actinopterygii), на долю которого приходится более 99% ныне живущих видов (всего их по разным оценкам 22—23 тыс. видов).
Иллюстрированная энциклопедия рыб. Ст.Франк 1983 г.
О костистых рыбах мы узнаем, что они появились в среднем триасовом периоде. Во время юрского периода они значительно распространились и в конце мелового периода обитали как в морях, так и в пресных водах. До сего времени нет единого мнения об их происхождении. Некоторые авторы предполагают, что они возникли в море, а другие - в пресных водах. Многие виды меняли обе среды, как это делают в наше время некоторые мигрирующие рыбы (например, лосось, угорь). Хотя современные морские и пресноводные костистые рыбы представлены почти 20 тыс. видами, достаточных палеонтологических данных, характеризующих их образование и развитие в прошлых геологических периодах, в распоряжении ученых нет. Наиболее подробные материалы такого рода имеются в отношении отряда СЕЛЬДЕОБРАЗНЫХ (Clupeiformes), положивших начало филогенетическому развитию многих современных костистых рыб. Во время юрского периода жили представители родов Leptolepis и Thrissops, являющиеся праотцами всего отряда сельдеобразных. Скелет головы Leptolepis dubius характеризуется, согласно Бергу, явным сходством с современными сельдями.
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
Костные рыбы (класс Osteichthyes)
обладают более или менее окостеневшим
внутренним скелетом, особенно в области
черепа, челюстей, жаберных крышек; обычно
имеются кожные кости на голове, а у
некоторых и на теле. Тело у большинства
покрыто костными чешуями. С каждой
стороны тела имеется только одно
наружное жаберное отверстие, прикрытое
жаберной крышкой. Как правило, у
большинства есть плавательный пузырь (или
легкое).
Этот класс включает два подкласса —
лопастеперых и лучеперых рыб.
КЛАСС КОСТНЫЕ РЫБЫ (OSTEICHTHYES)
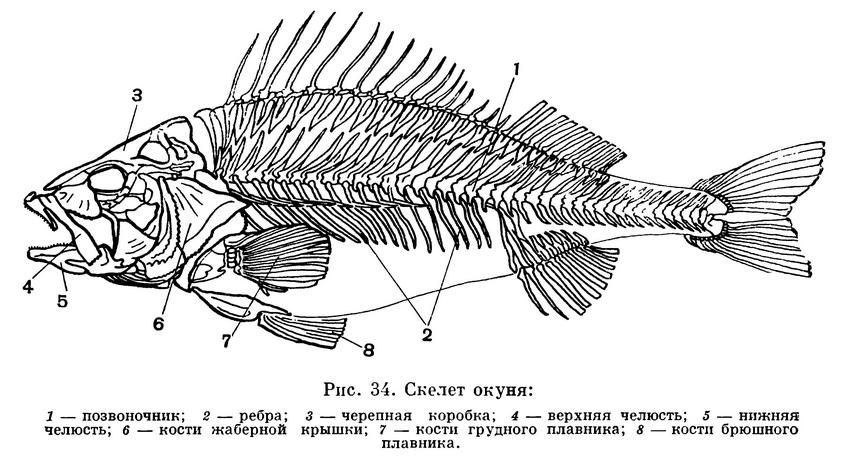
У костных рыб, как и у хрящевых,
имеются парные конечности — плавники, рот
образован хватающими челюстями с
зубами на них, жабры расположены на
имеющих внутреннюю скелетную опору
жаберных дугах, ноздри парные, во
внутреннем ухе имеются три полукружных
канала. В отличие от хрящевых рыб, в
скелете костных рыб имеется костная ткань,
в верхней части полости тела расположен
плавательный пузырь; жаберная полость
прикрыта укрепленной костным скелетом
жаберной крышкой; жабры имеют форму
свободно свисающих лепестков, а не
приросших к межжаберным перегородкам
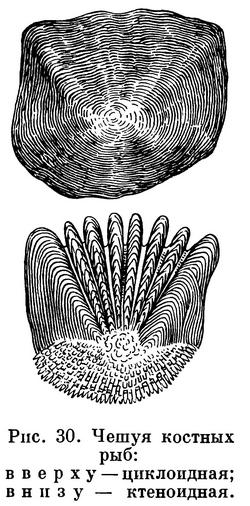
пластин; тело покрыто костной чешуей,
пластинками или голое вместо покрова
из зубовидной плакоидной чешуи.
Костные рыбы впервые появились на
нашей планете около 400 миллионов лет
назад. Наибольшего разнообразия они
достигли к началу нашей эры, далеко
обогнав всех других рыбообразных и рыб,
а также наземных позвоночных животных.
Это самая процветающая сейчас группа
позвоночных животных. Они
распространены во всех водах нашей планеты от
полюсов до экватора, в пресных водах,
солоноватых морях и океанах, от
горных озер и ручьев до
глубин океанских впадин, от
освещаемых солнцем коралловых
рифов до лишенных света пещер
и глубин океана. Исконные
обитатели водной среды, некоторые
рыбы приобрели способность
взлетать из воды, пролетая в
воздухе до нескольких сотен
метров, а у некоторых
развились приспособления в
жаберной полости или
плавательном пузыре, позволяющие им
долгое время быть вне воды в
атмосфере влажного воздуха
тропиков. Наконец, некоторые
рыбы — двоякодышащие,
многоперы — имеют наряду с
жабрами и ячеистый плавательный
пузырь или настоящие легкие,
позволяющие им дышать
атмосферным воздухом.
Среди костных рыб есть свои гиганты
и карлики — от достигающих 5—7 м
длины и 500—1500 кг веса пресноводных
белуги, калуги, сома, бразильской арапаимы и морских меч-рыбы и марлинов до
крошечных филиппинских бычков, 7—11 мм длины. Таким образом, костные
рыбы не достигают величины хрящевых,
среди которых имеются акулы до 15—18 м
длины, но зато среди них имеется много
гораздо более мелких.
Форма тела у большинства костных рыб
более или менее торпедообразная,
приближается у наиболее быстрых и
неутомимых пловцов, как например тунцы и
макрели, к идеально обтекаемым очертаниям.
Однако у многих рыб, ведущих иной образ
жизни, тело имеет другую форму.
Различимы разные типы строения. Стреловидный
тип, характеризующийся удлиненным
телом, приостренным рылом и
отодвинутыми назад, подобно оперению стрелы,
непарными плавниками, встречается обычно
у рыб, неподвижно стоящих в воде и
настигающих добычу или спасающихся от
врага внезапным броском — таковы,
например, щука, барракуда, сарган, сайра.
Змеевидная форма тела характерна для
угрей, передвигающихся, как
змеи, энергетически экономным
отталкиванием от окружающей
среды многими извивами тела.
Модификацией этого типа
является лентовидная форма,
способствующая, вероятно, как
увеличению эффективности
отталкивания, так и пассивному
сплыванию в течении.
Лентовидную форму тела имеют
мурены, цеполы, вогмеры,
некоторые глубоководные угри (авоцеттина), личинки угрей — лептоцефалы. Шаровидную форму
тела или форму неправильного
параллелепипеда имеют
прибрежные и сплывающие по
течению иглобрюхи, глубоководные
удильщики, крутящиеся среди
выростов коралловых рифов
закованные в костный
панцирь кузовки. Активное передвижение у этих рыо осуществляется
с помощью ундулирующих
(волнообразных) движений непарных плавников и
гребных движений грудных плавников.
Также с помощью ундулирующих
движений непарных плавников активно
перемещаются спинороги и луны-рыбы, имеющие
сжатое с боков дисковидное тело, а также
палочковидные морские иглы. Высокое,
сжатое с боков тело имеют держащиеся
вблизи крутых склонов, а также в пелагиали морские караси, морские лещи,
помпано. У луны-рыбы тело имеет форму
толстого диска. Наконец, у многих
донных рыб тело более или менее сплюснуто
сверху вниз, ширина тела больше его
высоты и глаза направлены вверх. Такую
форму имеет тело у бычков, керчаков,
плоскоголовое и других придонных рыб.
У собственно донных, лежащих на дне
камбал тело имеет форму плоского
ромба, эллипса или диска. У некоторых рыб
тело имеет совершенно своеобразную
причудливую форму, как например у
морского конька, напоминающего шахматную
фигуру коня, с головой, посаженной под
прямым углом к оси тела.
Тело рыбы внешне расчленяется на
голову, туловище и хвост, границами
которых принимают наружную жаберную щель
и, обычно, местоположение
заднепроходного (анального) отверстия. Для
регулирования положения тела служат
плавники: парные грудные и брюшные и
непарные спинной (их бывает от одного до
трех), хвостовой и анальный (бывает один
или два). Основной орган движения —
хвост. Однако у некоторых рыб, например
у морской иглы, луны-рыбы, спинорогов,
перемещение совершается
преимущественно с помощью волнообразных
(ундулирующих) движений непарных плавников.
На боках тела и на голове обычно
заметны поры боковой линии — органа
особого, свойственного только водным
животным, чувства восприятия движений воды.
Благодаря боковой линии даже
ослепленная рыба не натыкается на препятствия
и способна ловить движущуюся добычу.
Почти у всех рыб спина окрашена
темнее боков, а самая светлая часть тела —
брюхо. Такая окраска имеет защитное
значение, скрывая рыбу как при взгляде из
воздушной среды на кажущуюся сверху
темной поверхность воды, так и при взгляде снизу, навстречу свету небосвода.
У пресноводных рыб спина обычно бурая
или зеленоватая, соответственно
желтоватому оттенку пресной воды. У рыб
открытого моря, как например тунцов,
летучих рыб, океанической сельди, спина
темно-синяя или фиолетово-синяя. На
глубине от 100 до 200 м обычны серебристые
рыбы. Глубже, от 200 до 500 м, многие
рыбы красноватые или красного цвета.
Еще глубже их сменяют бурые,
фиолетово-черные и черные рыбы. Наконец, у
придонных рыб очень больших глубин
кожа нередко не окрашена вовсе. И также
вовсе не имеют окраски слепые рыбы
пещерных вод.
Напротив, окраска придонных рыб
малых глубин очень разнообразна — от
однотонно-серой или бурой до самых ярких
и необычайных сочетаний цветов и узоров.
Обычно, если имеются пятна и полосы на
теле рыбы, они делают ее менее заметной
на фоне камней и водорослей, среди
которых она живет. Такая защитная
окраска иногда замечательно точна: среди
гранитных скал встречаются рыбы с гранитоподобным узором, черные рыбы —
среди кусков лавы, оливково-желтые —
среди бурых водорослей и красные —
среди красных водорослей и кораллов.
Некоторые донные рыбы способны к
быстрому изменению узора и цвета
применительно к характеру окружающего
участка дна. Особенно замечательны в этом
отношении многие камбалы.
Помимо окраски под цвет окружающего
рыбу фона, существует и принципиально
иная окраска, характеризующаяся
резкими полосами, яркими пятнами на теле
и глазоподобными круглыми пятнами на
хвосте. Такая окраска искажает форму
тела рыбы, разбивая контур тела на части
и дезориентируя врага и добычу.
Окраска играет также существенную
роль для опознавания особей своего вида
и особей противоположного пола. У
многих рыб в период размножения окраска
специфически изменяется, стимулируя
изменения в поведении самца и самки,
ухаживание и нерест. Изменения окраски
служат также для отпугивания более
слабого соперника.
Совершенно необыкновенное богатство
и причудливое разнообразие узоров и
ярких сияющих красок всех цветов
радуги встречается у рыб, живущих среди
коралловых рифов — в условиях яркого
солнечного освещения и резких теней,
преимущественно у щетинозубых рыб,
рыб-бабочек, губанов, рифовых окуней,
мурен. Красочный, сияющий всеми
цветами мир этих рыб не уступает по своей
красоте миру бабочек и птиц.
У многих глубоководных рыб роль
окраски выполняют световые органы — фотофоры. Их расположение на теле
рыбы и цвет испускаемого ими света
специфичны для каждого вида и имеют также
характерные половые различия,
отличающие самца от самки.
Рот рыбы обычно вооружен зубами;
зубы бывают не только на челюстях, но
нередко и на нёбных костях, на сошнике,
на языке, на костях жаберного аппарата
(глоточные зубы). Бока глотки укреплены
пятью парами костных жаберных дуг, по
внутреннему краю которых расположены
жесткие жаберные тычинки, а по
внешнему — обильно снабженные кровью
лепестки жабр. Сквозь 4 пары щелей между
жаберными дугами рыба активно
пропускает воду, процеживая ее сквозь решетки
из жаберных тычинок и жаберных
лепестков. С помощью первых задерживаются
в глоточной полости и поступают в
пищевод рачки и другие организмы, служащие
рыбе пищей, тогда как в жаберных лепестках происходит окисление проходящей
крови путем извлечения из омывающей их
воды растворенного в ней кислорода.
Жабры служат органом дыхания рыбы.
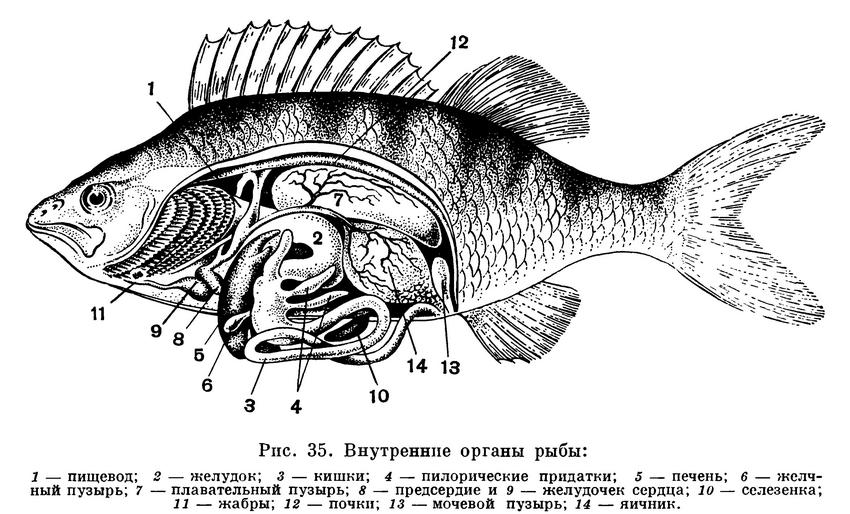
Кишечник обычно сравнительно
слабо дифференцирован на отделы:
специфичны для рыб слепые выросты — пилорические придатки (от 1 до
200), открывающиеся в начале средней
кишки, сейчас же за желудком; у
примитивных рыб, например осетровых, в
толстой кишке имеется спиральная
складка, как у акул и скатов.
К кишечнику прилегает лопастная печень, снабженная желчным пузырем.
Поджелудочная железа
обычно слабо обособлена: маленькие дольки
ее (островки) прилегают к желудку или
вкраплены в печень. Ею вырабатывается
инсулин, и возможно использование ее
в качестве сырья для получения этого
ценного лечебного препарата.
Сердце расположено в передней части
полости тела — в области,
соответствующей горлу рыбы. Оно состоит из
предсердия и желудочка, и через него проходит
только венозная кровь, нагнетаемая
сердцем в жабры. Оттуда же после
обогащения кислородом в жаберных лепестках
кровь поступает в различные органы тела.
У рыб двухкамерное сердце и только один
круг кровообращения. Только у
двоякодышащих рыб, в связи с наличием легких,
кровеносная система сложнее.
Почки у рыб имеют вид
темно-красных лент, располагающихся сейчас же под
позвоночником и простирающихся вдоль
всего спинного края полости тела. Между
ними и кишечником расположен
плавательный пузырь, несущий у рыб функции
гидростатического аппарата, а также
органа, регулирующего газообмен и, у
некоторых рыб,— функцию резонатора звука.
Мешковидные яичник и (или ястыки) у самок и лопастные беловатые семенники (или молоки) у самцов
имеют выводные каналы, открывающиеся
наружу на мочеполовом или особом
половом сосочке, позади заднепроходного
отверстия.
Мозг у рыб обычно очень невелик и
устроен весьма примитивно: кора
переднего мозга, служащая у высших
позвоночных ассоциативным центром, у
костных рыб вовсе неразвита в отличие от
акул, у которых она состоит из мозговой
ткани.
Центры различных чувств обособлены
в разных отделах мозга: обоняния — в
переднем мозге, зрения — в среднем, слуха
и осязания — в продолговатом, центр
координации движений — в мозжечке;
относительная величина этих отделов
соответствует роли различных чувств в жизни
рыбы, и внешний вид мозга позволяет
судить об образе жизни.
Особое значение имеет нижний мозговой
придаток — гипофиз, имеющий вид
маленькой луковички, сидящей на нижней
поверхности мозга, позади скрещения
зрительных нервов. Впрыскивание
созревающим самкам рыб экстракта гипофиза
чрезвычайно ускоряет дозревание икры
и применяется с этой целью в
промышленном рыбоводстве.
По образу жизни различают
пелагических рыб, держащихся в верхних слоях
и в открытом море (сельди, сардины,
сарганы, скумбрия, сайда, тунцы), и
придонных и донных рыб, обитающих на дне
или у дна (треска, пикша, камбаловые,
горбылевые, бычки и т. д.); стайных и
одиночных рыб. Особую группировку
составляют глубоководные рыбы, среди которых
различают батипелагических и
придонных.
По характеру питания различают
планктоноядных рыб, потребляющих
планктон (мелкие рачки и другие
организмы, «парящие» в толще воды), бентосоядных (бентос — организмы, живущие
на дне) и хищных рыб.
Вес пищи, потребляемой рыбой в
течение года (годовой рацион), составляет
обычно от 4—7 до 23—25 (у хищников)
собственных весов самой рыбы.
Многие рыбы зимой прекращают
питаться, некоторые виды собираются во
впадинах дна, погружаясь в зимнее
оцепенение. Таковы, например, многие
осетровые, карповые, сомы, во множестве
залегающие на «ямах» в дельте Волги,
камбалы, залегающие на зимовку в водах
Камчатки. Зимнее прекращение питания
обусловливает ежегодную зимнюю
приостановку роста, отпечатывающуюся на
костях рыб, в том числе на чешуе, в виде
уплотненных зон костного вещества —
«зимних колец».
Прекращают питаться рыбы обычно
также в период нереста (размножения),
нередко сильно истощаясь в этот период.
Полового созревания рыбы достигают
в возрасте от нескольких месяцев
(тюлька) до 6-11 (треска) и 16—20 (белуга)
лет. Созревшие рыбы обычно нерестуют
ежегодно, стареющие рыбы нерестуют не
каждый год и к концу жизни перестают
размножаться. Некоторые виды нерестуют
только один раз в жизни, после чего
погибают. Таковы дальневосточные лососи,
некоторые сельди, азовские и каспийские
бычки.
Плодовитость рыб колеблется от
нескольких десятков икринок (у некоторых
арктических рогаток, у бычков) до 1—9
миллионов икринок (у камбаловых и
тресковых) и даже 28 миллионов (у луны-рыбы). Но из этого огромного количества
икринок только единичные особи достигают
взрослого состояния. Икра либо тяжелее
воды (донная икра) и откладывается
рыбой на дно, на водные растения или
зарывается в грунт, либо ее удельный вес
примерно равен удельному весу воды, и она
плавает у поверхности или в толще воды
(пелагическая икра).
Большинство рыб откладывает икру при
строго определенных условиях среды,
специфических для каждого вида и гораздо
менее изменчивых, чем те, при которых
обычно живет рыба вне времени
размножения. Для откладки икры рыбы
собираются в места с определенной
температурой, соленостью, аэрацией, щелочностью,
грунтом или субстратом (для рыб с донной
икрой); все эти факторы могут иметь
существенное значение. Развивающиеся
икринки и личинки, особенно на ранних стадиях
развития, обычно гораздо чувствительнее
к факторам внешней среды, чем взрослые
рыбы, поэтому выметывание икры при
строго определенных значениях этих
факторов способствует их лучшему
выживанию. Стремление рыб собраться для
нереста в места и условия, наиболее
подходящие для развития икры, способствует
необходимой для размножения
концентрации производителей, а также обособлению
разных видов друг от друга.
Большинство рыб, выметывающих
пелагическую икру, ограничивает заботу о
потомстве подходом на определенные
нерестилища, места и условия среды,
благоприятные для развития выметываемой
икры.
Многие рыбы, однако, заботятся о своем
потомстве не только путем подыскивания
специальных условий для откладываемой
икры, но и оберегают ее от врагов на
всех этапах развития.
Живорождение, наблюдающееся у
некоторых костных рыб, обеспечивается
путем внутреннего оплодотворения и
развития икринок внутри яичника в отличие
от хрящевых рыб, у которых зародыши
развиваются в расширениях яйцеводов.
Развитие отложенной икры длится от
нескольких часов (тюлька, многие
аквариумные рыбки) до нескольких недель
(треска, камбала), нескольких месяцев
(лососи) и до года (агонус). Выходящие из
икры личинки обычно сначала
малоподвижны и имеют еще остаток желточного
мешка (личинки с желточным мешком или
предличинки), затем желток всасывается,
и личинка переходит к активному
питанию. Тело личинки претерпевает ряд
изменений, у некоторых групп (например,
камбаловых, угрей, сельдей) очень
значительных, порядка метаморфоза.
Окончательное приобретение личинкой облика,
близкого к облику взрослой рыбы,
происходит обычно с момента появления на
ее теле чешуи,— с этого времени личинка
становится мальком. Мальков, имеющих
возраст менее года, обычно называют
сеголетками. Живут рыбы от года (некоторые
бычки) до 50—70 лет (щуки, карпы,
палтусы); большей частью
продолжительность жизни не свыше 15—20 лет.
Личинки и мальки рыб имеют множество
врагов: их пожирают хищные личинки
насекомых, рачки, гидроиды и медузы,
черви-стрелки, лягушки, ужи, птицы и т. д.
Наибольшее количество личинок,
однако, пожирается рыбами.
Защитой от врагов многим рыбам служат не
только покровительственная окраска, но
и превращенные в острые шипы краевые
лучи плавников, шипы на жаберной
крышке и темени, скальпелевидные
отгибающиеся шипы на боках хвостового стебля у рыб-хирургов. У некоторых рыб боковая и
задняя поверхность шипов выстланы
ядовитой железистой тканью, и уколы таких
шипов могут вызывать сильную боль и
опухание, а иногда и более тяжелые
последствия. Такие ядовитые шипы имеются
у многих скорпеновых (морские ерши,
морские окуни), «змеек», или «драконов», рыб-звездочетов («морские коровы»), лировых
рыб, а в тропических морях также у бородавчаток, морских сомов, сигановых и
рыб-жаб. Особенно сильно развитые
ядовитые железы имеются у обитающих в
водах Южной Европы (включая Черное
море) «змеек», или «драконов», а в
тропических водах — у бородавчаток и рыб-жаб.
Сильные уколы шипов этих рыб
причиняют острую, жгущую боль,
усиливающуюся в течение получаса до почти
непереносимой, вызывающую рвоту,
затемнение и потерю сознания пострадавшего,
иногда сердечную слабость, конвульсии
и другие явления, вплоть до смертельного
исхода. Полное выздоровление от сильных
уколов змеек обычно требует нескольких
дней, затягиваясь иногда на несколько
месяцев, а выздоровление от уколов бородавчатки может растягиваться на много
месяцев, влияя на общее состояние здоровья
пострадавшего.
Рыбы иногда довольно сильно страдают
от бактериальных болезней и паразитов.
Некоторые болезни (например, краснуха
карпов и угрей, чума лососей) приводят
к массовой гибели рыб. Сильные
опустошения производят иногда и паразиты
(например, кровососущий червь-сосальщик
нитчия — у аральского шипа в 1936 г.).
Некоторые паразиты имеют своими
хозяевами пресноводную рыбу и человека,
передаваясь последнему при употреблении
в пищу недостаточно проваренной или
прожаренной рыбы; таковы в СССР широкий
и малый лентецы — в северо-западной
области и в Сибири, сосалыцик-метагонимус — на Амуре и кошачья
двуустка — в Сибири. Следует отметить еще
червя-ремнеца, или лигулу, имеющего хозяевами рыб и водоплавающую птицу, но
не передающегося человеку. У морских
рыб нет общих паразитов с человеком.
По характеру обитаемых вод различают
пресноводных, морских и проходных рыб.
Только немногие виды собственно морских
рыб, например речная камбала, мойва,
входят в низовья рек, и только немногие
пресноводные рыбы выходят в море.
Исключительное положение занимает наша трехиглая колюшка, одинаково хорошо
чувствующая себя как в пресной воде, так
и в море. Большие изменения солености
выдерживают в тропической области ханосы и илистые прыгуны. В солоноватой
воде лиманов и особенно внутренних морей
и озер, например в Азовском и Каспийском
морях, наряду с пресноводными и
собственно морскими рыбами живут и солоноватоводные рыбы, не входящие в пресную
воду и не встречающиеся в полносольных
морях. Таковы многие бычки, некоторые
атерины, а из окуневых рыб — морской
судак и азовская перкарина. В
тропической области в солоноватых водах вблизи
устьев рек держатся морские сомы, сребробрюшки, мохарры, некоторые сельдевые
и анчоусовые рыбы.
Проходными, или диадромными,
называют таких рыб, которые проводят часть
жизни в море, а часть в пресных водах,
переходя из одной среды в другую в
определенные периоды жизни.
Различия районов размножения,
откорма, роста, зимовок и летнего нагула
обусловливают существование закономерных
странствований по определенным путям
и в определенное время — миграций.
Миграции составляют непременную часть
жизненного цикла многих рыб.
Руководясь течениями, температурой и
соленостью воды, а также распределением
кормовых организмов, под влиянием
внутренних импульсов, многие рыбы совершают
сезонные, кормовые и нерестовые
миграции, активно проходя путь от пастбищ
и зимовок к местам размножения и
пассивно сплывая в виде икры и личинок от
мест размножения к местам откорма.
Большинство проходных рыб северного
полушария — лососи, большинство
осетровых, некоторые сельди, карповые и
окуневые — растут и кормятся в море, а
откладывают икру в пресной воде, в
которой только и возможно развитие икры
этих видов. Это анадромные проходные
рыбы. Громадные стаи тихоокеанских
лососей подходят в период размножения из
океана к устьям рек и поднимаются в их
верховья для нереста, совершая так
называемые анадромные миграции — вверх,
против течения. Проходные сельди,
осетры и белуги поднимаются для нереста в
реки Каспия, Азовского и Черного морей.
Вышедшие из икры анадромных проходных
рыб личинки пассивно сплывают вниз по
течению в море, где они растут и
откармливаются до созревания.
Имеются, однако, и проходные рыбы,
живущие и откармливающиеся в пресных
водоемах, а для нереста сплывающие вниз
по течению рек в море. Такие катадромные миграции совершают речные угри,
многие филиппинские бычки, некоторые
галаксиевые в водах Австралии и Новой
Зеландии.
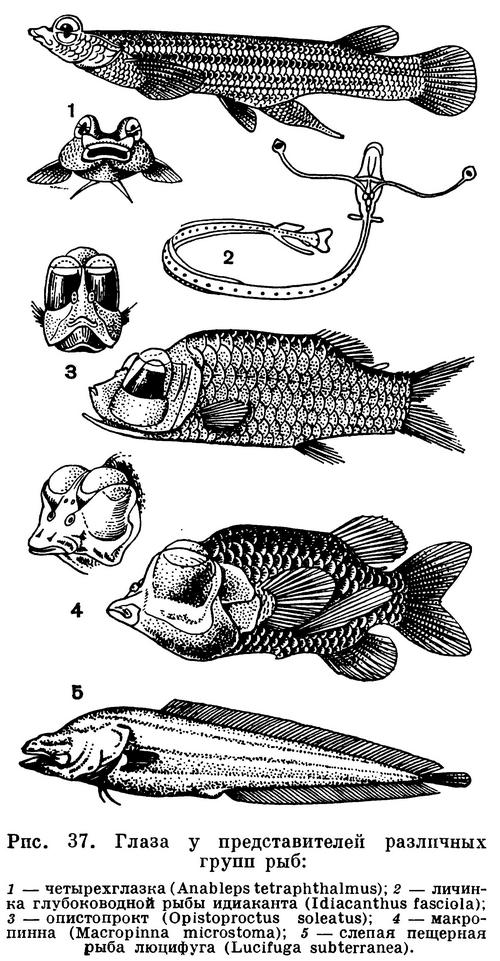
Жизнь и поведение рыб, как и всех
позвоночных, регулируется органами чувств.
У костных рыб развиты органы обоняния,
вкуса, зрения, слуха и равновесия, сейсмосенсорного чувства и чувства
электрического поля.
Орган обоняния у большинства костных
рыб имеет вид слепого мешка,
открывающегося наружу одной или двумя
ноздрями на конце рыла или впереди глаз.
Только у двоякодышащих рыб и немногих
придонных костных рыб обонятельный
мешок имеет и внутреннюю ноздрю — хоану, сообщающуюся и с ротовой
полостью. Органы вкуса или ощущения
химизма воды представлены группами
чувствующих клеток, расположенных в
коже усиков, губ, рта. Глаза рыб обычно
хорошо развиты, хотя у многих пещерных
и у некоторых глубоководных рыб глаза
вовсе атрофированы. Однако у некоторых
глубоководных рыб глаза бывают очень
велики и имеют цилиндрическую, а не
обычную сферическую форму; это так
называемые телескопические глаза. Органы
слуха и равновесия у рыб представлены
внутренним ухом, наружное ухо у них
отсутствует. У многих рыб имеется связь
внутреннего уха с плавательным пузырем
посредством цепи специальных косточек
(веберов аппарат карповых, вьюновых и
сомовых рыб) или с помощью доходящих до
слуховой капсулы идущих вперед
отростков плавательного пузыря (сельди,
анчоусы, треска, многие морские караси,
каменные окуни).
Непосредственно примыкают к органам
слуха по происхождению и функции
органы боковой линии, или сейсмосенсорного
чувства. Система боковой линии развита
у всех рыб — костных и хрящевых, а
также у водных амфибий. Она состоит из
рядов чувствующих кожных органов (плакод или купул), обычно помещающихся
в особых каналах, сообщающихся порами
с внешней средой или непосредственно
выступающих на поверхности тела. Это
специальные органы водных животных,
позволяющие улавливать
распространяющиеся в воде колебания, вплоть до
малейших сотрясений воды, возникающих
от активно плывущих животных.
Возможно, что эти органы также позволяют рыбам
ощущать изменения давления,
предвещающие разрядку в виде подводных
землетрясений.
Наконец, только у некоторых рыб среди
всех животных известны специальные
органы, генерирующие электрический ток
и разряды. Электрические органы этих
рыб состоят обычно из преобразованных
частей мышц и построены по принципу
вольтова столба. У некоторых рыб они
генерируют сильные, оглушающие добычу
или отпугивающие врага разряды
постоянного тока напряжением до 100, 200 и
даже 650 вольт и силой около 2 ампер (т. е.
мощностью свыше 1000 ватт). Таковы
американские электрические угри — гимноты (Electrophorus) и африканские электрические сомы (Malapterurus) — из
костных рыб, а также живущие в теплых морях
электрические скаты, или гнюсы
(Torpedo) — из хрящевых рыб. Другая группа
электрических рыб — африканские
клюворылы и гимнархи (Mormyridae), а
также некоторые виды гимнотов (кроме
видов рода Electrophorus) испускают серию
низковольтных импульсов, до 200 и даже
до 1600 импульсов в секунду, создавая
вокруг себя электрическое поле.
Искажение этого поля при попадании в него
возможной добычи или других объектов
позволяет рыбам уверенно находить себе
пищу в мутной воде африканских рек, а
также обнаруживать особей
противоположного пола. На коже таких рыб имеются
специальные чувствительные органы
восприятия электрического заряда —
электрорецепторы, устроенные наподобие
органов боковой линии и помещающиеся в
особых каналах.
Взаимосвязи в поведении рыб — «язык»
рыб — осуществляются в значительной
мере зрительными и слуховыми
восприятиями. Рыбы сигнализируют друг
другу о наличии пищи, об опасности, угрозе,
готовности к нересту. Такие сигналы
подаются позами и движениями,
соответствующей окраской и световыми вспышками
(у глубоководных рыб), выделением
химических веществ, созданием
электрических полей, звуками. У многих рыб
слегка наклоненное головой вниз, немного
изогнутое тело с несколько прижатыми
плавниками сигнализирует обнаружение
корма; другие рыбы, увидевшие в таком
положении собрата, дружно несутся к
этому месту и начинают разыскивать пищу.
Поза готовности к нападению («поза
агрессии») характеризуется напряженно
раздвинутыми плавниками,
оттопыренными жаберными крышками, открытым
ртом, волнообразными движениями
спинного плавника, резким изменением
окраски. При этом тело угрожающей
нападением рыбы кажется крупнее, а
контрастные цветные пятна, нередко имеющиеся
на жаберных крышках и спинном
плавнике, резко выделяются. Очевидно, такая
поза рассчитана на запугивание и
деморализацию противника. Разнообразные
позы принимаются рыбами во время их
«нерестовых игр», которыми самцы и
самки выражают друг другу степень
готовности к нересту, приглашают к постройке
гнезда и привлекают в гнездо (у строящих
гнезда рыб) и т. д. У стайных рыб хорошо
выражены сигналы, координирующие
движения: движущаяся вперед рыба обычно
вызывает у других стайных рыб так
называемый рефлекс следования.
Многие рыбы издают звуки; прежнее
представление о подводной среде как
о мире тишины совершенно неправильно.
Мы не слышим рыб только потому, что
только ничтожная часть энергии
изданного в воде звука переходит сквозь
поверхность воды в воздух, и потому, что
многие издаваемые рыбами звуки
распространяются на волнах, плохо слышимых
нами в воде без специальных приборов—
гидрофонов. Рыбы издают звуки в
диапазоне частот от 20—50 гц до 10—12 кгц при
пределах слышимости человеческого уха
от 16 гц до 20 кгц.
Звуки одних видов рыб напоминают
потрескивание или шелест, скрежет,
рычание и ворчание, других — цоканье,
одиночные удары и барабанный бой, третьих —
глухие стоны, карканье, хриплые крики,
хрюканье, пароходные гудки, вой
сирены, четвертых — птичий щебет,
кудахтанье, высокие трели. Сила рыбьих
звуков колеблется от еле слышимых до таких,
которые могут взрывать акустические
мины — от десятых долей бара до 100—
200 бар. Рыбы издают звуки с помощью
разных приспособлений и аппаратов: у
одних — это трение костей друг о друга —
глоточных или челюстных зубов («скрежетанье» зубами), зазубренных шипов или
сочленений жаберных крышек; у других —
это особые резонансные устройства
плавательных пузырей, издающие звуки при
резком сокращении пузырей или ударах
по ним особых звуковых косточек.
Особенно развиты звуковые аппараты у горбылевых рыб, тригл, или морских
петухов, скорпеновых рыб, рыб-жаб и
некоторых других. Слышимые из-под воды
звуки отдаленного барабанного боя,
испускаемые некоторыми американскими
горбылевыми рыбами-барабанщиками, не
раз вводили в заблуждение незнакомый
с ними европейский персонал кораблей,
пришедших в воды Америки. Звуки,
издаваемые черноморским
губаном-зеленушкой, слышны из аквария как глухой удар,
а звуки морского конька напоминают звук
лопнувшего стакана.
Разными звуками рыбы извещают о
наличии пищи, о появлении врага, о
занятости участка дна и охране его, о поиске
самки («нерестовые песни»).
Наряду с внутривидовыми
взаимосвязями огромное значение имеют
взаимоотношения разных видов рыб, а также рыб
и других видов животных.
Повседневны взаимоотношения
трофические и конкурентные: хищника и
добычи, потребителей сходной пищи, видов,
подыскивающих сходные условия для
нереста, и т. д.
Очень интересны взаимоотношения
особого типа, при которых партнеры
взаимосвязаны образом жизни с пользой для
обоих участников (симбиоз) или с пользой
для одного без вреда для другого
(комменсализм). Такие отношения связывают
некоторые виды рыб с беспозвоночными —
кишечнополостными, моллюсками,
ракообразными, иглокожими; с оболочниками
и высшими позвоночными; с бактериями,
а также разные виды рыб друг с другом.
Мальки некоторых тресковых,
ставридовых и нередко строматеевых рыб
держатся под колоколом крупных медуз,
странствуя под их защитой, а мальки
кубохвостов или алетов нередко
встречаются в полостях сальп и колоний пиросом
(огнетелок).
Небольшой номей (Nomeus albula) всю
жизнь проводит под защитой жгучих
щупалец сифонофоры-физалии,
распространяясь вместе с ней на огромных
пространствах тропической области Мирового
океана. Ярко окрашенная рыбка амфиприон
живет вместе с большими актиниями,
безнаказанно суетясь в чаще усаженных
ядовитыми стрекательными клетками
щупалец и прячась от врагов в полости
актинии. Наконец, некоторые скорпеновые,
ядовитая бородавчатка, некоторые агоновые рыбы несут на себе целые заросли
гидроидов, помогающих им маскироваться
на дне.
Под надежной защитой длинных игл
морских ежей диадем (между ними)
держатся головой вниз, вертикально, мелкие
ножебрюхие рыбки — кривохвостки или
эолиски. Замечателен симбиоз с
крабом каллианассой слепого бычка тифлогобиуса, который живет вместе с
крабом в отрытых хозяином норах и питается
остатками приносимой им пищи.
Но некоторые креветки сами обслуживают
крупных рыб коралловых рифов — мурен,
губанов, рифовых окуней: они
безбоязненно приближаются к этим рыбам, выедая
с их тела мелких рачков-паразитов. Это
креветки-чистильщики, и очищаемые рыбы подставляют им себя для очистки, не
покушаясь на своих «санитаров».
Такую же роль санитаров, выедающих
рачков-эктопаразитов с жабр и кожи
крупных рыб коралловых рифов, играют
и некоторые мелкие виды губанов и
бычков. Тело этих рыбок украшено одной
или двумя яркими темно-синими или
черными полосами на боках, и крупные
рифовые окуни отличают их от других рыб,
приоткрывают им жабры, раскрывают рот,
обеспечивая наилучший поиск и выедание
паразитов.
Широко известно содружество рыбы-лоцмана с акулами, которых она обычно
сопровождает, а также использование
рыбами-прилипалами акул и гигантских
скатов-рогачей, марлинов, морских черепах,
дельфинов и китов в качестве удобных
средств транспорта.
Совершенно замечателен симбиоз
некоторых рыб со светящимися бактериями:
светящиеся бактерии обитают в
специальных кожных карманах или каналах под
кожей, часто подостланных с внутренней
стороны отражающей свет тканью, а
снаружи прикрытых линзообразно вздутыми
прозрачными участками кожи или
снабженных регулирующими силу света
прикрывающими устройствами. Такой
симбиоз наблюдается у моровых и
долгохвостое (из трескообразных рыб), у сребробрюшек, моноцентровых
(рыбы — еловые шишки), фонареглазых (Anomalopidae), акропомовых рыб, а также у мешкоротов, удильщиков и некоторых других
рыб. Имеются даже специальные виды
светящихся бактерий, живущие только на
рыбах-хозяевах, которым они несомненно
полезны.
В качестве одностороннего развития
явлений симбиоза и комменсализма
может рассматриваться паразитизм —
явление, при котором паразитирующий вид
(обычно имеющий малые размеры) живет
на теле или внутри тела хозяина,
используя последнего и настолько завися от его
жизнедеятельности, что не может
существовать самостоятельно. Многие
организмы паразитируют на рыбах —
грибковые, бактерии, черви, ракообразные,
личинки моллюсков, а из позвоночных
круглоротые — миноги и миксины.
Существует и несколько видов рыб,
паразитирующих на более крупных рыбах.
Маленький южноамериканский сомик кандиру, или ванделлия,
имеющий угреобразное тело, обычно живет на жабрах более
крупных рыб, раня их своими
крышечными шипами и высасывая текущую кровь.
Глубоководный тупорылый угорь — сименхел — наподобие миксины вгрызается
в тело крупных рыб, питаясь их мышцами.
Своеобразны отношения самцов и самок
некоторых глубоководных рыб —
удильщиков: мелкие самцы, размером всего
несколько сантиметров, прикрепляются
и прирастают к телу гораздо более
крупных самок, причем их кровеносная
система становится придатком кровеносной
системы самки. В этом последнем случае
в отличие от настоящего паразитизма
неспособные к самостоятельному
существованию самцы нужны самке для
воспроизведения вида.
Многообразная и сложная экология рыб,
необыкновенное разнообразие их
поведения, образа жизни, путей общения и
взаимосвязей друг с другом и с животными
других классов и типов подводного мира
только слабо намечены в кратком общем
очерке и сообщаются ниже при описании
соответствующих групп и видов этого
замечательного класса позвоночных
животных.
Костные рыбы имеют очень большое
практическое значение для человека.
Миллионы людей заняты ловом, разведением
и обработкой рыбы, постройкой судов и
изготовлением рыболовного снаряжения.
В некоторых странах население питается
главным образом рыбой, и благосостояние
его зависит преимущественно от величины
улова.
Ужением рыбы и подводной охотой
увлекаются сотни тысяч людей, которым этот
замечательный спорт дает здоровье и
отдых. Еще больше число аквариумистов,
создающих красочный тихий мир в
стеклянных сосудах своих аквариумов.
Помимо продуктов питания, рыбы
служат сырьем для получения лекарства
(рыбий жир), корма для скота и птицы
(кормовая мука), удобрения для полей
(туки), технического жира, клея, кожи
и других продуктов, используемых в
пищевой и легкой промышленности.
Система костных рыб (класс Osteichthyes) все еще недостаточно разработана.
Относительно простые схемы, использовавшиеся еще недавно, неприемлемы после
обстоятельных морфолого-таксономических и палеонтологических исследований
последних десятилетий. Эти исследования
показали необходимость существенных
изменений в ранее принятых
представлениях. Основной толчок для разработки
новой системы рыб был дан появлением
в 1940 г. классического труда советского
ихтиолога Л. С. Берга, за которым
последовал поток многочисленных работ
японских и американских ученых. Работа над
новой, естественной системой рыб еще
далеко не закончена. Система, предлагаемая
в этой книге, отражает основные достижения новых исследований, однако
несколько упрощена и эклектична в целях
удобства рассмотрения огромного
разнообразия форм класса костных рыб.
Костные рыбы отличаются от хрящевых
рыб развитием внутреннего костного
скелета, наличием костей на голове
(особенно в области жаберных крышек и
челюстей), развитием покрова из костных
чешуи незубовидного типа, наличием
плавательного пузыря (или легкого) и другими
признаками.
Этот класс состоит из двух подклассов —
подкласса лопастеперых (Sarcopterygii)
и подкласса лучеперых (Actinopterygii).