(Cretoxyrhinidae Glückman, 1958) Семейство Кретоксириниды, Кретоксириновые, Остроносые акулы мелового периода, Family Cretoxyrhinidae Glückman, 1958 (Cretaceous sharp-nose sharks) 7 родов
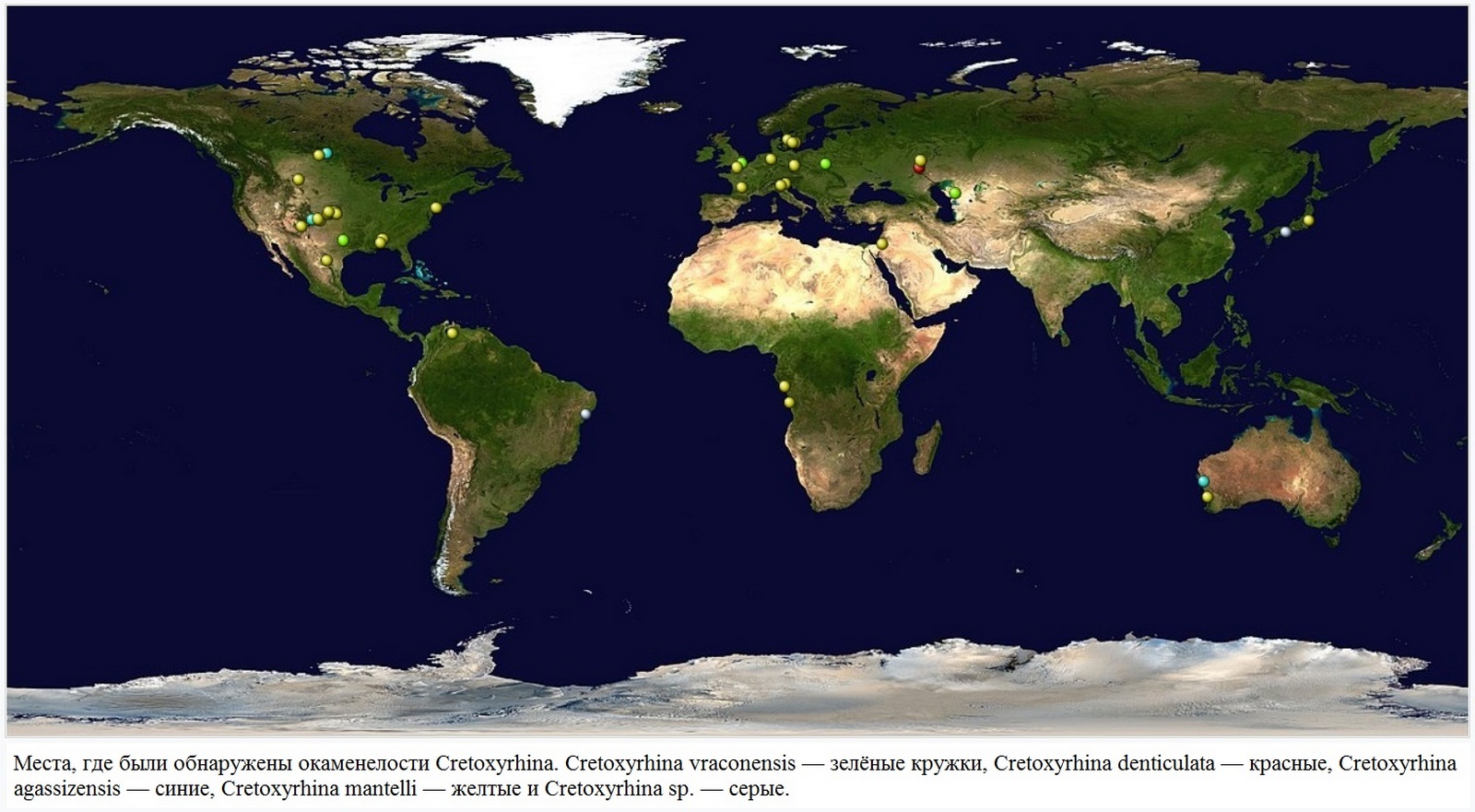
• Семейство † Cretoxyrhinidae Glückman, 1958 (Cretaceous sharp-nose sharks) — Остроносые акулы мелового периода, Кретоксириновые, Кретоксириниды — 130,0—13,65 млн лет назад, меловой, палеогеновый, неогеновый периоды. Остатки этих акул обнаружены преимущественно в Северной Америке (на территории США и Канады, частично в Мексике), Бразилии, встречаются в Европе (территория южной Франции, Испании, Германии, Великобритании, Дании, Украины, России), в Северной Африке, на Мадагаскаре, в Палестине, Сирии, Ливане, Иордании, Узбекистане, Пакистане, Индии, Австралии.
Название семейство получило от основного времени своего существования — мелового периода.
Кретоксириниды придержались вод от отмелей до средних глубин. Охотились на морских животных: костистую рыбу и более мелких акул. Следы от зубов крупных кретоксирин находили на костях мелких морских рептилий — мозазавров (семейство Mosasauridae) и плезиозавров (отряд Plesiosauria), что скорей всего поедались в основном в виде падали. По мнению исследователей, это были яйцеживородящие акулы. Хотя кретоксириниды эволюционировали конвергентно с большими белыми акулами (Carcharodon carcharias) и отодонтидами (Otodontidae), они не являются их предками, а представляют собой группу, которая полностью вымерла. Общая длина представителей семейства колебалась от 3 до 7 м. Голова большая, массивная. Морда была короткая и тупая. Глаза большие. Рот очень большой, широкий. Зубы мощные, длиной от 3 до 6 см. Верхняя челюсть выдаётся вперёд нижней. Туловище было крепким, широким. Эти акулы имели длинные грудные плавники, два спинных плавника, из которых вероятно передний был значительно больше заднего. Хвостовой стебель в основном был коротким, а хвостовой плавник большим и хорошо развитым.
• Род † Cretoxyrhina Glikman, 1958 (Cretaceous sharp-nose sharks — Остроносые акулы мелового периода, Кретоксирины) — 94,3—66,043 млн лет назад, поздний меловой период, Бразилия, Канада (Саскачеван), Конго-Браззавиль, Франция, Германия, Иордания, Казахстан, Российская Федерация, Швеция, США (Алабама, Колорадо, Канзас, Нью-Джерси). Зубы кретоксирин известны и из мела бывшего СССР: в Поволжье, Казахстане (Мангышлак). Род выделен советским палеонтологом Л. С. Гликманом в 1958 году. [Glikman, L.S. (1957) (Taxonomic significance of the accessory teeth of sharks of the families Lamnidae and Scapanorhynchidae). «in russian». Trudy geol. Muz. «A.P. Karpinskogo» Akad. Nauk S.S.S.R., 1: 103–109, 2 fig.]
Кретоксирин первым описал английский палеонтолог Гидеон Мантелл (Gideon Mantell) на основании восьми зубов Cretoxyrhina mantelli, которые он собирал на геологическом участке Southerham Grey Pit к юго-востоку от Льюиса (Lewes), Восточный Сассекс (East Sussex). В своей книге 1822 года «Окаменелости Саут-Даунс» («The fossils of the South Downs») он определил их как зубы, принадлежащие двум видам местных современных акул. Мантелл определил, что меньшие зубы принадлежат обыкновенной куньей акуле (Mustelus mustelus), а большие зубы — гладкой акуле-молот (Sphyrna zygaena), выразив некоторое сомнение по последнему определению. В 1843 году в Швейцарии натуралист Луи Агассис опубликовал третий том своей книги «Исследования ископаемых рыб» («Recherches sur les poissons fossiles»), в которой повторно исследовал восемь зубов Мантелла. Используя их и еще один зуб из коллекции Страсбургского музея — Strasbourg Museum (точное местонахождение находки зуба не указано, но он также был привезён из Англии), Агассис пришёл к выводу, что окаменелости на самом деле принадлежали одному виду вымерших акул, которые имели большое стоматологическое сходство с тремя видами (Oxyrhina hastalis, Oxyrhina xiphodon и Oxyrhina desorii), классифицированными в то время в ныне недействительном роде Oxyrhina (валидный род — Heterodon Latreille, 1801). Агассис отнёс этот вид к роду Oxyrhina, но отметил, что гораздо более толстый корень его зубов имеет достаточно большое значение, чтобы быть с научной точки зрения отдельным видом, и назвал таксон Oxyrhina mantellii, в честь Гидеона Мантелла (см. рисунок).
В конце 19 века палеонтологи описали множество видов, которые теперь являются синонимами Cretoxyrhina mantelli. По некоторым данным, в то время могло быть около 30 различных синонимов Oxyrhina mantelli. Большинство этих видов произошли от зубов, которые представляли разновидности Cretoxyrhina mantelli, но отклонялись от точных характеристик синтипов. Например, в 1870 году французский палеонтолог Анри Соваж (Henri Sauvage) идентифицировал зубы из Франции, которые очень напоминали синтипы Oxyrhina mantelli из Англии. Зубы также имели боковые выступы (небольшие эмалированные бугорки, которые находятся у основания основной коронки зуба), которые не присутствуют в синтипах, что побудило его описать зубы под видовым названием Otodus oxyrinoides на основании боковых выступов. В 1873 году американский палеонтолог Джозеф Лейди (Joseph Leidy) идентифицировал зубы из Канзаса и Миссисипи и описал их под видовым названием Oxyrhina extensiona. Эти зубы были шире и более крупные, чем у синтипов Oxyrhina mantelli из Англии.
Все изменилось с открытием некоторых исключительно хорошо сохранившихся скелетов акул в формации Ниобрара (Niobrara Formation) в Западном Канзасе, США. Чарльз Штернберг (Charles H. Sternberg) обнаружил первый скелет в 1890 году, который он описал в статье 1907 года: «Примечательным в этом образце является то, что позвоночный столб, хотя и был из хрящевого материала, был почти полным, и что было на месте большое количество зубов (250 зубов). Когда Час Истман (Chas. R. Eastman) из Гарварда описал этот образец, он оказался настолько полным, что уничтожил около тридцати синонимов, используемых для обозначения этого животного и происходящих от многих зубов, найденных в прежние времена.» [Sternberg, Ch. H., 1907, Some animals discovered in the fossil beds of Kansas.]
Чарльз Истман (Charles R. Eastman) опубликовал свой анализ скелета в 1894 году. В этой статье он реконструировал зубной ряд на основе разрозненного набора зубов скелета. Используя реконструкцию, Истман идентифицировал множество вымерших видов акул и обнаружил, что их окаменелости на самом деле представляют собой разные типы зубов Oxyrhina mantelli, которые он все отнёс к этому виду. Этот скелет, который Штернберг (Sternberg) продал Мюнхенскому университету Людвига-Максимилиана (Ludwig Maximilian University of Munich), был разрушен в 1944 году в результате бомбардировки союзников во время Второй мировой войны. В 1891 году сын Штернберга Джордж Штернберг (George F. Sternberg) обнаружил второй скелет Oxyrhina mantelli, который сейчас находится в Музей естественной истории Канзасского университета (University of Kansas Museum of Natural History) под названием KUVP 247 (см. рисунок, второй с верху). Сообщается, что этот скелет имеет длину 6,1 метра и состоит из частичного позвоночного столба с остатками скелета Xiphactinus в виде содержимого желудка и части челюстей с видимыми примерно 150 зубами. Этот скелет считался одним из величайших научных открытий того года из-за неожиданной сохранности хрящей. Джордж Штернберг на протяжении своей карьеры обнаружил ещё несколько скелетов Oxyrhina mantelli. Его наиболее заметными находками были FHSM VP-323 и FHSM VP-2187 (см. рисунок, третий с верху), найденные в 1950 и 1965 годах соответственно. Первый представляет собой частичный скелет, состоящий из хорошо сохранившегося набора челюстей, пары пяти жабр и нескольких позвонков, в то время как последний представляет собой почти полный скелет с почти полным позвоночным столбом и исключительно сохранившимся черепом, содержащим большую часть черепных элементов, челюсти, зубы, набор чешуек и фрагменты грудных поясов и плавников в их естественном положении. Оба скелета в настоящее время хранятся в Музее естественной истории Штернберга (Sternberg Museum of Natural History). В 1968 году коллекционер по имени Тим Басгалл (Tim Basgall) обнаружил ещё один примечательный скелет, который, как и FHSM VP-2187, также состоял из почти полного позвоночного столба и частично сохранившегося черепа. Эта окаменелость находится в Музее естественной истории Канзасского университета под названием KUVP 69102.
В 1958 году советский палеонтолог Леонид Гликман (Leonid Glickman) обнаружил, что дизайн зубов Oxyrhina mantelli, реконструированный Истманом (Eastman), дал достаточные основания, чтобы обосновать появление нового рода — Cretoxyrhina. Он также идентифицировал второй вид рода Cretoxyrhina на основе некоторых из более ранних зубов кретоксирин, которые он назвал Cretoxyrhina denticulata. Первоначально Гликман обозначил Cretoxyrhina mantelli как типовой вид, но он внезапно заменил это положение другим таксоном, обозначенным как «Isurus denticulatus» без объяснения причин в статье 1964 года. Этот шаг теперь отклонен как недействительная таксономическая поправка. Это, тем не менее, привело к тому, что российский палеонтолог Виктор Железко (Viktor Zhelezko) ошибочно объявил род Cretoxyrhina недействительнымв статье 2000 года, отождествив Isurus denticulatus (и, следовательно, род Cretoxyrhina в целом) с другим таксоном, идентифицированным как Pseudoisurus tomosus. Железко также описал новый вид, родственный Cretoxyrhina mantelli, на основе материала зубов из Казахстана, который он определил как Pseudoisurus vraconensis в соответствии с его таксономической переоценкой. Исследование, проведенное в 2013 году куратором и палеонтологом музея Западной Австралии (Western Australian Museum) Микаэлем Сиверсоном (Mikael Siverson), исправило таксономическую ошибку, восстановив род Cretoxyrhina и поместив вид Pseudoisurus vraconensis в него. В 2010 году британские и канадские палеонтологи Чарли Андервуд (Charlie Underwood) и Стивен Кумба (Stephen Cumbaa) описали Telodontaspis agassizensis из зубов, найденных в озере Агассис (Lake Agassiz) в Манитобе, Канада, которые ранее были идентифицированы как молодые зубы акулы из рода Cretoxyrhina. Этот вид был вновь включен в род Cretoxyrhina в результате исследования, проведенного в 2013 году американским палеонтологом Майклом Ньюбри (Michael Newbrey) с использованием дополнительных ископаемых материалов того же вида, найденного в Западной Австралии.
Между 1997 и 2008 годами палеонтолог Кеншу Шимада (Kenshu Shimada) опубликовал серию статей, в которых он проанализировал скелеты Cretoxyrhina mantelli, в том числе найденные Штернбергами, с использованием модернизированных методов для всестороннего исследования возможной биологии Cretoxyrhina. Некоторые из его работ включают разработку более точных стоматологических, морфологических, физиологических и палеоэкологических реконструкций, онтогенетических и филогенетических исследований на основе морфологических переменных. Исследования Шимады (Shimada) по кретоксирине помогли пролить свет на понимание акул и, с помощью новых методов, и других вымерших животных.
Название рода Cretoxyrhina образовано от слова creto (сокращение от Cretaceous — Меловой период) и от имени рода Oxyrhina, название которого происходит от древнегреческих слов οξυς (oxus) — «острый» и ρινα (rhina) — «нос». В совокупности они означают «остроносая акула мелового периода», хотя считается, что у Cretoxyrhina была довольно тупая морда.
Cretoxyrhina имела сходство с современной большой белой акулой (Carcharodon carcharias) по размеру, форме и экологии, но эти две акулы не являются близкими родственниками, и их сходство является результатом конвергентной эволюции. Cretoxyrhina традиционно относят к семейству ламноидных акул Cretoxyrhinidae, которое традиционно включает другие роды, приводящие к парафилетическому или полифилетическому семейству. Сиверсон (Siverson, 1999) заметил, что Cretoxyrhinidae использовался в качестве «таксона мусорной корзины» для акул мелового и палеогенового периода, и заявил, что род Cretoxyrhina был единственным действительным членом этого семейства.
Cretoxyrhina содержит четыре действительных вида: Cretoxyrhina agassizensis, Cretoxyrhina denticulata, Cretoxyrhina mantelli и Cretoxyrhina vraconensis. Эти виды представляют собой хроновиды. Самые ранние окаменелости Cretoxyrhina датируются 107 миллионами лет и принадлежат Cretoxyrhina vraconensis. Род будет развиваться благодаря Cretoxyrhina vraconensis, эволюционирующей в Cretoxyrhina denticulata в течение раннего сеномана (100,5—93,9 млн лет назад — Сеноманский ярус), которая в середине сеномана превратилась в Cretoxyrhina agassizensis, которая, в свою очередь, превратилась в Cretoxyrhina mantelli в позднем сеномане. Примечательно, что Cretoxyrhina agassizensis существовала до среднего турона (93,9—89,8 млн лет назад — Туронский ярус) и на короткое время была современником Cretoxyrhina mantelli. Это прогрессирование характеризовалось уменьшением латеральных зубцов, увеличением размера и прочности зубов. В интервале поздний альб (113,0—100,5 млн лет назад — Альбский ярус) — средний турон (93,9—89,8 млн лет назад — Туронский ярус) наблюдается в основном сокращение латеральных выступов на зубах. Cretoxyrhina vraconensis имел боковые выступы на всех зубах, за исключением нескольких в переднем положении, которые постепенно ограничивались только задними латерозадними зубами у взрослых к концу временного интервала у Cretoxyrhina mantelli. Поздний сеноман (100,5—93,9 млн лет назад — Сеноманский ярус) — коньяк (89,8—86,3 млн лет назад — Коньякский ярус) — этот временной интервал характеризовался быстрым увеличением размера зубов (и тела), значительным уменьшением отношения высота-коронка / ширина коронка и переходом от формы зубов разрывающего типа к форме зубов режущего типа. Размер зубов у особей Cretoxyrhina mantelli в пределах Западного внутреннего моря достиг своего пика около 86 миллиона лет назад во время последнего коньякского периода, а затем начинает медленно уменьшаться. В Европе этот пик приходится на более ранний период позднего турона. Самая молодая окаменелость Cretoxyrhina mantelli была обнаружена в формации Медвежья Лапа (Bearpaw Formation) в провинции Альберта, Канада, возраст которой составляет 73,2 миллиона лет. Единственный зуб, идентифицированный как Cretoxyrhina sp. был извлечен из близлежащей формации Подковообразного каньона (Horseshoe Canyon Formation) и датирован 70,44 миллиона лет, что позволяет предположить, что кретоксирины, возможно, дожила до маастрихта (72,1—66,0 млн лет назад — Маастрихтский ярус). Однако формация каньона Подкова имеет только солоноватоводные отложения, несмотря на то, что Cretoxyrhina была морской акулой, что делает вероятным то, что окаменелость была переработанной из более старого слоя. Переработанные окаменелости создаются путем эрозионной эксгумации (высвобождения) окаменелостей из горной породы, в которой они были первоначально отложены, и их повторного отложения в более молодых осадочных отложениях.
Филогенетические исследования на основе морфологических данных, проведенные Шимадой (Shimada) в 2005 году, показали, что Cretoxyrhina может быть родственником современных лисьих акул (семейство Alopiidae). В исследовании также говорится, что результаты являются преждевременными и могут быть неточными, и рекомендовано, чтобы Cretoxyrhina оставалась в семействе Cretoxyrhinidae, в основном ссылаясь на отсутствие для этого во время анализа существенных данных. [Kenshu Shimada (2005). «Phylogeny of lamniform sharks (Chondrichthyes: Elasmobranchii) and the contribution of dental characters to lamniform systematics». Paleontological Research. 9: 55–72.]
Другая возможная модель эволюции рода Cretoxyrhina, предложенная в 2014 году палеонтологом Каюсом Дидрихом (Cajus Diedrich), предполагает, что Cretoxyrhina mantelli была родственна акулам мако из рода Isurus и была частью расширенной линии Изурус, начиная с аптской (125,0—112,0 млн лет назад — Аптский ярус) стадии в раннем меловом периоде. Согласно этой модели, линия Isurus была инициирована видом Isurus appendiculatus (Cretolamna appendiculata), который в середине сеномана (100,5—93,9 млн лет назад — Сеноманский ярус) превратился в Isurus denticulatus (Cretoxyrhina denticulata), затем в Isurus mantelli (Cretoxyrhina mantelli) в начале коньяка (89,8—86,3 млн лет назад — Коньякский ярус), затем Isurus schoutedenti в палеоцене, затем в Isurus praecursor, где продолжается остальная часть линии рода Isurus. В исследовании утверждалось, что отсутствие соответствующих окаменелостей в маастрихте (72,1—66,0 млн лет назад — Маастрихтский ярус) было не результатом преждевременного исчезновения Cretoxyrhina mantelli, а просто пробелом в летописи окаменелостей. [C. G. Diedrich (2014). «Skeleton of the Fossil Shark Isurus denticulatus from the Turonian (Late Cretaceous) of Germany – Ecological Coevolution with Prey of Mackerel Sharks». Paleontology Journal. 2014: 1–20.]
Шимада (Shimada) и его коллега-палеонтолог Филипп Стернс (Phillip Sternes) опубликовали в 2018 году плакат, в котором высказали сомнение в достоверности этого предложения, отметив, что интерпретация исследования в значительной степени основана на данных, которые были выбраны произвольно и не цитировали ни Шимада (1997), ни Шимада (2005), которые являются ключевыми статьями относительно систематического положения вида Cretoxyrhina mantelli. [Phillip Sternes and Kenshu Shimada (2018), Caudal fin of the Late Cretaceous shark, Cretoxyrhina mantelli (Lamniformes: Cretoxyrhinidae), morphometrically compared to that of extant lamniform sharks, Society of Vertebrate Paleontology, archived from the original on 2018-12-05, retrieved 2018-12-05]
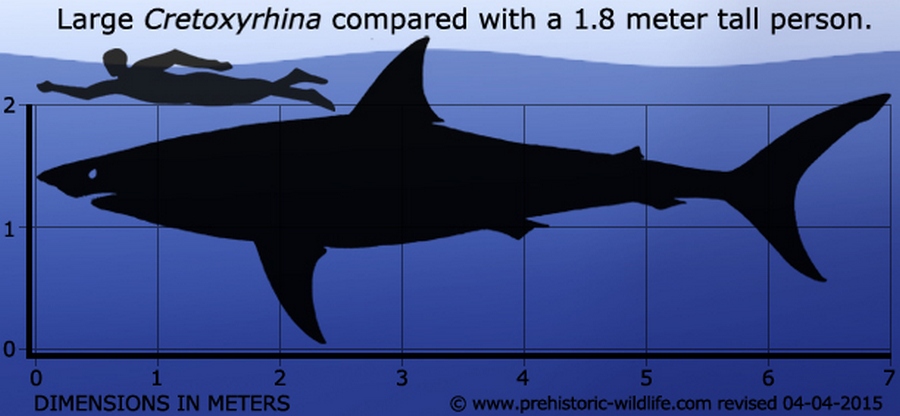
Кретоксирины — крупные акулы, в среднем 5—6 метров длиной и крупнейшими особями длиной более 7 метров (некоторые оценки указывают длину до 8—9 метров) и весом более 4944 кг (4,866—5,450 тонн). Cretoxyrhina была одной из крупнейших акул своего времени. По очертаниям тела кретоксирины напоминают современных белых акул (Carcharodon carcharias) и акул-мако (род Isurus). Зубы внешне сходны с зубами акул-мако (первоначально их включали в род Isurus / Oxyrhina). Однако строение эмали зубов отличается от такового у современных родов, эмаль гораздо толще, сами зубы шире. Зубы высотой до 7—8 см были похожи на бритву, покрыты толстой эмалью и предназначены для нанесения колющих ударов и резания добычи. Корни задних зубов широко расставлены, расстояние между ветвями в несколько раз превышает высоту коронки. Боковые зубчики на передних зубах могут отсутствовать. По краям челюстей зубы меньше, всего в челюстях около 80 зубов. Отмечено увеличение размера зубов этого рода от более древних канзасских фаун к более молодым. Кретоксирина была также одной из самых быстро плавающих акул, согласно гидродинамическим расчетам, способность к взрывной скорости достигала 70 километров в час. Было высказано предположение, что Cretoxyrhina охотилась, бросаясь на свою добычу на высокой скорости, чтобы нанести мощные удары, подобно большой белой акуле (Carcharodon carcharias), для этого она полагалась на хорошее зрение.
Отличительные характеристики зубов Cretoxyrhina включают почти симметричную или наклонную треугольную форму, острые лезвия без зубцов, видимые шейки зубов (bourlette — бурлет) и толстое эмалевое покрытие. Зубной ряд Cretoxyrhina (см. рисунок) обладает основными зубными характеристиками ламноидной акулы: зубные ряды расположены близко друг к другу без какого-либо перекрытия. Передние зубы прямые и почти симметричные, а боковые — скошенные. Сторона зуба, обращённая ко рту, выпуклая и имеет массивные выступы и питательные борозды на корне, тогда как лабиальная сторона, обращенная наружу, плоская или слегка вздутая. У молодых особей были боковые выступы на всех зубах, а у Cretoxyrhina vraconensis они стабильно сохранялись их в зрелом возрасте. Боковые выступы сохранялись только до всех латерозадних зубов во взрослом возрасте у Cretoxyrhina denticulata и Cretoxyrhina agassizensis и только до задних латерозадних зубов у Cretoxyrhina mantelli. Боковые выступы Cretoxyrhina vraconensis и Cretoxyrhina denticulata округлые, а у Cretoxyrhina agassizensis заострённые. Передние зубы Cretoxyrhina vraconensis имеют высоту 2,1—3,5 см, в то время как самый большой известный зуб Cretoxyrhina denticulata — 3 сантиметра. Зубы Cretoxyrhina mantelli больше, их средняя наклонная высота составляет 3—4 сантиметра. Самый большой зуб, обнаруженный у этого вида, мог иметь размер до 8 сантиметров.
Зубной ряд Cretoxyrhina mantelli (см. рисунок) является одним из самых известных среди всех вымерших акул благодаря ископаемым скелетам, таким как FHSM VP-2187 (см. рисунок, третий с верху), который состоит из почти полностью сочленённого зубного ряда. Другие скелеты Cretoxyrhina mantelli, такие как KUVP-247 (см. рисунок, сторой с верху) и KUVP-69102, также включают частичные челюсти с некоторыми зубами в их естественных положениях, некоторые из которых не присутствовали в более полных скелетах, таких как FHSM VP-2187. Используя эти образцы, Шимада (1997) реконструировал зубную формулу, а потом исправил её (Шимада, 2002):
S4.A2.I4.LP11 (+?) / s1?.a2.i1.lp15(+?).
Это означает, что спереди назад у Cretoxyrhina mantelli были: четыре симфизных зуба (маленькие зубы, расположенные в симфизе челюсти), два передних зуба, четыре промежуточных зуба и одиннадцать или более латерозадних зубов для верхней челюсти и, возможно, один симфизный зуб, два передних зуба, один промежуточный зуб и пятнадцать или более латерозадних зубов для нижней челюсти. Структура зубного ряда демонстрирует зубную структуру, подходящую для поведения при кормлении, аналогичную современным акулам мако (род Isurus), с большими копьевидными зубами в передней части, чтобы пронзить и зафиксировать добычу, и изогнутыми задними зубами, чтобы разрезать её на кусочки размером с укус. Этот механизм, часто неофициально описываемый палеонтологами, как «разрезание и нарезка кубиками». В 2011 году палеонтологи Джим Бурдон и Майк Эверхарт реконструировали зубной ряд нескольких особей Cretoxyrhina mantelli в зависимости от связанного с ними набора зубов. Они обнаружили, что две из этих реконструкций показывают некоторые заметные различия в размере первого промежуточного зуба (I1) и латеральных профилях, сделали вывод, что эти различия могут отражать половой диморфизм или индивидуальные вариации. [Jim Bourdon and Michael Everhart (2011). «Analysis of an associated Cretoxyrhina mantelli dentition from the Late Cretaceous (Smoky Hill Chalk, Late Coniacian) of western Kansas». Transactions of the Kansas Academy of Science. 114 (1–2): 15–32.]
Анализ черепа и чешуек позволяет предположить, что Cretoxyrhina mantelli имела коническую голову с плоским и широким черепом в дорсальной части. Рострум не выходил далеко вперед от переднего края черепной коробки, что позволяет предположить, что морда была тупой. Подобно современным акуле-крокодил (Pseudocarcharias kamoharai) и лисьим акулам (семейство Alopiidae), Cretoxyrhina mantelli имела пропорционально большие глаза, при этом орбита занимала примерно одну треть всей длины черепа, что давало ей острое зрение. Как хищник, хорошее зрение было важно при охоте на крупную добычу, которой питалась Cretoxyrhina. Напротив, менее развитые орбитальные, но более сильные обонятельные органы у более зависимых от запаха вороньих акул Squalicorax более подходили для оппортунистических падальщиков. Челюсти Cretoxyrhina mantelli были кинетически мощными. У них немного большая передняя кривизна и более мощное строение, чем у современных акул мако (род Isurus), но в остальном они были похожи по общей форме. Гиомандибула (hyomandibula) имеет удлинённую форму и, как полагали, может раскачиваться в стороны, что позволяло бы выпячивать челюсть и глубоко кусать. [Kenshu Shimada (1997). «Skeletal Anatomy of the Late Cretaceous Lamniform Shark, Cretoxyrhina mantelli, from the Niobrara Chalk in Kansas». Journal of Vertebrate Paleontology. 17 (4): 642–652.]
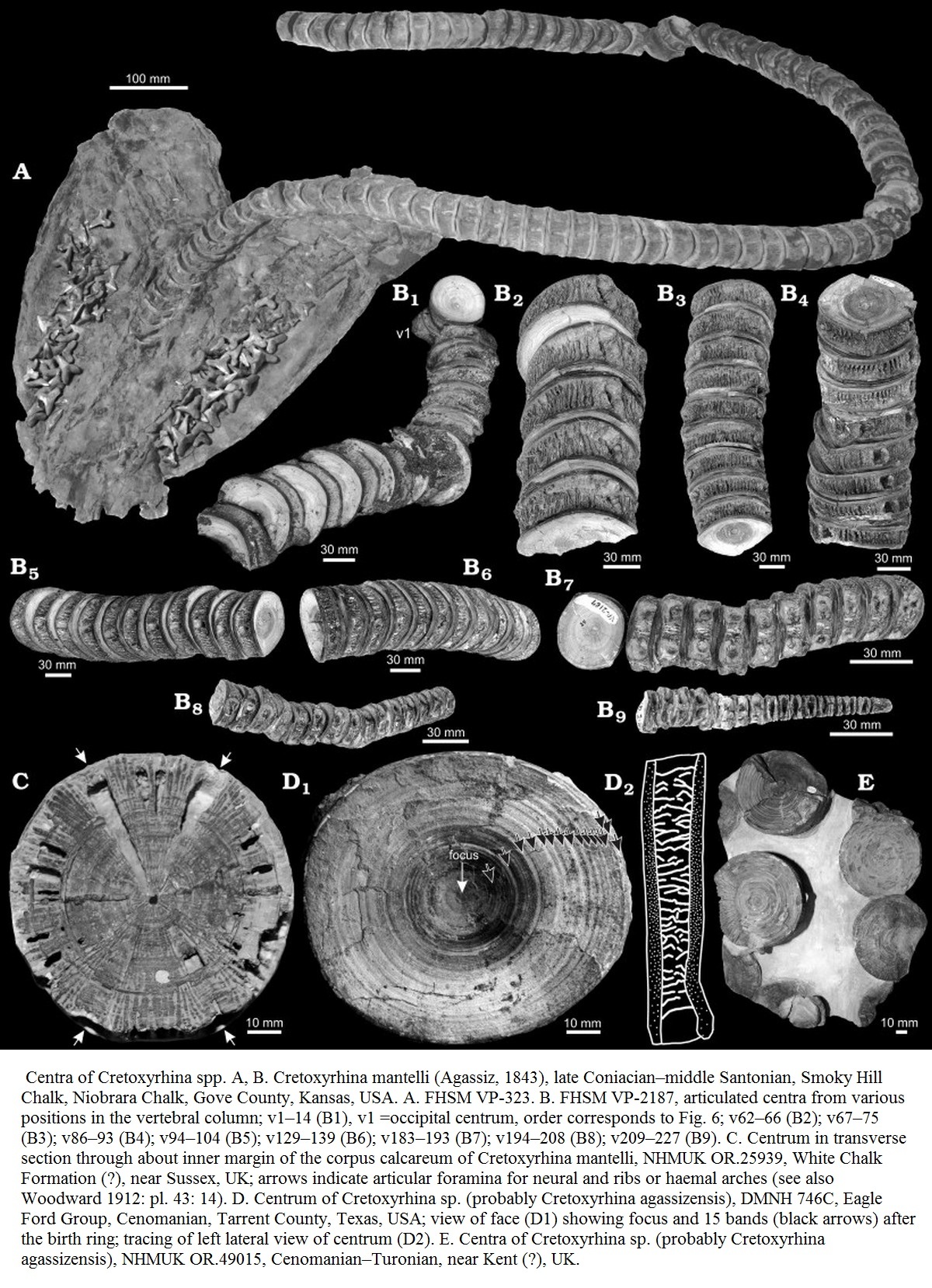
Большинство видов Cretoxyrhina представлены только ископаемыми зубами и позвонками. Как и у всех акул, скелет Cretoxyrhina был из хряща, который менее подвержен окаменению, чем кости. Тем не менее, окаменелости Cretoxyrhina mantelli из формации Ниобрара (Niobrara Formation) были обнаружены исключительно сохранившимися. Это произошло из-за того, что мел в формации имеет высокое содержание кальция, что делает кальцинирование хрящей более распространённой. При кальцинировании мягкие ткани затвердевают, что делает их более склонными к окаменению. Были обнаружены многочисленные скелеты, состоящие из почти целых позвоночных столбов. Диаметр самого большого позвонка составлял 87 миллиметров. Два экземпляра с наиболее хорошо сохранившимися позвоночными столбами (FHSM VP-2187 и KUVP 69102) имеют 218 и 201 позвонков, соответственно, и почти все позвонки в позвоночном столбе сохранились; у обоих отсутствует только часть кончика хвоста. При оценке длины хвоста общее количество позвонков составляет приблизительно 230 центров, что является уникальным, поскольку все известные ныне живущие ламноидные акулы обладают числом позвонков либо менее 197, либо более 282, без промежуточного числа. Позвонки в области туловища были большими и округлыми, создавая в целом веретеновидное, коренастое туловище. Анализ частично полной окаменелости хвостового плавника показывает, что у Cretoxyrhina он был в форме полумесяца, наиболее похожий на хвостовой плавник современных ламнид, китовых акул (Rhincodon typus) и гигантских акул (Cetorhinus maximus). Поскольку расположенные сзади ипохордальные лучи кажутся у Cretoxyrhina тоньше и короче по сравнению с таковыми у Rhincodon, Cetorhinus и у всех ламнид, доли полулунного хвоста у Cretoxyrhina mantelli могли быть немного уже, чем у современных акул. Переход к хвостовым позвонкам оценивается между 140-м и 160-м позвонками из 230, в результате чего общее количество хвостовых позвонков составляет 70—90 и составляет примерно 30—39% позвоночника (см. рисунок). Переход от прекаудальной (группы позвонков перед хвостовыми позвонками) к хвостовым позвонкам также отмечен позвоночным изгибом примерно на 45 °, что является самым высоким возможным углом, известным у современных акул и чаще всего встречается у быстро плавающих акул, таких как ламниды. Эти свойства хвоста, наряду с другими особенностями, такими как гладкая чешуя, параллельная оси тела, плезодический грудной плавник (грудной плавник, в котором простирается хрящ, что придаёт ему более надежную структуру, которая помогает уменьшить сопротивление) и веретеновидное, коренастое телосложение свидетельствует о способности Cretoxyrhina mantelli быстро плавать. [Kenshu Shimada, Stephen L. Cumbaa, and Deanne van Rooyen (2006). «Caudal Fin Skeleton of the Late Cretaceous Lamniform shark, Cretoxyrhina mantelli, from the Niobrara Chalk of Kansas». New Mexico Museum of Natural History Bulletin. 35: 185–191.]
Кретоксирина представляет собой одну из самых ранних форм и возможных источников эндотермии у ламноидных акул. Обладая региональной эндотермией (также известной как мезотермия), кретоксирина, возможно, обладала телосложением, аналогичным современным региональным эндотермическим акулам, таким как представители семейств лисьих акул и ламнид, где красные мышцы расположены ближе к оси тела по сравнению с эктотермическими акулами (чьи красные мышцы расположены ближе к окружности тела), и присутствует система специализированных кровеносных сосудов, называемая rete mirabile («чудесная сеть»), позволяющая сохранять метаболическое тепло и передавать его жизненно важным органам. Такое морфологическое строение позволяет акуле быть частично теплокровной и, таким образом, эффективно функционировать в более холодных водах, где была обнаружена Cretoxyrhina. Окаменелости были найдены в районах, где палеоклиматические оценки показывают температуру поверхности всего 5 ° C. Региональная эндотермия у кретоксирины, возможно, возникла в ответ на растущее давление со стороны прогрессирующего глобального похолодания и конкуренции со стороны мозазавров (семейство Mosasauridae) и других морских рептилий, которые также развили эндотермию. [Humberto G. Ferron (2017). «Regional endothermy as a trigger for gigantism in some extinct macropredatory sharks».]
Кретоксирина обладала очень плотными перекрывающимися плакоидными чешуйками, расположенными параллельно оси тела и покрытыми узорами из параллельных гребней, разделенными U-образными канавками, каждая из которых имела среднюю ширину около 45 микрометров (см. фото. Такое образование на чешуе позволяет эффективно снизить сопротивление и увеличить скоростные возможности — морфотип, наблюдаемый только у самых быстрых из известных акул. Таким образом, некоторые другие очевидные аспекты чешуйчатости Cretoxyrhina, такие как высокая плотность чешуек вместе с заметным наложением или наличие у чешуек постепенного истончения, также интерпретировались как адаптация для повышения гидродинамической эффективности у самых быстрых пелагических видов. Несколько гладких округлых чешуек также были описаны у Cretoxyrhina, но они, вероятно, ограничивались мордой и, возможно, другими участками, подверженными высокой абразивной нагрузке. Наконец, некоторые другие моменты, такие как коническая голова, форма и количество позвонков у Cretoxyrhina и отсутствие дорсо-вентрального уплощения в кретоламном позвонке, также указывают на существование веретеновидных тел у кретоксиринид, совместимых с активным пелагическим образом жизни. У Cretoxyrhina также был самый экстремальный случай хвостового плавника «Тип 4 — Type 4», имеющий самый высокий из известных углов Кобба (кривизна хвостовых позвонков) и угол изгиба хрящей хвоста (49 ° и 133 ° соответственно), когда-либо зарегистрированный у ламноидных акул. Конструкция хвостового плавника «Тип 4» представляет собой конструкцию с максимальной симметрией лопастей хвостового плавника, которая наиболее эффективна при быстром плавании, среди акул он встречается только у ламнид. В исследовании 2017 года, проведенном аспирантом Умберто Ферроном (Humberto Ferron), проанализированы взаимосвязи между морфологическими переменными, включая скелет и хвостовой плавник Cretoxyrhina mantelli, и смоделирована средняя крейсерская скорость 12 км/ч (7,5 миль в час) и взрывная скорость плавания около 70 км/ч (43 мили в час), что делает кретоксирину, возможно, одной из самых быстрых известных акул с высокой манёвренностью. Для сравнения, современная большая белая акула была смоделирована так, чтобы развивать скорость до 56 км/ч (35 миль/ч), а короткоперый мако (Isurus oxyrinchus), самая быстрая из ныне живущих акул, была смоделирована со скоростью 70 км/ч (43 мили в час).
Хотя никаких ископаемых свидетельств этого не было найдено, предполагается, что Cretoxyrhina была яйцеживородящей, как и все современные ламнообразные акулы. У яйцеживородящих акул детеныши вылупляются и растут внутри матери, конкурируя со своими собратьями через каннибализм, такой как оофагия (поедание яиц). Поскольку кретоксирина населяла олиготрофные и пелагические воды, где были обычными хищники, такие как крупные мозазавры, такие как тилозавры (род Tylosaurus), и макрохищные рыбы, такие как ксифактинусу (Xiphactinus audax), вполне вероятно, что это также была акула с r-селекцией, у которой рождались много младенцев, чтобы компенсировать высокий уровень смертности на ранних этапах развития. Точно так же обстоят дела у пелагических акул, таких как большая белая акула (Carcharodon carcharias), акулы-молотилки (семейство Alopiidae), акулы-мако (род Isurus), акулы-порбигли (Lamna nasus) и акулы-крокодилы (Pseudocarcharias kamoharai), производящие от двух до девяти младенцев в помёте. В экологии, теория r/K селекции (отбора) сводится к выбору компромисса между количеством и качеством потомства. По размеру затрат энергии, необходимой для размножения, виды организмов можно разделить на две группы: рудералы (r-стратеги) и конкуренты (К-стратеги). Акцент либо на увеличении количества потомства за счёт уменьшения индивидуальных родительских вложений r-стратегов (имеют невысокую продолжительность жизни, короткие жизненные циклы, небольшие размеры, высокую рождаемость, обычно один период размножения в течение жизни, способность к переживанию неблагоприятных периодов в состоянии покоя), либо на уменьшенном количестве потомства с соответствующим увеличенным родительским вложением K-стратегов (характеризуются низкой смертностью на ранних этапах развития и высокой продолжительностью жизни), широко варьируется, по-видимому, для достижения успеха в определенных условиях.
Как и все ламнообразные акулы, Cretoxyrhina каждый год наращивает на своих позвонках кольцо роста. Шимада (Shimada, 1997) вывел линейное уравнение для расчета общей длины тела кретоксирины с использованием диаметра тела позвонка, которое показано ниже, где TL представляет общую длину тела, а CD представляет диаметр каждого кольцо роста.
TL = 0,281 + 5,746CD
Используя это линейное уравнение, Шимада (2008) впервые провел измерения на наиболее хорошо сохранившемся образце Cretoxyrhina mantelli, FHSM VP-2187 (см. рисунок, третий с верху). Измерения показали, что при рождении длина акулы составляла 1,28 метра и вес около 16,3 кг, а также был отмечен быстрый рост в первые два года жизни, удвоение длины в течение 3,3 лет. С тех пор рост становился устойчивым и постепенным, средняя длина увеличивалась на 21,1 сантиметра в год, пока акула не умерла в возрасте примерно 15 лет, когда выросла до 5 метров. Используя модель роста фон Берталанфи (von Bertalanffy) для FHSM VP-2187, максимальная продолжительность жизни Cretoxyrhina mantelli была оценена в 38,2 года. К этому возрасту акула вырастет более 7 метров в длину. Основываясь на аллометрическом измерении большой белой акулы, Шимада обнаружил, что такая особь весила бы до 3400 килограммов (3,3—3,7 тонны).
Повторное измерение, проведенное Ньюбреем и соавторами (Newbrey et al., 2013), обнаружило, что Cretoxyrhina mantelli и Cretoxyrhina agassizensis достигали половой зрелости в возрасте от четырех до пяти лет, и предложили возможный пересмотр измерений годичных колец в FHSM VP-2187. Продолжительность жизни FHSM VP-2187 и максимальную продолжительность жизни Cretoxyrhina mantelli также предлагалось увеличить до 18 и 21 года соответственно с использованием новых измерений. В исследовании 2019 года, проведенном итальянским ученым Джакопо Амальфитано (Jacopo Amalfitano), было проведено краткое измерение позвонков по двум окаменелостям Cretoxyrhina mantelli, и было обнаружено, что пожилая особь умерла примерно в возрасте 26 лет. Измерения также проводились на других скелетах Cretoxyrhina mantelli и позвонке Cretoxyrhina agassizensis, что дало результаты с аналогичными темпами быстрого роста на ранних этапах жизни. Такой быстрый рост в течение нескольких лет мог бы помочь кретоксиринам выживать за счет быстрого выхода из периода младенчества из-за его уязвимости, поскольку у взрослых акул будет мало естественных врагов. Исследование синтипного зуба Cretoxyrhina mantelli из Англии также выявило и рассчитало максимальную длину индивидуума — 8 метров, что сделало зуб самым большим из известных образцов. При применении аллометрической шкалы, используемой Шимада (2008), Cretoxyrhina mantelli такой длины дает предполагаемую массу тела около 4944 кг (4,866—5,450 тонн).
Мощные кинетические челюсти, высокая скорость и большой размер кретоксирин позволяют предположить, что это очень агрессивный хищник. Связь кретоксирины с разнообразным количеством окаменелостей, показывающих признаки пожирания, подтверждает, что это был активный высший хищник, который питался большей частью разнообразной морской мегафауны в позднем меловом периоде. Наивысший трофический уровень, который занимали эти акулы, был разделён только с крупными мозазаврами, такими как Тилозавр (род Tylosaurus), во время последних стадий позднего мелового периода. Кретоксирина в основном охотилась на других активных хищников, включая ихтиодектид (Ichthyodectidae — семейство лучепёрых крупных рыб, включая ксифактинов — Xiphactinus), плезиозавров (отряд Plesiosauria, Plesiosaurs), черепах (отряд Testudines), мозазавров (семейство Mosasauridae), и других акул. Большинство окаменелостей кретоксирин, питающихся другими животными, состоят из больших и глубоких следов укусов и проколов на костях, иногда с врезанными в них зубами. Изолированные кости мозазавров и других морских рептилий, которые были частично растворены в результате пищеварения или обнаружены в копролитах, также являются примерами питания кретоксирины. Известно несколько скелетов Cretoxyrhina, содержащих содержимое желудка, в том числе большой скелет Cretoxyrhina mantelli (KUVP 247), в котором в области желудка находятся скелетные остатки большого ихтиодектида Xiphactinus и мозазавра. Кретоксирина могла иногда питаться птерозаврами, о чём свидетельствует набор шейных позвонков птеранодона из формации Ниобрара с глубоко расположенным между ними зубом Cretoxyrhina mantelli (см. рисунок). Хотя нельзя с уверенностью сказать, было ли окаменелость результатом хищничества или поедания падали, вполне вероятно, что птеранодон и подобные птерозавры были естественными целями для таких хищников, как Cretoxyrhina, поскольку они обычно заходили в воду, чтобы питаться, и, таким образом, были в пределах досягаемости.
Хотя Cretoxyrhina в основном была активным охотником, она также была приспособленцем и, возможно, время от времени собирала падаль. Многие окаменелости со следами кормления Cretoxyrhina не показывают признаков заживления, что свидетельствует о преднамеренном нападении хищников на живое животное, и вероятно, что, по крайней мере, некоторые следы кормления были сделаны в результате нападения при активной охоте. Остатки частичных скелетов динозавров, таких как клаозавр (род Claosaurus) и ниобраразавр (род Niobrarasaurus), демонстрируют признаки питания и переваривания Cretoxyrhina mantelli. Скорее всего, из-за палеогеографического местоположения окаменелостей в открытом океане, это были мертвые туши, попавшие в океан.
Стратегии охоты Cretoxyrhina недостаточно хорошо документированы, потому что многие окаменелости с отметками кормления Cretoxyrhina не могут быть различимы между хищничеством или падалью. Если они действительно были результатом первого, это означало бы, что кретоксирина, скорее всего, использовала стратегию охоты, включающую главный мощный и смертельный удар, подобный тарану, наблюдаемому у современных серых акул (семейство Carcharhinidae) и ламнидов (семейство Lamnidae). В исследовании 2004 года, проведённом экспертами по акулам Витторио Габриотти (Vittorio Gabriotti) и Алессандро Де Маддалена (Alessandro De Maddalena), было отмечено, что современная большая белая акула (Carcharodon carcharias), достигающая в длину более 4 метров, обычно таранит свою добычу с огромной скоростью и силой, чтобы нанести одиночные смертельные удары, иногда настолько мощные, что жертва будет вытолкнут из воды силой удара. Поскольку Кретоксирина обладала крепким коренастым телосложением, способным к быстрому плаванию, мощными кинетическими челюстями, как у большой белой акулы, и достигала такой же или большей длины, такой стиль охоты был бы вероятен.
Кретоксирина имела космополитическое распространение, окаменелости были обнаружены по всему миру. Известные места включают Северную Америку, Европу, Израиль и Казахстан (карта). Cretoxyrhina в основном обитала в умеренном и субтропическом поясах. Она была обнаружена в широтах вплоть до 55 ° северной широты, где по палеоклиматическим оценкам средняя температура поверхности моря составляет 5—10 ° C . Окаменелости кретоксирины наиболее известны в районе Западного Внутреннего моря, который сейчас находится в центральной части Соединенных Штатов и Канады. Во время выступления в 2012 году Микаэль Сиверсон (Mikael Siverson) отметил, что особи Cretoxyrhina, живущие в открытом море, обычно были крупнее тех, что населяли Западный внутренний морской путь, а некоторые из окаменелостей Cretoxyrhina mantelli в открытом море имеют общую длину до 8 метров, возможно, 9 метров.
Кретоксирина населяла в основном пелагические океаны от умеренного до субтропического пояса. Зуб кретоксирины, обнаруженный в формации Подковообразного каньона (Horseshoe Canyon Formation) в Альберте (формация, где единственные найденные водные отложения происходят из солоноватой воды и не имеют океанического происхождения), даёт повод предположить, что кретоксирина могла иногда заходить в частично пресноводные устья рек и в другие подобные водоёмы. Возможно, вынос нижележащего слоя (переработанные окаменелости) может быть более вероятным объяснением такого явления. Климат морских экосистем во временном диапазоне кретоксирины был в целом намного теплее, чем в наши дни, из-за более высоких уровней углекислого газа и других парниковых газов в атмосфере, на которые в то время влияла форма континентов. В интервале сеномана и турона 97—91 млн лет назад наблюдался пик температуры поверхности моря, в среднем более 35 °, и температуры придонной воды около 20 ° C, на 7—8 ° C теплее, чем в наши дни. Примерно в это же время Cretoxyrhina сосуществовала с взрывным увеличением разнообразия фауны, например мозазавров (семейство Mosasauridae). Этот интервал также включал рост глобальных уровней стабильных изотопов углерода 13, что свидетельствует о значительном истощении кислорода в океане и вызвало аноксическое событие (кислородное обеднение) в сеномане и туроне. Хотя это событие привело к исчезновению до 27% морских беспозвоночных, позвоночные животные, такие как Cretoxyrhina, в целом не пострадали. В остальные временные промежутки мелового периода наблюдалось прогрессирующее глобальное похолодание земных океанов, что привело к появлению умеренных экосистем и возможному оледенению в раннем маастрихте (72,1—66,0 млн лет назад — Маастрихтский ярус). Субтропические районы сохранили высокое биоразнообразие всех таксонов, в то время как экосистемы умеренного пояса, как правило, имели гораздо меньшее разнообразие. В Северной Америке в субтропических провинциях преобладали акулы, черепахи и мозазавры, такие как Tylosaurus и Clidastes, в то время как в провинциях с умеренным климатом преобладали плезиозавры, морские птицы гесперорнитиды и мозазавры Platecarpus.
Кретоксирина жила рядом со многими хищниками, которые имели сходный трофический уровень в разнообразной пелагической экосистеме в меловом периоде. В число этих хищников входили крупные морские рептилии, некоторые из которых уже занимали высший трофический уровень, когда впервые появилась Cretoxyrhina. В период от альба до турона, примерно от 107 до 91 млн лет назад, Cretoxyrhina была современником и сосуществовала вместе с плиозаврами (род Pliosaurus) среднего мела. Некоторые из этих плиозавров, например, мегацефалозавр (род Megacephalosaurus) достигали длины 9 метров. К среднему турону, около 91 млн лет назад, плиозавры вымерли. Считается, что увеличение разнообразия видового состава фауны, могло быть основным фактором, способствовавшим ускорению исчезновения таких акул, как Cretoxyrhina. Экологическая пустота, которую они оставили, была быстро заполнена появляющимися мозазаврами (семейство Mosasauridae), которые также стали занимать самые высокие трофические уровни. Крупные мозазавры, такие как тилозавры (род Tylosaurus), достигавшие в длину более 14 метров, могли конкурировать с Cretoxyrhina, и были обнаружены доказательства межвидовых взаимодействий, такие как следы укусов любого из них. Было также много акул, которые играли с кретоксириной аналогичную экологическую роль, такие как кардабиодоны (род Cardabiodon) и двардиусы (род Dwardius), последний демонстрирует доказательства прямой конкуренции с Cretoxyrhina vraconensis на основе сложных схем взаимодействия между ними.
В исследовании 2010 года палеонтологов Коринн Майерс (Corinne Myers) и Брюса Либермана (Bruce Lieberman) о конкуренции в Западном внутреннем море использовались количественные аналитические методы, основанные на географических информационных системах и тектонических реконструкциях, чтобы восстановить гипотетические конкурентные отношения между десятью наиболее распространенными и многочисленными морскими позвоночными в регионе, включая Cretoxyrhina. Их расчеты обнаружили отрицательную корреляцию между распределением Cretoxyrhina и тремя потенциальными конкурентами Squalicorax kaupi, Tylosaurus proriger и Platecarpus spp.; статистически значимая отрицательная корреляция означает, что на распространение одного вида повлияло то, что он уступил место другому. Однако ни одна из взаимосвязей не была статистически значимой, этого указывает на то, что вымирание вряд ли было результатом конкуренции.
Причины исчезновения Cretoxyrhina неясны. Что известно, так это то, что численность медленно снижалась на протяжении миллионов лет. С момента своего пика в размерах и в распространении в коньяке — 89,8—86,3 млн лет назад (Коньякский ярус) количество окаменелостей Cretoxyrhina постепенно уменьшалось до окончательного вымирания во время кампана — 83,6—72,1 млн лет назад (Кампанский ярус). Сиверсон и Линдгрен (Siverson and Lindgren, 2005) отметили, что возраст самых молодых окаменелостей Cretoxyrhina различается между континентами. В Австралии самые молодые окаменелости Cretoxyrhina были датированы приблизительно 83 млн лет назад во время сантона, в то время как самые молодые североамериканские окаменелости, известные в то время (которые были датированы ранним кампаном), были как минимум на два миллиона лет старше, чем самые молодые окаменелости в Европе. Различия между возрастами предполагают, что Cretoxyrhina в таких областях со временем, возможно, исчезала локально, пока не вымер весь род в целом.
Было отмечено, что упадок Cretoxyrhina совпадает с появлением новых хищников, таких как тилозавры (род Tylosaurus), предполагая, что возрастающее давление со стороны конкуренции с мозазаврами и другими хищниками аналогичного трофического уровня, возможно, сыграло серьезный вклад в упадок Cretoxyrhina и возможное вымирание. Другим возможным фактором было постепенное обмеление и сокращение площади Западного внутреннего моря, что привело к исчезновению пелагических сред, предпочитаемых акулой; этот фактор не объясняет упадок и исчезновение Cretoxyrhina где-либо ещё. Было высказано предположение, что исчезновение Cretoxyrhina могло способствовать дальнейшему увеличению разнообразия мозазавров (семейство Mosasauridae).
До появления специализированных форм мозазавров (род Mosasaurus), таких как тилозавр (Tylosaurus, см. рисунок), кретоксирина, вероятно, была одним из доминирующих водных хищников в период после вымирания крупных плиозавров (семейство Pliosauridae). Прочные, но гладкие зубы кретоксирин едва ли позволяли им убивать и разделывать относительно крупных животных эффективнее, чем это делают современные акулы-мако (род Isurus). Это говорит о том, что в первую очередь кретоксирины были ихтиофагами и обычно проглатывали свою жертву целиком. Обычной пищей для этих акул были многочисленные костные рыбы, вплоть до небольших ксифактинов (Xiphactinus audax, см. рисунок), а также меньшие акулы, птерозавры (отряд Pterosauria, см. рисунок) и иногда динозавры (см. рисунок). Однако, также известны следы от зубов кретоксирин (и сами зубы, воткнувшиеся в кость) на костях мелких мозазавров (см. рисунок), на панцирях морских черепах (см. рисунок) и, возможно, на плавнике 6-метрового подростка эласмозавра (род Elasmosaurus). Вместе с остатками скелетов кретоксирин также были найдены остатки полупереваренных костей этих животных. Поэтому кажется весьма вероятным, что эти акулы, также как и скваликораксы (семейство Anacoracidae), часто питались трупами морских ящеров. Косвенно тягу кретоксирин к поеданию падали также подтверждают следы от акульих зубов на костях гадрозавров (Hadrosaurus foulkii) из среднего мела Канзаса (трупы динозавров попадали в море и поедались акулами). Несмотря на это, известно два случая обнаружения позвонков некрупных мозазавров (род Mosasaurus), имеющих на себе следы от укусов кретоксирин со следами заживления. Одна из этих находок может говорить о том, что при большой разнице в размерах кретоксирины всё-таки могли атаковать морских рептилий, также как акулы мако порой атакуют небольших морских млекопитающих. В то время как другая скорее демонстрирует травму, полученную мозазавром при нападении на кретоксирину (кусающая акула была около 3 метров в длину, а платекрап (Platecarpus tympaniticus) или молодой тилозавр — около 6 метров) и самообороне последней, либо случайный укус акулы при совместном с морским ящером кормлении.
Сами кретоксирины, возможно, иногда становились жертвой больших и средних по размерам мозазавров (род Mosasaurus), а факт их вымирания часто объясняется расцветом этих морских рептилий, пришедших на смену вымершим ихтиозаврам (отряд Ichthyosauria) и плиозаврам (род Pliosaurus).
Как высший хищник, Cretoxyrhina играла решающую роль в морских экосистемах, в которых она обитала. Это был космополитический род, и его окаменелости были обнаружены по всему миру, хотя чаще всего в районе Западного внутреннего морского пути в Северной Америке. Кретоксирины предпочитали в основном субтропическую или умеренную пелагическую среду, но были также известны в водах с температурой 5 ° C. Пик расцвета рода — 89,8—86,3 млн лет назад (Коньякский ярус), но впоследствии род переживал непрерывный спад численности видов до их исчезновения во время кампана (83,6—72,1 млн лет назад — Кампанский ярус). Одним из факторов этой кончины могло быть усиление давления со стороны конкуренции с хищниками, которые возникли примерно в то же время, в первую очередь гигантский мозазавр — тилозавр (род Tylosaurus). Другие возможные факторы включают постепенное исчезновение Западного внутреннего моря (Western Interior Seaway — древнее эпиконтинентальное море, существовавшее с середины до конца мелового периода на территории современных Канады и США, разделяя её на Ларамидию и Аппалачию).
• † Cretoxyrhina agassizensis Underwood & Cumbaa, 2010 [синоним Telodontaspis agassizensis] (Кретоксирина Агассиса) — середина сеномана (100,5—93,9 млн лет назад — Сеноманский ярус) — середина турона (93,9—89,8 млн лет назад — Туронский ярус), Канада (провинция Манитоба), западная Австралия, США (группа Eagle Ford, округ Таррант, Техас). В 2010 году британский и канадский палеонтологи Чарли Андервуд (Charlie Underwood) и Стивен Кумба (Stephen Cumbaa) описали Telodontaspis agassizensis из зубов, найденных в озере Агассис (Lake Agassiz) в Манитобе, Канада, которые ранее были идентифицированы как молодые зубы акулы из рода Cretoxyrhina. [Underwood, C.J. & Cumbaa, S.L. (2010) Chondrichthyans from a Cenomanian (Late Cretaceous) bonebed, Saskatchewan, Canada. Palaeontology, 53 (4): 903–944]
Этот вид был вновь включен в род Cretoxyrhina в результате исследования, проведенного в 2013 году американским палеонтологом Майклом Ньюбри (Michael Newbrey) с использованием дополнительных ископаемых материалов того же вида, найденного в Западной Австралии (мергель Хейкок, плато Пиллаварра). [Michael G. Newbrey, Mikael Siversson, Todd D. Cook, Allison M. Fotheringham, and Rebecca L. Sanchez (2013). «Vertebral Morphology, Dentition, Age, Growth, and Ecology of the Large Lamniform Shark Cardabiodon ricki». Acta Palaeontologica Polonica. 60 (4): 877–897.]
Cretoxyrhina agassizensis произошла в середине сеномане (100,5—93,9 млн лет назад — Сеноманский ярус) от Cretoxyrhina denticulata из раннего сеномана. Примечательно, что Cretoxyrhina agassizensis существовала до среднего турона (93,9—89,8 млн лет назад — Туронский ярус) и на короткое время была современником Cretoxyrhina mantelli.
Cretoxyrhina agassizensis (Underwood & Cumbaa, 2010) может быть диагностирована как вид с очень тонким бугорком на передних зубах (см. фото) в ювенильной и суб-взрослой стадии (более широкий на той же высоте у более старых Cretoxyrhina denticulata (Glikman, 1957) и более молодых Cretoxyrhina mantelli), наличие зубцов с острой вершиной на многих латерально-передних зубах (не закругленных, как это обычно бывает в топотипе Cretoxyrhina denticulata, см. фото) и отсутствие полных режущих кромок на маленьких ювенильных передних зубах. Губная поверхность бугорка в передних зубах все больше сужается в основании с уменьшением возраста Cretoxyrhina agassizensis.
Зуб WAM 13.6.2 из базального слоя марля Хейкок, нижняя часть реки Мерчисон, западная Австралия, неотличим от передних зубов Telodontaspis agassizensis Underwood & Cumbaa, 2010. Узкие срединные борозды вдоль нижней части базальная поверхность корня является общей чертой ранней кретоксирины (Siverson and Lindgren 2005; Siverson et al. 2013; Underwood и Cumbaa 2010, ошибочно идентифицирован как зуб Roulletia). Подобные борозды присутствуют на голотипе Telodontaspis agassizensis (Underwood & Cumbaa 2010), описанном из пачки Belle Fourche (вероятно, зона Acanthoceras amphibolum позднего и среднего сеномана) формации Эшвилл, Саскачеван, Канада. Номинальный Telodontaspis встречается вместе с более крупной типичной Cretoxyrhina в его типовой местности и в базальной части Haycock Marl. Постепенный переход базального сужения губной поверхности бугорка передних зубов от морфологии «Telodontaspis» через «юную» стадию кретоксирины до стадии суб-взрослого или взрослого и идентичные борозды на основной поверхности корня у двух номинальных таксонов указывают на то, что родовые названия обозначают разные онтогенетические стадии у одного вида. На синонимию также указывает тот факт, что Telodontaspis не встречается в отложениях, в которых отсутствуют типичные большие зубы Cretoxyrhina. Как указано Siverson et al. (2013), онтогенетическая гетеродональность очень выражена у Cretoxyrhina, и, по-видимому, Андервуд и Кумба (2010) недооценили степень онтогенетической гетеродонности в течение первых нескольких лет у особей этого рода. Самый ранний образец этого вида встречается в пачке Belle Fourche формации Ashville (описанной как Cretoxyrhina denticulata, Telodontaspis agassizensis и Roulletia canadensis, тогда как самое молодое месторождение находится в раннем среднем туроне (93,9—89,8 млн лет назад — Туронский ярус) фейрпортской пачки карлильских сланцев (описанных Сиверсоном и Линдгреном 2005 как Cretoxyrhina mantelli). Образец среднего сеномана (100,5—93,9 млн лет назад — Сеноманский ярус) Cretoxyrhina из самого верхнего алевролита Gearle в CY Creek, Giralia Anticline (типовой пласт для Cardabiodon ricki) включает передние зубы типа Cretoxyrhina agassizensis и латерозадние зубы типа Cretoxyrhina denticulata (бугорки присутствуют на всех обычно округлых задних зубах), и возможно, что эти два вида являются хроносегментами одной линии (т. е. Cretoxyrhina denticulata от раннего до раннего среднего сеномана, за которым следует Cretoxyrhina agassizensis с конца среднего сеномана до начала среднего турона).
• † Cretoxyrhina denticulata Glikman, 1957 [синонимы Cretoxyrhina cf. denticulata, Isurus denticulatus, Pseudoisurus denticulatus] произошла в раннем сеномане (100,5—93,9 млн лет назад — Сеноманский ярус) от Cretoxyrhina vraconensis. В 1957 году советский палеонтолог Леонид Гликман (Leonid Glickman) идентифицировал второй вид рода Cretoxyrhina на основе некоторых из более ранних зубов кретоксирин, которые он назвал Cretoxyrhina denticulata. [Glikman, L.S. (1957) (Taxonomic significance of the accessory teeth of sharks of the families Lamnidae and Scapanorhynchidae). «in russian». Trudy geol. Muz. «A.P. Karpinskogo» Akad. Nauk S.S.S.R., 1: 103–109, 2 fig.]
Название вида denticulata происходит от латинского слова «denticulus» — «маленький зуб» и суффикса «ata» — «владение», что вместе означает «наличие маленьких зубов». Это отсылка к появлению боковых выступов (бугорков) на большинстве зубов Cretoxyrhina denticulata.
Cretoxyrhina denticulata (Glikman, 1957) может быть диагностирован как вид более широкими и закруглёнными бугорками на передних зубах в ювенильной и суб-взрослой стадии (см. фото) по сравнению с очень тонкими выступами с острой вершиной на многих латерозадних зубах у более молодой Cretoxyrhina agassizensis (Underwood and Cumbaa, 2010) (см. фото). Морфология зубов изменилась в течение альба — турона от разрывающего (Cretoxyrhina denticulatus) до режущего (Cretoxyrhina mantelli) типа (коньяк — кампан).
• † Cretoxyrhina mantelli Agassiz, 1843 [синонимы Isurus mantelli (Agassiz, 1843), Oxyrhina mantelli Agassiz, 1843] (Ginsu shark — Акула Гинсу, Кретоксирина Мантелла) — типовой вид, 94,3—66,043 млн лет назад, меловой период Канада (Саскачеван), Конго-Браззавиль, Франция, Германия, Иордания, Швеция, США (Алабама, Колорадо, Канзас, Нью-Джерси), является наиболее известным видом из рода Cretoxyrhina. Описан по нескольким довольно полным скелетам. Самые полные остатки происходят из отложений Канзасского мелового моря, возрастом 100—90 миллионов лет (сеноман — сантон).
Видовой эпитет бинома mantelli переводится как «Мантелла», который Луи Агассис дал в честь английского палеонтолога Гидеона Мантелла (Gideon Mantell) за то, что он предоставил ему синтипы этого вида.
М. Эверхарт (Mike Everhart) назвал эту акулу «Акулой Гинсу», в честь знаменитой в США марки ножей, поскольку теоретический механизм питания акулы с большими копьевидными зубами в передней части, чтобы пронзить и зафиксировать добычу, и изогнутыми задними зубами, чтобы разрезать её на кусочки размером с укус часто сравнивают с «разрезанием и нарезкой кубиками» при использовании ножа.
Cretoxyrhina mantelli произошла в позднем сеномане (100,5—93,9 млн лет назад — Сеноманский ярус) от Cretoxyrhina agassizensis из среднего сеномана. Примечательно, что Cretoxyrhina agassizensis существовала до среднего турона (93,9—89,8 млн лет назад — Туронский ярус) и на короткое время была современником Cretoxyrhina mantelli. Зубы Cretoxyrhina mantelli больше, чем у других видов рода, их средняя наклонная высота составляет 3—4 сантиметра. Самый большой зуб, обнаруженный у этого вида, мог иметь размер до 8 сантиметров. Размер зубов у особей Cretoxyrhina mantelli в пределах Западного внутреннего моря достиг своего пика около 86 миллиона лет назад во время последнего коньякского периода, а затем начинает медленно уменьшаться. В Европе этот пик приходится на более ранний период позднего турона. Самая молодая окаменелость Cretoxyrhina mantelli была обнаружена в формации Медвежья Лапа (Bearpaw Formation) в провинции Альберта, Канада, возраст которой составляет 73,2 миллиона лет.
У молодых особей были боковые выступы на всех зубах, а у Cretoxyrhina vraconensis они стабильно сохранялись их в зрелом возрасте. Боковые выступы сохранялись только до всех латерозадних зубов во взрослом возрасте у Cretoxyrhina denticulata и Cretoxyrhina agassizensis и только до задних латерозадних зубов у Cretoxyrhina mantelli. Боковые выступы Cretoxyrhina vraconensis и Cretoxyrhina denticulata округлые, а у Cretoxyrhina agassizensis заострённые. Передние зубы Cretoxyrhina vraconensis имеют высоту 2,1—3,5 см, в то время как самый большой известный зуб Cretoxyrhina denticulata — 3 сантиметра. Зубы Cretoxyrhina mantelli больше, их средняя наклонная высота составляет 3—4 сантиметра. Самый большой зуб, обнаруженный у этого вида, мог иметь размер до 8 сантиметров.
Анализ черепа и чешуек позволяет предположить, что Cretoxyrhina mantelli имела коническую голову с плоским и широким черепом в дорсальной части. Рострум не выходил далеко вперед от переднего края черепной коробки, что позволяет предположить, что морда была тупой. Подобно современным акуле-крокодил (Pseudocarcharias kamoharai) и лисьим акулам (семейство Alopiidae), Cretoxyrhina mantelli имела пропорционально большие глаза, при этом орбита занимала примерно одну треть всей длины черепа (см. рисунок), что давало ей острое зрение. Как хищник, хорошее зрение было важно при охоте на крупную добычу, которой питалась Cretoxyrhina. Челюсти Cretoxyrhina mantelli были кинетически мощными. У них немного большая передняя кривизна и более мощное строение, чем у современных акул мако (род Isurus), но в остальном они были похожи по общей форме. Гиомандибула (hyomandibula) имеет удлинённую форму и, как полагали, может раскачиваться в стороны, что позволяло бы выпячивать челюсть и глубоко кусать. [Kenshu Shimada (1997). «Skeletal Anatomy of the Late Cretaceous Lamniform Shark, Cretoxyrhina mantelli, from the Niobrara Chalk in Kansas». Journal of Vertebrate Paleontology. 17 (4): 642–652.]
С конца 19 века в Канзасе было обнаружено несколько окаменелостей исключительно хорошо сохранившихся скелетов кретоксирин. Учёные успешно рассчитали её жизненный цикл, используя позвонки некоторых скелетов. Кретоксирина быстро росла в раннем возрасте и достигала половой зрелости в возрасте от четырех до пяти лет. По расчетам, её срок жизни составлял почти сорок лет. Анатомический анализ скелетов кретоксирин показал, что акулы обладали оптическими особенностями наиболее близкими к лисьим акулам (семейство Alopiidae).
• † Cretoxyrhina vraconensis Zhelezko, 2000 [синонимы Cretoxyrhina aff. vraconensis, Pseudoisurus vraconensis Zhelezko, 2000]. Самые ранние окаменелости кретоксирин датируются 107 миллионами лет и принадлежат Cretoxyrhina vraconensis. Железко описал этот вид, родственный Cretoxyrhina mantelli, на основе материала зубов из Казахстана, который он определил как Pseudoisurus vraconensis в соответствии с его таксономической переоценкой. [Zhelezko, V.I. (2000) (The evolution of teeth system of sharks of the genus Pseudoisurus Glückman, 1957 - the biggest pelagic sharks of Eurasia) «in russian». Materialy po Stratigrafii i Paleontologii Urala, 4: 136–141, pl. 1–4.]
Исследование, проведенное в 2013 году куратором и палеонтологом музея Западной Австралии (Western Australian Museum) Микаэлем Сиверсоном (Mikael Siverson), исправило таксономическую ошибку, восстановив род Cretoxyrhina и поместив вид Pseudoisurus vraconensis в него. Род Cretoxyrhina начал развиваться благодаря Cretoxyrhina vraconensis, эволюционирующей в Cretoxyrhina denticulata в течение раннего сеномана (100,5—93,9 млн лет назад — Сеноманский ярус), которая в середине сеномана превратилась в Cretoxyrhina agassizensis.
Морфология зубов из отложений Канева (Приднепровье, Украина) согласуется с таковой у Cretoxyrhina vraconensis (Сиверсон и др., 2013) и отличается от «Pseudoisurus» vraconensis (Glikman, 1957) и Cretoxyrhina mantelli (Agassiz, 1838) развитыми треугольными боковыми выступами на боковых сторонах и редуцированными выступами на передних зубах, округлыми долями корня и одним или двумя вертикальными лабиальными выступами на зубах базальной (низкой) части коронки.
Голотип номинальной акулы «Pseudoisurus» vraconensis включает отдельный зуб из позднеальбской толщи (113,0—100,5 млн лет назад — Альбский ярус) районе колодца Бесакты (Besakty), Мангышлак, Казахстан. На уровне видов этот зуб неотличим от отдельных зубов из зоны Mortoniceras rostratum сланца Pawpaw на северо-востоке Техаса, США. Его стратиграфическое происхождение указывает на то, что он одного возраста или несколько старше техасских образцов. Большое количество зубов типа «Pseudoisurus» vraconensis из несколько более молодых слоев вблизи границы альба и сеномана в Колбае (Kolbay), восточный Мангышлак, Казахстан, позволил реконструировать зубной ряд в соответствии со стоматологическим шаблоном Cretoxyrhina, уточняя родовую принадлежность Pseudoisurus vraconensis. В образцах из Кольбая дистальная доля корня трёх возможных третьих верхних передних зубов не сдавлена лабиолингвально по сравнению с мезиальной долей. Лабиолингвальное уплощение корневой доли, обращенной к мезиальной стороне промежуточной перемычки, является характерной чертой современных ламнид. Промежуточная перемычка увеличивает структурную прочность передней части нёбной кости, одновременно изолируя верхнюю переднюю часть зуба, способствуя глубокому проникновению за счёт их увеличенных бугорков.
Особи Cretoxyrhina от позднего коньяка (89,8—86,3 млн лет назад — Коньякский ярус) до раннего кампана (83,6—72,1 млн лет назад — Кампанский ярус) участка Smoky Hill Chalk из формации Niobrara Chalk в Канзасе, США, демонстрируют сильно разобщённую гетеродонтию в передней половине зубного ряда верхней челюсти. Этот так называемый образец ламноидных зубов рассматривается некоторыми исследователями как синапоморфия Lamniformes. Карликовые зубы, образующие ламноидный зубной узор у образцов Smoky Hill Chalk, по-видимому, соответствуют относительно более крупным соответствующим зубам у Cretoxyrhina vraconensis, старше на 15 миллионов лет. Вероятное ослабление неоднородности в верхней челюсти в Cretoxyrhina с увеличением геологического возраста указывает на то, что структура ламноидных зубов могла развиваться независимо в двух или более ламниформных кладах.
 1f teeth (Cretoxyrhina agassizensis).jpg) |
 1f teeth.jpg) |
 2f teeth.jpg) |
 3f teeth.jpg) |
 4f teeth.jpg) |
 5f teeth.jpg) |
 6f teeth.jpg) |
 7f teeth.jpg) |
 1f.jpg) |
 2f.jpg) |
 3f.jpg) |
 4f.jpg) |
 5f.jpg) |
 6f.jpg) |
 7f.jpg) |
 8f.jpg) |
 9f.jpg) |
 90f.jpg) |
 91f.jpg) |
 92f.jpg) |
 93f.jpg) |
 1f Pteranodon.jpg) |
 1f (Tylosaurus proriger).jpg) |
 |
 |
 1f (Cretoxyrhina mantelli).jpg) |
 1f (Cretoxyrhina mantelli).jpg) |
 1f (Squalicorax falcatus).jpg) |
 82f skeleton.jpg) |
 83f skeleton.jpg) |
 84f skeleton.jpg) |
 85f skeleton.jpg) |
 86p skeleton.jpg) |
 87p skeleton.jpg) |
 88p skeleton.jpg) |
 97p skeleton.jpg) |
 94f scale.jpg) |
 80f teeth.jpg) |
 81f teeth.jpg) |
 95f teeth.jpg) |
 98f teeth.jpg) |
 99f teeth.jpg) |
 96f teeth.jpg) |
 1f (Oxyrhina retroflexus), (Oxyrhina xiphodon).jpg) |
 1f teeth.jpg) |
 2f teeth.jpg) |
 3f teeth.jpg) |
 4f teeth.jpg) |
 5f teeth.jpg) |
 6f teeth.jpg) |
 7f teeth.jpg) |
 8f teeth.jpg) |
 9f teeth.jpg) |
 90f teeth.jpg) |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
• Род † Cretodus Sokolov, 1965 (Кретодусы, Кретоды) — 112,6—66,043 млн лет назад, поздний меловой период Канада (Саскачеван), Франция, Германия, Индия, Италия, Казахстан, Швеция, Тунис, США (Алабама, Колорадо, Делавэр, Джорджия, Миннесота, Нью-Джерси, Нью-Мексико, Южная Каролина, Южная Дакота, Юта). Этот род наиболее известен по образованиям, сформированным в Западном внутреннем морском море, но также известен из позднего мела Европы, Африки и, возможно, Азии. В основном он известен по отдельным зубам и позвонкам, хотя было обнаружено несколько связанных зубных рядов и шипов. В настоящее время род Cretodus относят к предложенному монотипичному семейству † Pseudoscapanorhynchidae Herman, 1979. Шимада и Эверхарт (K. Shimada and M. J. Everhart 2019) отнесли виды Cretodus crassidens, Cretodus gigantea, Cretodus houghtonorum, Cretodus longiplicatus, Cretodus semiplicatus к этому роду, утверждая, что они являются хроновидами, отличающиеся тремя степеням ширины кронки зубов.
[Shimada, Kenshu; Everhart, Michael J. (2019-07-04). "A new large Late Cretaceous lamniform shark from North America, with comments on the taxonomy, paleoecology, and evolution of the genus Cretodus". Journal of Vertebrate Paleontology. 39 (4): e1673399.]
• † Cretodus borodini Cappetta & Case, 1975 [синоним Plicatolamna borodini Cappetta & Case, 1975] — 84,9—66,043 млн лет назад, меловой период Швеции, США (Делавэр, Джорджия, Нью-Джерси, Южная Каролина). [Cappetta, H. & Case, G.R. (1975) Contribution à l'étude des Sélaciens du groupe Monmouth (Campanien-Maestrichtien) du New Jersey. Palaeontographica, Abt. A, 151 (1–3): 1–46, 11 fig., 3 tabl., 9 pl.]
Зубы Protolamna borodini маленькие, в среднем около 6—7 мм в длину с максимальным размером чуть менее 13 мм. Этот вид имеет глубокий U-образный корень, большие одиночные треугольные выступы на каждой стороне и сильные базальные складки на губной поверхности коронки. У корня нет питательных каналов и большого язычного бугорка. Cretodus borodini можно легко спутать с более мелкой кошачьей акулой Scyliorhinus (см. фото).
• † Cretodus crassidens (Dixon, 1850) [синонимы Cretodus aff. crassidens, Cretodus cf. crassidens, Isurus crassidens, Leptostyrax crassidens, Otodus pinguis, Oxyrhina crassidens, Plicatolamna crassidens] — сеноман (100,5—93,9 млн лет назад — Сеноманский ярус) — туронское (93,9—89,8 млн лет назад — Туронский ярус) время (поздний мел). [Dixon, F. (1850) The Geology and Fossils of the Tertiary and Cretaceous Formations of Sussex. 469 p., 16 fig., 45 pl]. Вид был распространён по всему миру. До сих пор он был известен в Северной Америке только по отдельным зубам (Shimada, 2006). Первоначально вид был назван Oxyrhina crassidens Фредериком Диксоном (1850) на основании единственного зуба, найденного в графстве Западный Сассекс на юге Англии. Название рода в биномене было изменено М. Соколовым на Cretodus в 1965 году.
Приблизительно 120 зубов и 60 позвонков акулы Cretodus crassidens, были недавно собраны в сланцах Блю Хилл (Blue Hill — средний турон) около Типтона в округе Митчелл, штат Канзас, США. Образец, который сейчас хранится в Музее естественной истории Штернберга, Хейс, Канзас (FHSM VP-17575, см. фото), имеет важное значение, поскольку представляет собой первый известный достаточно полный ассоциированный материал рода Cretodus. Набор зубов включает как функциональные, так и сменные зубы, в которых самый высокий полностью развитый зуб имеет высоту коронки около 41 мм и общую высоту зуба 52,5 мм. Хотя проявляется сильная тенденция к моногнатической гетеродонтии, предполагаемый зубной паттерн относится к ламноидному типу с, по меньшей мере, 10 зубными рядами в каждом квадранте челюсти. Поскольку многие зубы имеют высокое прямое главное остриё, по крайней мере, с одной парой боковых зубцов, зубной ряд был пригоден для захвата добычи. Шимада (2006) отметил, что «зубы Cretodus crassidens большие и прочные... Каждый зуб имеет центральный бугорок с парой треугольных боковых выступов и двулопастный корень. Центральный бугорок показывает множество мелких морщинистых бороздок вдоль кромки. Боковые бугорки имеют широкое основание и треугольную форму, но по размерам намного меньше центрального бугорка. Корень массивный, а базальная вогнутость варьируется от узкой до широкой, предположительно в зависимости от положения зубов». Cretodus crassidens известен только по зубам, которые обычны в туронских морских отложениях Северной Америки. Schwimmer et al. (2002) недавно предположили, что Cretodus crassidens является конспецифическим видом Cretodus semiplicatus (Münster in Agassiz, Cappetta and Case (1999), and Cicimurri (2004)), которые отделили Cretodus crassidens от Cretodus semiplicatus. [Schwimmer, D.R., Hooks, G.E., III and Johnson, B. 2002. Revised taxonomy, age, and geographic range of the large lamniform shark Cretodus semiplicatus. Journal of Vertebrate Paleontology 22:704–707.]
В отличие от зубов, многие позвонки находились в сочленении внутри конкрементов неправильной формы, из которых был взят общий счёт позвонков на основе рентгенографического исследования с использованием компьютерного томографа. Позвонки также относятся к ламноидному типу и достигают 74 мм в диаметре. Cretodus был экстраполирован с образцами другой акулы мелового периода, Cretoxyrhina mantelli Agassiz, 1843, этот найденный экземпляр Cretodus имел общую длину от 4,2 до 5,1 м. Поскольку крупные особи Cretoxyrhina mantelli ранее считались грозными хищниками, вполне вероятно, что крупные особи Cretodus также занимали самый высокий трофический уровень в пределах их предпочтительного ареала. В то время как Cretodus crassidens и Cretoxyrhina mantelli жили одновременно, судя по летописи окаменелостей, эти два таксона, вероятно, практиковали разделение ресурсов в пределах североамериканских западных внутренних районов, потому что первый вид чаще встречается в прибрежных месторождениях, а второй — в морских.
• † Cretodus gigantea (Case, 2001) [синонимы Cretodus giganteus, Protolamna gigantea] — сеноман (100,5—93,9 млн лет назад — Сеноманский ярус), Миннесота, США. [Case, G.R. (2001) A new selachian fauna from the Coleraine Formation (Upper Cretaceous/Cenomanian) of Minnesota. Palaeontographica, Abt. A, 261(4–6): 103–112, 2 pl.] Вид описан на основании только двух довольно плохо сохранившихся зубов и диагностируется только по их большому размеру (более 5 см); поэтому этот вид считается nomen dubium. Nomen dubium («название сомнительное») — означает, что данное название имеет неопределённое применение или по причине невозможности удовлетворительной типификации, или из-за невозможности установить, к какому таксону должен быть отнесён номенклатурный тип данного названия.
• † Cretodus houghtonorum Shimada & Everhart, 2019 — 94,3—89,3 млн лет назад. Типовой образец: FHSM VP-17575, частичный скелет (разрозненный частичный скелет, состоящий не менее чем из 134 зубов, 61 позвонка и 23 плакоидных чешуек). Его местонахождение — Типтон (Tipton), недалеко от туронских морских сланцев в формации Карлайл (Carlile Formation) в Канзасе, США. [K. Shimada and M. J. Everhart. 2019. A new large Late Cretaceous lamniform shark from North America, with comments on the taxonomy, paleoecology, and evolution of the genus Cretodus. Journal of Vertebrate Paleontology 39: e1673399]
В 2010 году Кейт и Дебора Хоутоны, владельцы ранчо в штате Канзас, нашли останки ископаемого животного и передали их учёным. Ископаемая акула была раскопана в 2010 году исследователями Кеншу Шимада и Майклом Эверхартом и двумя жителями центрального Канзаса, Фредом Смитом и Гейл Пирсон. (Шимада — профессор палеобиологии в Университете ДеПола в Чикаго. Он и Эверхарт являются младшими научными сотрудниками Музея естественной истории Штернберга Государственного университета Форт-Хейса в Хейсе, штат Канзас.) Это позволило совершить открытие нового вида древних акул, который в честь первооткрывателей получил название Cretodus houghtonorum. Находка состоит из 134 зубов, 61 позвонка, 23 плацентарных чешуек и фрагментов кальцинированного хряща. Несмотря на то, что скелет сохранился не полностью и даже без головы — это лучший образец рода Cretodus в Северной Америке.
«Многое из того, что мы знаем о вымерших акулах, основано на изолированных зубах, но связанный образец, представляющий отдельную особь акулы, подобную той, которую мы описываем, предоставляет огромное количество анатомической информации, которая, в свою очередь, позволяет лучше понять её экологию», — сказал Шимада, ведущий автор исследования.
Изучая находку, палеонтологи из Университета Де Поля в Чикаго полагали, что перед ними останки уже известного вида Cretodus crassidens, первоначально описанного в Великобритании, и затем к которому отнесли многие останки из Северной Америки. Однако ни один зуб не соответствовал форме зуба оригинального образца Cretodus crassidens или любого другого известного вида Cretodus, и тогда ученые поняли, что все североамериканские находки принадлежат к другому виду, который населял эту часть планеты 91 млн. лет назад — во время позднего мелового периода, последней части эпохи динозавров. Cretodus houghtonorum, сохранившийся в отложениях, отложившихся в древнем океане, называемом Западным внутренним морским путем, который покрывал середину Северной Америки в течение позднего мелового периода (от 144 до 66 миллионов лет назад), был впечатляющей акулой, рост которой оценивался примерно в 17 футов или чуть больше (на основе нового исследования, опубликованного в Журнале палеонтологии позвоночных, длиной более 5 метров).
Cretodus houghtonorum в среднем имел не менее 5 метров в длину, и моделирование (модель роста акулы, откалиброванная по наблюдаемым кольцам роста позвонков) показало, что эта акула могла вырастать и до 6,8 метров, рождаясь с впечатляющим размером — 1,2 метра. Это позволяет предположить, что к позднему меловому периоду каннибализм среди эмбрионов акул уже развился. «Что более захватывающе, так это предполагаемый большой размер при рождении, почти 4 фута или 1,2 метра в длину, что позволяет предположить, что каннибалистическое поведение при вынашивании эмбрионов, обычно наблюдаемое в матке современных женских ламниформ, должно было развиться уже к позднему меловому периоду», — добавил Шимада.
Анализ анатомии Cretodus houghtonorum привёл к выводу, что это была довольно медлительная рыба, принадлежащая к группе ламнообразные (Lamniformes), которая включает в себя современных больших белых (Carcharodon carcharias), большеротых (Megachasma pelagios) и песчаных тигровых акул (род Carcharias).
Еще одним интересным открытием стало то, что вместе с этой окаменелостью были найдены зубы другой ископаемой акулы, принадлежащей роду Скваликоракс (Squalicorax), а также фрагменты двух плавниковых шипов вида из отряда гибодонтообразные (Hybodontiformes). Учёные предполагают, что Cretodus houghtonorum питался мелкими гибодонтами вместе со скваликораксом, либо же Squalicorax, как более крупный хищник, съел представителя новооткрытого вида после того, как эта акула пообедала гибодонтом. «Косвенно, мы думаем, что акула, возможно, питалась гораздо более мелким гибодонтом, а после смерти была съедена скваликораксом», — сказал Эверхарт.
• † Cretodus longiplicatus Werner, 1989 [синоним Semiplicatodus sulcatus] — альб (113,0—100,5 млн лет назад — Альбский ярус) — турон (93,9—89,8 млн лет назад — Туронский ярус). [Werner, C. (1989) Die Elasmobranchier-Fauna des Gebel Dist Member der Bahariya Formation (Obercenoman) der Oase Bahariya, Ägypten. Palaeo Ichthyologica, 5, 1–112]
В 2012 году в Казахстане на уровне пласта отложений, состоящего из жёлто-серых и серых глин, был обнаружен зуб морской акулы Cretodus longiplicatus. Этот вид характерен для сеномана региона Тетис (Werner 1989; Kennedy et al. 2008; Underwood и другие. 2011). По данным Соколова (1978), этот вид [= Semiplicatodus sulcatus в его употребление] широко распространён в позднеальб-сеноманских отложениях полуострова Мангышлак и западного Туркменистана, но редко встречается в туроне (93,9—89,8 млн лет назад — Туронский ярус) Казахстана.
• † Cretodus semiplicatus (Agassiz, 1843) [синонимы Lamna semiplicata (Agassiz, 1843); Otodus semiplicatus Agassiz, 1843; Oxyrhina crassidens Dixon, 1850; Cretodus aff. semiplicatus, Cretodus divaricatus, Cretodus rudis, Otodus cf. semiplicatus, Otodus divaricatus, Otodus rudis, Otodus semiplicatus] — 94,3—84,9 млн лет назад, меловой период Канада (Саскачеван), Франция, Германия, США (Алабама, Колорадо, Джорджия, Миннесота, Нью-Мексико). [Agassiz, L. (1843) Recherches sur les poissons fossiles, 15th and 16th livraisons (March 1843). Jent and Gassmann, Soleure(text) and H. Nicolet, Neuchâtel(planches). – vol. 3: [i]-[iv], 157-390, 382*-382**, 1–32, [33]-[34], pl. 1, 18, 22, 22a, 22b, 26a, 38, 40b, 40c, 40d, 45, 47]
утолщённый зуб.
У Cretodus semiplicatus большие, мощные, крепкие, утолщённые ламноидные акульи зубы, ударно-рвущего типа, с высокой короной и вертикальными или почти вертикальными коронками в переднем и передне-латеральном рядах (см. фото). Самые большие известные зубы достигают общей высоты 66 мм. Большинство зубов с парными, толстыми, сильно расходящимися треугольными боковыми выступами, образующими сплошную губную режущую поверхность с основным бугорком. Бугорки могут иметь одиночные выемки на боковых краях. Коронки в профиль слегка сигмовидные, дистально с гладкой эмелоидом. Основания коронок и зубные ленты с многочисленными, равномерно расположенными, хорошо разграниченными короткими вертикальными канавками на губной стороне всех зубов, достигающих не более 20% высоты коронки. На губной поверхности больших зубов также имеется от двух до пяти глубоких вертикальных складок. Самая длинная и глубокая борозда медиальна до середины бугорка. Лингвальная сторона с одинаковыми вертикальными бороздками на коронке и зубной ленте, достигающих 20—40% высоты коронки; язычные борозды более обширны и лучше разграничены на меньших зубах (т.е. менее 4,0 см). Корни очень крепкие, с выступающими язычными полками на всех зубах и нечёткими питательными бороздками. Корни U-образные, глубоко раздвоенные и узкие с боков, особенно в нижней челюсти.
Распространение — Африка: Ангола; Азия: Ливан, Индия; Европа: Бельгия, Франция, Англия, Германия, Литва; Северная Америка: Аризона, Нью-Мексико, Техас, Канзас, Южная Дакота, Алабама, Джорджия. Пересмотренный стратиграфический диапазон — верхний мел: сеноман (100,5—93,9 млн лет назад — Сеноманский ярус) — сантон (86,3—83,6 млн лет назад — Сантонский ярус).
 1f left teeth Scyliorhinus (right).jpg) |
 1f teeth.jpg) |
 2f teeth.jpg) |
 3f teeth.jpg) |
 4f teeth.jpg) |
 5f teeth.jpg) |
 1f teeth.jpg) |
 2f teeth.jpg) |
 3f teeth.jpg) |
 4f teeth.jpg) |
 5f teeth.jpg) |
 6f teeth.jpg) |
 1f teeth.jpg) |
 1f teeth.jpg) |
 2f teeth.jpg) |
 3f teeth.jpg) |
 |
 |
 |
• Род † Cretolamna Glikman, 1958 (Кретоламны) — 130,0—13,65 млн лет назад, меловой период Австралии, Бразилии, Конго-Браззавиль, Египта, Франции, Германии, Израиля, Японии, Иордании, Казахстана, Мадагаскара, Мексики, Марокко, Новой Зеландии, Российской Федерации, Испании, Швеции, Сирии, Украины, Соединенного Королевства, США (Алабама, Колорадо, Делавэр, Джорджия, Канзас, Мэриленд, Миннесота, Миссисипи, Монтана, Нью-Джерси, Нью-Мексико, Северная Каролина, Южная Дакота, Техас); палеоцен Дании, США (Калифорния, Миссисипи, Нью-Джерси, Южная Каролина); эоцен Египта, Пакистана, Сенегала, США (Луизиана, Миссисипи, Вирджиния), Узбекистана, Западной Сахары; олигоцен Пакистана; миоцен Франции, США (Мэриленд, Нью-Джерси).
Кретоламна была впервые описана швейцарским натуралистом Луи Агассисом с использованием пяти зубов, ранее идентифицированных как Mustelus mustelus, и собранных английским палеонтологом Гидеоном Мантеллом (Gideon Mantell) в Southerham Grey Pit около Льюиса (Lewes), Восточный Суссекс (East Sussex). В своей публикации 1835 года «Rapport sur les poissons fossiles découverts en Angleterre» Агассис повторно идентифицировал зубы как новый вид под таксоном Lamna appendiculata. В 1843 году Агассис опубликовал «Recherches sur les poissons fossiles», в который повторно исследовал пять зубов Мантелла. Используя их, а так же восемь дополнительных зубов, собранных Мантеллом, и ещё двадцать зубов, собранных различными палеонтологами в разных местах [один зуб, найденный 3-м графом Эннискилленом (3rd Earl of Enniskillen) в Speeton Clay в Йоркшире (Yorkshire); один зуб из коллекции Страсбургского музея (Strasbourg Museum) из неустановленного местонахождения; один зуб из коллекции музея Хэнкока (Hancock Museum) из Marly Chalk возле Кембриджа (Cambridge); и шесть зубов из коллекции немецкого палеонтолога Генриха Георга Бронна (Heinrich Georg Bronn) из меловых отложений в окрестностях Ахена (Aachen)], он описал вид, зубы которого имели толстые выпуклые корни, боковые выступы и были чрезвычайно разнообразны. Агассис заметил, что некоторые из исследованных зубов могут быть достаточно вариабельными, чтобы принадлежать к отдельному виду, но в конечном итоге объединили их под новым таксоном Otodus appendiculatus. Позже в 1958 году советский палеонтолог Леонид Гликман обнаружил, что этот вид принадлежит к новому роду — Cretolamna.
Несмотря на замечания Агассиса об изменчивости, его окончательная широкая интерпретация Otodus appendiculatus впоследствии привела к тому, что этот вид превратился в таксон мусорной корзины, что привело к интерпретации Cretolamna appendiculata как изменчивого космополитического вида с диапазоном жизни 50 миллионов лет. Ситуация изменилась, когда палеонтолог Микаэль Сиверссон (Mikael Siversson) обнаружил, что двадцать пять синтипов на самом деле представляют собой смесь по крайней мере шести или более различных видов, включая три дополнительных рода Dwardius, Cretoxyrhina и Cretodus. Чтобы устранить таксономический вопрос, он переименовал в 1999 году один из синтипов в качестве единственного лектотипа Cretolamna appendiculata. В 2015 году Микаэль Сиверссон возглавил исследование, которое пересмотрело таксономическую ситуацию и установило обновленное описание вида, что привело к появлению шести дополнительных видов Cretolamna — Cretolamna catoxodon, Cretolamna deschutteri, Cretolamna ewelli, Cretolamna gertericorum, Cretolamna hattini и Cretolamna sarcoporthea.
До Сиверссона были описаны другие виды Cretolamna. Другой вид, описанный Агассисом под таксоном Otodus latus, был в 1908 году понижен в звании до разновидности Cretolamna appendiculata и в 1977 году переведён в подвид французским палеонтологом Жаком Германом (Jaques Herman) и, наконец, в 2012 году повышен до уровня вида как Cretolamna lata Германом и палеонтологом Ван Ваэсом Хильде (Van Waes Hilde). В 1897 году французский палеонтолог Фернан Прием (Fernand Priem) описал единственный зуб из песчаника Кёпинге (Köpinge Sandstone) в Скании (Scania), Швеция, под таксоном Lamna borealis. Это название будет изменено на Cretolamna borealis Гликманом в статье 1980 года.
В 1902 году немецкий палеонтолог Йоханнес Ваннер (Johannes Wanner) описал зубы из египетских меловых отложений около оазиса Дахла (Dakhla Oasis) и Фарафры (Farafra). Он отметил, что зубы почти идентичны зубам Otodus appendiculatus, за исключением того, что египетские зубы также содержали две чёткие пары боковых выступов (особенность, отсутствующая у Otodus appendiculatus). Ваннер пришел к выводу, что зубы принадлежали к новому виду, тесно связанному с Otodus appendiculatus, и поместил его в таксон Otodus biauriculatus. В 1935 году французский палеонтолог Камиль Арамбург (Camille Arambourg) описал новый подвид Cretolamna biauriculata из зубов, обнаруженных в марокканских фосфатах под таксоном Lamna biauriculata maroccana, который был выделен в отдельный вид в 1997 году.
В 1972 году французский ихтиолог Анри Каппетта (Henri Cappetta) описал зубы из маастрихтских отложений недалеко от колодца Ментес (Mentès well) в Тахуа (Tahoua), Нигер, которые он отнёс к подвиду Lamna biauriculata nigeriana. Этот подвид также будет возведён в разряд своего собственного вида в 1991 году. В 1975 году Каппетта и американский палеонтолог Джерард Кейс (Gerard Case) исследовали зубы креталамны описанные Арамбургом в 1952 году из месторождений в Марокко и предложили, что они принадлежат новому подвиду типового вида и назначили таксон Cretolamna appendiculata arambourgi, который Сиверссон и др. (2015) возвели в отдельный вид. В 2018 году американские палеонтологи Джун Эберсол (Jun Ebersole) и Дана Эрет (Dana Ehret) описали новый вид Cretolamna из различных зубов формации Eutaw и Мурвильских меловых отложений в Алабаме, который они назвали Cretolamna bryanti.
Название рода Cretolamna происходит от латинского слова creta — «мел» и названия рода Lamna, который представляет собой латинизацию древнегреческого слова λαμνα (lamna) — «свирепая акула», что в совокупности означает «меловая акула» или «акула мелового периода», и относится к меловым отложениям, в которых были найдены образцы типового вида († Cretolamna appendiculata) этого рода.
Предметом разногласий было правильное написание «Cretolamna» и «Cretalamna». Первоначально Гликман описал этот род с намерением назвать его «Cretolamna», но во время публикации соответствующей статьи 1958 года произошла типографская ошибка, в которой название рода было написано с ошибкой как «Cretalamna». Гликман указал на орфографическую ошибку и продолжил использовать предполагаемое написание «Cretolamna» в более поздних работах. Это написание было повсеместно принято до 1999 года, когда Сиверссон (Siversson) заметил, что это нарушает статьи 32 и 33 МКЗН (Международный кодекс зоологической номенклатуры (ICZN) — международно признанный свод правил, касающихся употребления научных названий таксонов животных.), восстановив «Cretalamna» как допустимое написание. С тех пор восстановление названия «Cretalamna» приобрело известность и к 2010-м годам было принято большинством палеонтологов. Однако некоторые палеонтологи, в том числе Каппетта, категорически против этого. В попытке подавить использование «Cretalamna», Каппетта (Cappetta) обратился к представителю Международной комиссии по зоологической номенклатуре, утверждая, что первоначальные намерения Гликмана и преобладающее использование «Cretolamna» до Сиверссона (1999) обеспечивает его приоритет. Международная комиссия, которая, как сообщается, была впечатлена «духом» Каппетты, впоследствии создала статью 33.3.1 Кодекса 2000 года издания, чтобы разрешить эту ситуацию в будущем. В ней говорится, что «когда неоправданная поправка превалирует и приписывается первоначальному автору и дате, она считается оправданной поправкой». В то время как Каппетта утверждал в справочнике 2012 года, что это новое положение оправдывает приоритет слова «Cretolamna» из-за преобладающего использования этого написания до его замены Сиверссоном в 1999 году, сам Сиверссон указал в статье 2015 года, что это положение не может работать задним числом, и что продолжающееся преобладающее использование слова «Cretalamna» с момента введения этого положения по иронии судьбы обеспечивает его приоритет, а не угрожает ему. «Cretolamna» в настоящее время остается распространённым правописание, и палеонтологи выразили маловероятность возврата к использованию слова «Cretalamna».
Кретоламна была акулой среднего размера с предполагаемой общей длиной 2,3—3 метра. Предполагается, что у неё было веретенообразное тело, сравнимое с таковым у современных ламниформных акул. Об этом свидетельствуют округлые центры позвонков кретоламны. Учитывая размер зубов Cretolamna относительно размера её тела, вполне вероятно, что её хвостовой плавник не использовался для оглушения добычи (как в случае с лисьей акулой — семейство Alopiidae). Вместо этого хвостовой плавник кретоламны был больше похож на плавник большой белой акулы (Carcharodon carcharias).
Зубы Cretolamna отличаются широким треугольным выступом и двумя боковыми выступами. Режущие кромки зубьев похожи на бритву, а стороны имеют гладкую поверхность. Симметрия зубов переменная; некоторые имеют точную двустороннюю симметрию, тогда как другие имеют высокую асимметрию. Соседние зубы не перекрываются. Точная дентиция кретоламны является неопределённой из-за неполных ископаемых образцов. Традиционно большинство реконструкций её зубных рядов строились из отдельных зубов. Основываясь на образце Cretolamna hattini (LACM 128126), зубной ряд акулы следует ламноидному образцу с по крайней мере пятнадцатью верхними зубными рядами и восемью нижними зубными рядами с каждой стороны челюсти. Верхние зубные ряды содержат, спереди назад: два симфизиальных, два передних, один промежуточный и десять боковых зубных рядов. Нижние зубные ряды содержат: два передних, один промежуточный и пять боковых зубных рядов. Это указано в зубной формуле S2?.A2.I1.L10(+?) / s?.a2.i1.l4, построенный в 2007 году в ходе исследования LACM 128126 палеонтологом Кеншу Шимада (Kenshu Shimada). Возможно, что Cretalamna содержала более двух симфизиальных зубных рядов, поскольку родственная Cretoxyrhina mantelli имела четыре верхних симфизиальных зубных рядов.
У Cretolamna hattini верхняя и нижняя челюсти похожи на челюсти Cretoxyrhina mantelli. Челюсти также напоминают челюсти современных алопиид (акулы-молотилки — семейство Alopiidae) и ламнид (сельдевые акулы — семейство Lamnidae). Ограниченные ископаемые свидетельства предполагают, что верхняя челюсть простиралась над нижней челюстью, давая кретоламне субтерминальный рот.
Родом Cretolamna был широко распространённым, найденным в Северной Африке (Марокко), Ближнем Востоке (Иордания), Северной Америке и на Восточном побережье, и на Среднем Западе и в Центральной Америке (Тоноси, Панама). Отложения в Марокко обычно имеют эоценовый возраст; отложения в Иордании относятся к меловому и эоценовому возрасту; и большинство месторождений в США относятся к меловому и палеоценовому возрасту. Cretolamna maroccana более распространена в Марокко и Иордании, тогда как Cretolamna appendiculata более распространена в Соединенных Штатах. Оба вида в какой-то момент пересекались.
Ископаемые свидетельства кретоламны обнаружены в отложениях, представляющих разнообразный набор морской среды, что указывает на то, что эта акула была способна адаптироваться к широкому кругу сред обитания. Это могло быть связано с её способностью существовать в длительном временном диапазоне. Веретенообразное тело кретоламны предполагает, что это была пелагическая акула. Меловые воды, населенные кретоламной, также были домом для разнообразных хрящевых рыб, костистых рыб, черепах, чешуекрылых, плезиозавров, птерозавров, птиц и даже некоторых нептичьих динозавров.
Морфология зубов кретоламны предполагает, что это была универсальная акула. Это был хищник, который охотился на крупных костистых рыб, черепах, мозазавров, кальмаров и других акул. Некоторые образцы зубов Cretolamna сильно изношены — вероятно, в результате резкого изменения объектов питания.
Возможным фактором исчезновения Cretolamna является усиление конкуренции с более новыми акулами-универсалами во время кайнозоя. Широко распространено мнение, что представители рода Otodus (и, следовательно, рода Carcharocles) произошли от кретоламны из-за его сильного сходства с некоторыми видами в пределах рода.
• † Cretolamna appendiculata (Agassiz, 1843) [синонимы Galeocerdo tremauxi Sauvage, 1880; Lamna appendiculata Agassiz, 1843; Lamna appendiculatus Agassiz, 1843; Otodus appendiculatis (Agassiz, 1843); Otodus appendiculatus (Agassiz, 1843); Cretalamna aff. appendiculata; Cretalamna appendiculata; Cretalamna cf. appendiculata; Cretalamna ex. gr. appendiculata; Cretolamna aff. appendiculata; Cretolamna basalis; Cretolamna cf. appendiculata; Cretolamna levis; Lamna basalis; Lamna cf. appendiculata; Lamna lata; Lamna manitobensis; Otodus basalis; Otodus latus; Otodus levis; Otodus marginata; Protolamna manitobensis] — типовой вид — 99,7—13,65 млн лет назад, меловой период Бразилии, Конго-Браззавиль, Египта, Франции, Германии, Израиля, Японии, Иордании, Мадагаскара, Новой Зеландии, Российской Федерации, Испании, Швеции, Сирии, Украины, Соединенного Королевства, США (Алабама, Колорадо, Делавэр, Джорджия, Канзас, Мэриленд, Миннесота, Миссисипи, Монтана, Нью-Джерси, Нью-Мексико, Северная Каролина, Южная Дакота, Техас); палеоцен Дании, США (Калифорния, Нью-Джерси, Южная Каролина); эоцен США (Вирджиния); миоцен Франции, США (Мэриленд, Нью-Джерси). [Agassiz, L. (1835) Rapport sur les poissons fossiles découverts depuis la publication de la troisième livraison. In: Feuilleton additionel sur les Recherches sur les poissons fossiles. Quatrième livraison(Janvier 1835). Imprimerie de Petitpierre et Prince(text), Neuchatel, pp. 39-64]
Название типового вида appendiculata происходит от латинского слова appendiculatus — «имеющий отросток», «имеющий придаток», ссылаясь на толстые выпуклые корни зубов Cretolamna appendiculata. Местные жители Нью-Джерси называют этих акул «porbeagle» — «порбигль», окаменелые останки зубов которых являются обычной находкой в меловых ручьях этого штата. Эти зубы имеют сжатый корень, имеющий прямоугольную форму. Главный бугорок широкий, с двумя коренастыми выступами треугольной формы. Зубы Cretolamna appendiculata в среднем составляют от 1/2 до 3/4 дюйма, а некоторые зубы чуть более 1 дюйма. Эти зубы, особенно передние, могут напоминать зубы † Archaeolamna kopingensis.
Cretolamna appendiculata рассматривается как наиболее вероятный меловой предок культовой белой акулы (Carcharodon carcharias) [например, Applegate and Espinosa-Arrubarrena 1996; Purdy 1996] и / или предок группы Otodus, кульминацией которой стал Carcharocles megalodon [например, Cappetta 1987, 2012; Siverson 1992a; Zhelezko 2000; Zhelezko, Kozlov 1999; Ward 2009].
Лектотип: латерозадний зуб, предположительно верхней челюсти (местонахождение неизвестно). Типовой горизонт: сеноман (100,5—93,9 млн лет назад — Сеноманский ярус) — ранний коньяк (89,8—86,3 млн лет назад — Коньякский ярус), поздний мел. Типовая местность: район Льюиса, Восточный Суссекс, юг Англии.
Географический и стратиграфический ареал. На данный момент вид может быть точно идентифицирован только по типовой территории в Англии и раннему турону (93,9—89,8 млн лет назад — Туронский ярус) на севере Франции.
Материал. Десять зубов (WAM 13.5.1–7, UMT-BET 1–3) из Tourtia de Bettrechies (ранний турон, поздний мел); Карьер SECAB, Беттречи (Tourtia de Bettrechies), Франция.
Исправленный диагноз: боковые бугорки на верхних и задних зубах относительно небольшие и умеренно расходящиеся. Основной выступ губно (лабиально) изогнут в профиль в верхних боковых зубах (см. фото). Нижние передние зубы с высоким корнем, и умеренно высоким треугольным бугорком с лингвальной стороны (см. фото). У крупных особей корень массивный в нижних передних отделах, с выступающим лингвальным бугорком в профиль и треугольным контуром в базальном направлении. Верхние передние и передние верхние латерозадние зубы с глубоким U-образным срединным вдавлением базального края корня и округлыми лопастями. Дистальная доля корня лабиолингвально сдавлена относительно мезиальной доли в передних верхних латерозадних зубах. Лингвальный бугорок корня высокий, но апикально-базальный узкий в профиль в верхних и нижних латерозадних зубах.
Первый верхний передний зубной ряд (см. рисунок 2): зуб высотой 15,5 мм относится к первому верхнему правому ряду (рис. 2А). Отсутствует один боковой бугорок, предположительно дистальный. Основной бугорок прямой в губных / язычных проекциях и лингвально изогнут в профиле. Сохранённый бугорок симметричен и расположен под углом 45 ° к центральному бугорку. Лингвальный бугорок корня хорошо разграничен на базальном виде. Корень почти симметричен в губной / язычной проекции, хотя мезиальная доля несколько шире и лабиолингвально менее сжата.
Второй верхний передний зубной ряд: зуб высокой сохранности высотой 17 мм соответствует правой позиции А2 (рис. 2В). Крепкий центральный бугорок умеренно высокий и слегка изогнут на губно-лингвальной проекции, предположительно по направлению к комиссуре. При виде в профиль бугорок слегка изогнут в лабиальном направлении. Боковые бугорки субтреугольные и умеренно расходящиеся. Корень относительно изящный и слегка асимметричный при взгляде на губы, мезиальная доля несколько короче и шире, чем дистальная. В базальной проекции корень асимметричен, дистальная доля более сжата, чем мезиальная.
Верхние латерозадние зубные ряды: из четырёх хорошо сохранившихся верхних латерозадних зубов три (рис. 2C — E), высотой 14—15 мм, относятся к рядам, расположенным спереди (вероятно, в диапазоне LP1 — LP4), тогда как один (рис. 2F), высотой 9 мм, является расположенным сзади зубом (расчетное положение LP7 или LP8). У всех зубов режущие кромки несколько лабиально изогнуты в профиле. Бугорок слегка или умеренно наклонен дистально у передних зубов, но сильно изогнут дистально у задних зубов. Корень заметно асимметричен в двух наиболее передних зубах (рис. 2C, D), которые также имеют глубокую U-образную срединную выемку на базальном крае корня. Боковые выступы субтреугольной формы, среднего размера и относительно вертикальные на всех четырех зубах. В базальном виде корень симметричен для расположенного сзади зуба (рис. 2F3), но асимметричен для зубов, расположенных спереди, с лабиолингвально сжатой дистальной долей относительно мезиальной доли (рис. 2C1, D2, E3).
Второй нижний передний зубной ряд (см. рисунок 3): полный зуб высотой 21,5 мм от крупной акулы относится ко второму нижнему переднему ряду (рис. 3А). Треугольный бугорок немного наклонен дистально на губно-лингвальной проекции и умеренно толстый. Режущие кромки бугорка прямые при виде в профиль. Корень массивный и асимметричный на губной проекции, так как мезиальная доля длиннее и уже дистальной доли. Треугольный контур характеризует базовый вид. При виде в профиль шейка и язычный бугор резко поднимаются от лингвальной границы бугорка.
Нижние латерозадние зубные ряды: три зуба относятся к нижним боковым зубным рядам. Самый большой из трёх (вероятно, lp1), размером 18 мм в высоту, хорошо сохранился, за исключением небольшой недостающей части мезиальной доли корня (рис. 3B). Основной бугорок слегка наклонен дистально на губных / лингвальных проекциях и очень слегка лингвально изогнут в профиле. Корень заметно асимметричен с угловатой и широкой дистальной долей и более острой и узкой мезиальной долей. Лингвальный выступ относительно узкий и хорошо разграничен на виде снизу (Рис. 3B4). Срединное углубление базального края корня глубоко U-образное. У зуба среднего размера отсутствует вершина бугорка (рис. 3C3). В сохранившемся виде он имеет высоту 10 мм и должен был располагаться в средней части предполагаемой нижней латерозадней полости. Основной бугорок умеренно изогнут в дистальном направлении на губной / язычной проекции и слегка лингвально изогнут в профиль. Мезиальный бугорок заметно больше дистального. Срединное углубление базального края корня образует широкоугольную V. Самый маленький из хорошо сохранившихся нижних латерозадних зубов имеет высоту 6,5 мм (рис. 3D). Вершина дистально наклонного основного бугорка отсутствует. Скруглённое углубление посередине базального края корня широкое и глубокое. Обе доли корня субпрямоугольные в базальном виде. Узкая срединная бороздка разделяет язычный бугорок корня.
Примечания. Зубы Cretolamna appendiculata больше всего напоминают зубы ранней кампанской Cretolamna sarcoportheta, и оба вида включены в группу Cretolamna appendiculata. Предполагаемые производные признаки, объединяющие их, включают заметную асимметрию корня на базальном виде в передне (не сзади) расположенных верхних латерозадних зубах, при этом дистальная доля лабиолингвально сжата относительно мезиальной доли (рис. 2C1, D2, E3, 5B2, E1, F1), сильно сжатая дистальная доля в предполагаемых вторых верхних передних зубах (рис. 2B2, 4D1, E3, F3; обратите внимание, что бугорок кажется несколько изогнутым дистально у Cretolamna appendiculata, но слегка изогнутым у Cretolamna sarcoportheta) и очень заметный язычный выступ корня в нижних передних зубах (рис. 3A2, 7A2, B2, C21). Помимо морфологии предполагаемого A2, сопоставимые зубы кампанского вида отличаются от зубов Cretolamna appendiculata более выступающим лингвальным выступом корня (см., например, рис. 2C4, 5F3), менее вогнутой губной поверхностью на виде в профиль. В профиль верхние латерозадние зубы из сопоставимых рядов (сравните рис. 2D4 и E2 с 5F3 и H2) имеют несколько более широкий бугорок (в среднем), в передне расположенных латерозадних зубах более расходящиеся бугорки и менее сильно изогнутая срединная выемка корня.
• † Cretolamna arambourgi (Cappetta & Case, 1975) [синонимы Cretolamna appendiculata arambourgi Cappetta & Case, 1975; Cretalamna arambourgi (Cappetta & Case, 1975)] — дан (66,0—61,6 млн лет назад — Датский ярус). [Cappetta, H. & Case, G.R. (1975) Contribution à l'étude des Sélaciens du groupe Monmouth (Campanien-Maestrichtien) du New Jersey. Palaeontographica, Abt. A, 151(1–3), 1–46]
В 1975 году Каппетта (Cappetta) и американский палеонтолог Джерард Кейс (Gerard Case) исследовали зубы Cretolamna, описанные Арамбургом (Arambourg) из датских отложений в Марокко в 1952 году и первоначально обозначенные как «Lamna appendiculata», и предположили, что они представляют собой новый подвид типового вида, и присвоили ему таксон Cretolamna appendiculata arambourgi, который Siversson et al. (2015) возвёл в отдельный вид.
Название вида arambourgi дано в честь палеонтолога Камиля Арамбурга (Camille Arambourg) за открытие типовых образцов и его вклад в палеонтологию Северной Африки.
• † Cretolamna aschersoni (Stromer, 1905) [синонимы Lamna aschersoni (Stromer, 1905); Otodus aschersoni Stromer, 1905] — меловой период до эоцена. Как считается, Cretolamna aschersoni была в длину от 2 до 3 метров и обитала в Северной Африке, Северной Америке и Иордании.
Фосфатные рудники возле Хурибги (Khouribga) в Марокко являются крупнейшими в мире. Возраст фосфатных отложений колеблется от мела до эоцена. Зубы ископаемых акул чрезвычайно распространены в марокканских фосфатных отложениях (см. фото). Хотя возраст окаменелых зубов часто называют эоценом или нижним эоценом. Классификация рода Cretolamna постоянно меняется и идентификация образцов часто бывает неверной, и имеет смысл найти документ, в котором изначально был описан вид. В данном случае «Die fischreste des mittleren und oberen Eozäns von Aegypten», найти статью не так-то просто. Вся коллекция Стромера была потеряна во время бомбардировки Мюнхена Королевскими ВВС в 1944 году.
• † Cretolamna biauriculata (Wanner, 1902) [синонимы Otodus biauriculata Wanner, 1902; Cretalamna biauriculata, Cretalamna cf. biauriculata, Otodus biauriculatus] — 70,6—66,043 млн лет назад, меловой период Израиля, Иордании, Сирии, США (Северная Каролина). [Wanner, J. (1902) I. Die Fauna der obersten weissen Kreide der libyschen Wuste. Palaeontographica, 30(2), 91–152]
Название вида biauriculata происходит от латинских слов префикса bi — «дву-», «двух-» и auriculata — «ушастый», что вместе означает «имеющий два уха». Это ссылка на большие боковые выступы на зубах у этого вида, которые чем-то напоминают пару ушей (см. фото).
• † Cretolamna borealis (Priem, 1897) [синонимы Cretalamna borealis (Priem, 1897); Lamna borealis Priem, 1897] — сантон (86,3—83,6 млн лет назад — Сантонский ярус) — кампан (83,6—72,1 млн лет назад — Кампанский ярус). [Priem, M.F. (1897) Sur des dents d'élasmobranches de divers gisements sénoniens (Villedieu, Meudon, FoIx-les-Caves). Bulletin de la Société géologique de France, (Serie 3), 25, 40–56]
В 1897 году французский палеонтолог Фернан Прием (Fernand Priem) под таксоном Lamna borealis описал единственный большой верхний латерозадний зуб из песчаника Кёпинге (Köpinge) в Скании (Scania), Швеция, а также другой, ещё более крупный верхний латерозадний зуб из «мела» карьера Факсе (Faxe quarry, зона NP3), Дания. Зуб из Кёпинге обозначен как лектотип. Выбор кампанского зуба в качестве лектотипа, а не датского образца из карьера Факсе, основан на том факте, что описание Приема (1897) основано в первую очередь на кампанском образце. «Мел» в районе Кёпинге на самом деле представляет собой песчаник и имеет возраст (на основе большого количества белемнитов и аммонитов) от самого поздне-раннего кампана до средне-позднего кампана (Christensen 1986: 9). Название Lamna borealis будет изменено на «Cretolamna borealis» Гликманом в своей статье 1980 года.
Название вида borealis происходит от латинского слова borealis — «северный» и указывает на место открытия окаменелостей в ископаемых месторождениях северной Швеции.
Cretolamna borealis постоянно рассматривается российскими студентами как действительный вид на протяжении последних 30 лет. Гликман (1980) правильно сгруппировал крупные верхние латерозадние зубы (Glikman 1980) типа Cretolamna borealis из кампанских отложений в пустыне Кызылкум с высоким верхним передним зубом Cretolamna «appendiculata var. pachyrhiza» из того же месторождения. Аверьянов и Попов (1995) пречислили к Cretolamna borealis образцы из фосфатного конгломерата возле села Широкий Карамыш, примерно в 50 км к западу от Саратова, Россия. По комплексу моллюсков этот образец был отнесён к позднераннему кампанскому возрасту. При исследовании кампанских акул из Делавэра, США, Лаугинигер (Lauginiger) и Хартштейн (Hartstein) в 1983 году правильно различили зубы Cretolamna borealis (описанные ими как Cretolamna appendiculata pachyrhiza) из базальных песков Mount Laurel Sand (ранне поздний кампан в трехкратном североамериканском делении яруса) из зубов группового типа Cretolamna appendiculata (описываемого как Cretolamna appendiculata lata) из поздне-среднего кампана формации Маршаллтаун (Marshalltown Formation).
Шимада с соавторами (Shimada et al., 2010) описали изолированные выпавшие зубы Cretolamna (называемые Cretolamna appendiculata), обнаруженные в прямой связи со скелетом плезиозавра (голотип Futabasaurus suzukii) в раннесантонской (86,3—83,6 млн лет назад — Сантонский ярус) формации Тамаяма (Tamayama Formation) в центральной Японии. Рисунки с 82 выпавшими зубами указывают на присутствие более чем одного вида Cretolamna. Один неполный (кажется, что вершина бугорка отсутствует) передний зуб с тупыми выступами (Shimada et al. 2010: рис. 2.41) похож на A1 Cretolamna borealis (рис. 9A). Зубы из средней части предполагаемой латерозадней части, характерные для видов группы Cretolamna borealis (сильно изогнутый дистально основной выступ, расходящиеся бугорки и относительно глубоко выемка мезиальной вогнутости базального края корня), представлены как минимум тремя экземплярами.
Географический и стратиграфический ареал Cretolamna borealis. Сантон Японии (Хонсю, префектура Фукусима); Кампан Швеции (провинция Скания), Бельгии (провинция Эно), Франции (провинции Пикардия и Пуату-Шаранта), России (Саратовская область), Узбекистана (пустыня Кызылкум) и США (Делавэр).
Исправленный диагноз. Передние зубы удлинены и окружены парой относительно небольших, обычно округлых и сильно расходящихся бугорков. Губная поверхность бугорка почти такая же выпуклая, как и язычная поверхность большинства латерозадних зубов. Большие вертикальные складки обычно присутствуют в базальной части лабиальной стороны бугорка латерозадних зубов. На латерально-задних зубах большие бугорки, обычно, с выпуклым внутренним краем и вогнутым внешним краем. Корень прямоугольный и симметричный, с небольшим, но относительно хорошо обозначенным выступом на виде базальной части у большинства латерозадних зубов.
Описание. Первый верхний передний зубной ряд (см. рисунок 9): один зуб относится к первому верхнему правому переднему ряду (рис. 9А). В сохранившемся виде он имеет высоту 32 мм. Повреждены оба кончика долей корня, что приводит к уменьшению высоты зуба примерно на 1—2 мм. Основной выступ, как это типично для зубов этого вида, встречающихся в прибрежных слоях Фенноскандинавского щита, сильно изношен в результате манипуляций с добычей, что приводит к дальнейшему уменьшению первоначальной высоты примерно на 5—6 мм. Исходная высота, таким образом, составляет 38—40 мм. Высокий бугорок слегка изогнут дистально. Его губное основание находится на одном уровне с губной поверхностью корня (рис. 9А3). Оба боковых бугорка имеют округлую, плохо очерченную вершину и расходятся мезиально. Базальный край массивного корня сильно изогнут. Обе удлинённые доли лабиолингвально сжаты. Корень довольно симметричен на губной / язычной проекции, но асимметричен в базальной проекции, с более сжатой дистальной долей.
Второй верхний передний зубной ряд: к этому ряду относятся три зуба. Проиллюстрированы и описаны наиболее хорошо сохранившиеся. Правый зуб большего размера (рис. 9В) имеет высоту 32,5 мм, тогда как левый зуб немного меньшего размера (рис. 9С) имеет высоту 30 мм. Последний имеет несколько изношенную вершину и изначально должен был быть примерно на 1 мм выше. Левый зуб практически цел, в то время как у большего правого зуба отсутствует мезиальный выступ и часть язычного выступа корня. На виде в профиль оба зуба имеют слегка изогнутые по языку основные выступы (рис. 9B3, C2). Боковые бугорки на обоих зубах расходятся, а они несколько наклонены дистально со слегка загнутой вершиной. Корень асимметричный с более длинной мезиальной долей. Хорошо обозначенный язычный бугорок корня занимает довольно небольшую площадь в центре лингвальной поверхности корня.
Третий верхний передний зубной ряд: среди зубов из неформальной зоны Belemnellocamax mammillatus бассейна Кристианстад (Kristianstad Basin) отсутствует зубной ряд A3 с сохранённым неповреждённым корнем. Однако плохо сохранившийся зуб из участка Угнсмуннарна (Ugnsmunnarna site — WAM 13.5.20, расчётная исходная высота 30 мм) имеет относительно широкий основной выступ, загнутый на губной / язычной проекции и лабиально изогнутый в профиле (аналогично острию A3 у Cretolamna ewelli; рис. 12E). Зуб здесь не показан из-за очень плохой сохранности корня. Несколько дополнительных зубов Cretolamna borealis из Угнсмуннарны (коллекция Петера Седерстрёма — Peter Cederström) также плохо сохранились, что непонятно, учитывая, что этот вид, по-видимому, был относительно обычен на этом участке.
Верхние латерозадние зубные ряды: расположенные спереди верхние латерозадние зубы имеют треугольный основной выступ, слегка наклоненный дистально на губно-лингвальной проекции и прямой в профиль. Корень заметно асимметричен с короткой, угловатой и широкой дистальной долей и более удлинённой и острой мезиальной долей (WAM 13.5.22). Срединное углубление базального края корня V-образное. Зубы, расположенные ближе кзади, имеют дистально изогнутый бугорок (рис. 9D, E). Боковые края долей корня обычно вертикальные (рис. 9E1). Боковые выступы большие, у большинства из них выпуклая внутренняя режущая кромка.
Первый нижний передний зубной ряд (см. рисунок 10): хорошо сохранившийся зуб высотой 32,5 мм имеет черты, типичные для первого нижнего переднего ряда у современных макрофаговых акул, и поэтому соответствует этому положению зуба (рис. 10А). Основной выступ почти идеально прямой на губной / язычной проекции и довольно толстый по сравнению с остриями на двух проиллюстрированных вторых верхних передних зубах. При виде в профиль основной выступ заметно изогнут по языку. Боковые бугорки треугольные, расходящиеся, с тупой вершиной. Корень крепкий, почти симметричный, с довольно острыми лопастями. Лингвальный бугорок корня у основания широкий.
Нижние латерозадние зубные ряды: Зубы, относящиеся к предполагаемой нижней латерозадней полости, отличаются от верхних латерозадних зубов тем, что имеют более вертикальный основной выступ и более или менее лингвально изогнутый в профиле. Как и в случае с верхней челюстью, некоторые нижние латерозадние зубы имеют грубые вертикальные морщины в нижней части губной поверхности бугорка (WAM 13.5.21).
Примечания. Зубы Cretolamna borealis наиболее близки к зубам двух других видов, отнесенных к группе Cretolamna borealis: Cretolamna gertericorum и Cretolamna ewelli. Для этой видовой группы характерны удлинённые передние зубы типа A3 с бугорком, который загибается на губной / язычной проекции (рис. 12E1) и лабиально изогнут в профиль (рис. 12E2), нижние передние зубы сильно изогнуты по языку при виде в профиль (рис. 10A4, 11F2), часто прямоугольный корень в базальном виде в латерозадних зубах (рис. 9E4, 11G3) с небольшим, но хорошо разграниченным выступом (рис. 9D3, E4, 11G3, 12F4) и умеренно расширенными латерально комиссуральными зубами (Рис. 11E).
Зубы Cretolamna borealis отличаются несколькими аспектами от похожих на них зубов Cretolamna sarcoportheta. Зубы Cretolamna borealis достигают гораздо больших размеров, передние зубы примерно до 38—40 мм в высоту по сравнению с примерно 23 мм (первый верхний передний зуб из отложений бассейна Кристианстада — Kristianstad Basin) Cretolamna sarcoportheta. Передние зубы Cretolamna sarcoportheta имеют сравнительно короткий, прочный выступ и массивный корень, в то время как основное остриё передних зубов Cretolamna borealis очень высокое и относительно тонкое. Кроме этого размеры куспид становятся у Cretolamna borealis шире и корень увеличивается в толщине. Основной выступ на зубах Cretolamna borealis имеет обычно более выпуклую губную поверхность и боковые выступы часто более расходятся, чем у зубов Cretolamna sarcoportheta (см. фото). Это доведено до крайности у самых ранних кампанских Cretolamna borealis.
• † Cretolamna bryanti (Ebersole & Ehret, 2018) [синоним Cretalamna bryanti Ebersole & Ehret, 2018] — меловой период. [Jun A. Ebersole; Dana J. Ehret (2018). «A new species of Cretalamna sensu stricto (Lamniformes, Otodontidae) from the Late Cretaceous (Santonian-Campanian) of Alabama, USA»]
В 2018 году американские палеонтологи Джун Эберсол (Jun Ebersole) и Дана Эрет (Dana Ehret) провели морфометрический анализ различных зубов Cretolamna из формации Ютау (Eutaw Formation) и Мурвильского мела (Mooreville Chalk) в Алабаме (метод, основанный на сравнении соотношений различных измерений друг с другом, а не на простом распределении образцов по группам на основе их очевидной формы) и описали новый вид, который они назвали Cretolamna bryanti.
Вид назван в честь семьи Брайантов (Bryant), которая помогла улучшить репутацию Университета Алабамы (University of Alabama), Алабамского Музей естественной истории (Alabama Museum of Natural History) и Научного центра Маквэйна (McWane Science Center), благодаря их приверженности к образованию и поддержке науки.
Десятилетия сбора с обнажений верхнего мелового песчаника Tombigbee Sand Member формации Ютау и Мурвильского мела в Алабаме, США, позволили получить большое количество изолированных зубов Cretolamna (sensu stricto). Многие из этих зубов ранее относили к вымершей позднемеловой акуле Cretolamna appendiculata (Agassiz, 1843), таксону, который теперь считается в значительной степени ограниченным европейским туроном (93,9—89,8 млн лет назад — Туронский ярус). Недавние исследования пролили свет на разнообразие позднемеловых таксонов Cretolamna (ss), и здесь был открыт новый вид из Алабамы, Cretolamna bryanti. Его зубы кажутся похожими на зубы представителей группы видов Cretolamna borealis, но их можно отличить от других видов по следующим признакам: передние зубы (см. фото) с более выраженным и треугольным язычным корневым выступом, более широкий треугольный бугорок и более высокий корень относительно высоты коронки; передние латерально-задние зубы (см. фото) имеют дистально наклоненный или крючковатый главный выступ и более одной пары боковых бугорков; и латерально-передние зубы имеют сильный дистально крючковатый главный выступ и корень, который в значительной степени симметричен в базальном направлении. В настоящее время вид Cretolamna bryanti стратиграфически приурочен к сантонским (86,3—83,6 млн лет назад — Сантонский ярус) / кампанским (83,6—72,1 млн лет назад — Кампанский ярус) Dicarinella asymetrica Sigal, 1952 и Globotruncanita elevata Brotzen, 1934. Образцы планктонных фораминифер в пачке Tombigbee Sand формации Ютау и Мурвильского мела и зубы Cretolamna bryanti были собраны только в четырех округах (Даллас, Грин, Хейл и Монтгомери) центральной и западной Алабамы. Открытие Cretolamna bryanti в Алабаме дополняет наши знания о разнообразии и распространении позднемеловых отодонтид в этом регионе.
• † Cretolamna caraibaea (Leriche, 1938) [синонимы Serratolamna caraibaea (Leriche, 1938); Lamna caraibaea Leriche, 1938; Lamna caraibaea var. africana Dartevelle & Casier, 1943 (?); Serratolamna cf. caraibaea] — маастрихт (72,1—66,0 млн лет назад — Маастрихтский ярус) Марокко (Юсуфия — Youssoufia), США (Техас). [Leriche, M. (1938) Contribution à l'étude des poissons fossiles des pays riverains de la Méditerranée américaine (Venezuela, Trinité, Antilles, Mexique). Mémoires de la Société Paléontologique Suisse, 61(1), 1–42]
Некоторые учёные относят Cretolamna caraibaea к роду Serratolamna Landemaine, 1991.
• † Cretolamna catoxodon (Siverson, Lindgren, Newbrey, Cederström & Cook, 2012) [синоним Cretalamna catoxodon Siverson, Lindgren, Newbrey, Cederström & Cook, 2012] — сеноман (100,5—93,9 млн лет назад — Сеноманский ярус) Австралии. [Mikael Siversson, Johan Lindgren, Michael G. Newbrey, Peter Cederström, Todd D. Cook, 2015, «Cenomanian–Campanian (Late Cretaceous) mid-palaeolatitude sharks of Cretalamna appendiculata type. Acta Palaeontologica Polonica, 60(2), 339–384»]
В средней сеноманской части алевролита Гирл (Gearle Siltstone) в Западной Австралии обнаружены обнаружены окаменелые зубы Cretolamna catoxodon и «Cretalamna» gunsoni. Cretalamna catoxodon не может быть помещён ни в одну из трёх видовых групп, признанных в туронском периоде (93,9—89,8 млн лет назад — Туронский ярус).
Название вида catoxodon происходит от древнегреческих слов κατοξυς (katoxys) — «очень острый» и οδους (odous) — «зуб», «зубы», что вместе означает «очень острый зуб», имея в виду необычно острые режущие кромки на основном острие первого и второго нижних передних зубов этой акулы.
Диагноз. Передние зубы с тонким остриём; прямые, слегка изогнутые или умеренно наклонные в дистальном направлении. Наружный край острия более или менее вертикален на передних верхних зубах. Корень симметричен на губных / лингвальных проекциях на первых верхних и нижних передних зубах, но асимметричен на базальной проекции с одной долей более сжатой, чем другой. Лингвальный бугорок корня в верхних передних зубах плохо обозначен базально, но сильно выступает в профиль и нависает над шейкой. Большинство верхних латерозадних зубов снабжено закругленными бугорками, без отчетливой вершины.
Первый верхний передний зубной ряд (см. рисунок 20): четыре зуба относятся к первому верхнему переднему ряду. Все они схожи по размеру и общей морфологии и могут характеризоваться симметричным или почти симметричным корнем и слегка изогнутым дистально или наклонным остриём. Один из зубов почти идеально сохранился и обозначен как голотип для этого вида (рис. 20А). Он немного больше трёх других зубов и, имея высоту 24 мм, является самым большим зубом этого вида в этой коллекции. Доли корня у трёх других зубов более острые (рис. 20В3), но это, вероятно, во многом является результатом их слегка корродированной сохранности. Боковые выступы небольшие, с довольно тупой вершиной на всех четырёх зубах. У хорошо сохранившегося голотипа корень массивный с заметным язычным выступом, нависающим над шейкой (рис. 20А2).
Второй верхний передний зубной ряд: этому ряду соответствует один зуб (рис. 20D). Верхушка изношена, почти наверняка из-за манипуляций с добычей, и в сохранённом виде зуб имеет 17 мм в высоту. Остриё сильно изогнуто в дистальном направлении. Как и в зубах, относящихся к первому верхнему переднему ряду, выступы очень маленькие. Корень почти симметричен на губной / язычной проекции с немного менее сильно изогнутым базальным краем корня на губной / лингвальной проекции по сравнению с зубами, относящимися к первому верхнему переднему ряду.
Верхние латерозадние зубные ряды (см. рисунок 21): Зубы, расположенные в этой части челюсти, имеют тонкие, но лабиолингвально толстые, слегка изогнутые (рис. 21A2) основные выступы до сильно изогнутых дистально (зубы расположены ближе к задней части ряда; рис. 21G3). Боковые выступы невысокие, с необычно тупой и плохо очерченной вершиной у большинства зубов (например, рис. 21D1). Режущие кромки сигмоидальные (наиболее передние зубы; рис. 21A3) или прямые в профиле (рис. 21E2). Срединное углубление базального края корня по типу Cretolamna deschutteri, образует широкую букву V с закругленной вершиной. При виде с основания язычный бугорок широкий, но плохо отделён от остального язычного края корня (рис. 21E4).
Первый нижний передний зубной ряд: к этому ряду относится единственный зуб высотой 18 мм. Зуб имеет обесцвеченную эмальоидную ткань и подвергся некоторой коррозии корня по лингвальной границе корня и шейки и на внутренней стороне одной из долей (последнее повреждение незначительно). Помимо этих незначительных недостатков, зуб хорошо сохранился. Основной выступ симметричный, узкий. Его средняя треть мезио-дистально вогнута (рис. 20E1). Есть одна пара маленьких зубцов. Корень массивный, почти идеально симметричный и с большим язычным выступом. Конечности долей довольно острые.
Второй нижний передний зубной ряд: два зуба относятся к ряду a2. Один, высотой 22 мм, несколько корродирован (рис. 20F), тогда как другой, более крупный зуб сильно корродирован (рис. 20C). Последний имеет высоту почти 23 мм, но изначально должен был быть высотой 24—25 мм. Эти два зуба отличаются от зуба a1 асимметричным корнем и широким бугорком.
Нижние латерозадние зубные ряды (см. рисунок 22): основные выступы нижних латерозадних зубов относительно вертикальные, а бугорки обычно имеют четко выраженную вершину. Корень в зубах нижней челюсти глубже, чем в соответствующих зубах верхней челюсти (рис. 21G3, 22E2), и не расширяется в латеральном направлении за пределы латеральной границы коронки в такой же степени, как в верхних латерозадних зубах (рис. 21D1, 22B1).
• † Cretolamna deschutteri (Siverson, Lindgren, Newbrey, Cederström & Cook, 2012) [синоним Cretalamna deschutteri Siverson, Lindgren, Newbrey, Cederström & Cook, 2012] — турон (93,9—89,8 млн лет назад — Туронский ярус), Франция. [Siverson, M. & Lindgren, J. & Newbrey, M.G. & Cederström, P. & Cook, T.D. (2015) Cenomanian–Campanian (Late Cretaceous) mid-palaeolatitude sharks of Cretalamna appendiculata type. Acta Palaeontologica Polonica, 60(2), 339–384]
Вид назван в честь палеонтолога Питера Де Шуттера (Pieter De Schutter) за его усилия в поисках зубов Cretolamna из карьера SECAB, Беттречи (Tourtia de Bettrechies), север Франции, которые стали доступны Сиверссону и др. (2015), и за его работу по кайнозойским акулам из Бельгии.
Диагноз. Значительно увеличены верхние и нижние передние зубы. Большинство верхних и нижних боковых зубов с заметно сжатым остриём в лабиолингвальном направлении. Лингвальный бугорок корня относительно хорошо разграничен на базальном виде как на передних, так и на верхних зубах, но широкий и с плохо выраженным основанием у латерозадних зубов. Срединное углубление на базальном крае корня у передних зубов глубокое и U-образное, а у латерозадних зубов образует широкий и неглубокий V-образное с закругленной вершиной. Лингвальный бугорок корня передних зубов широкий, в профиль куполообразный.
Первый верхний передний зубной ряд (см. рисунок 18): неполный зуб высотой 23,5 мм соответствует позиции A1 (рис. 18A). Отсутствует мезиальный бугорок и поврежден дистальный конец обеих долей корня. Бугорок немного наклонен, предположительно в дистальном направлении. Губная поверхность зуба в профиль умеренно выпуклая. Дистальный бугорок треугольный, расходящийся и, что любопытно, с вершиной, значительно более изношенной от манипуляций с добычей, чем вершина бугорка. Основание короны образует выступ на границе короны и корня. Корень заметно асимметричен на базальном виде, мезиальная доля значительно более сжата, чем дистальная. Основание язычного бугорка хорошо очерчено на базальном виде. Базальная треть бугорка, шейка и язычный бугорок корня образуют плавно изогнутую арку при виде в профиль. Базальный край корня U-образный, мезиальная доля уже дистальной.
Верхние латерозадние зубные ряды: четыре зуба относятся к предполагаемой верхней латерозадней полости; два с левой стороны челюсти (рис. 18B, C) и два с правой стороны (рис. 18E, G). Оба верхних левых зуба, скорее всего, принадлежат передней половине предполагаемой верхней латерозадней части, а два правых зуба — задней части. Основной выступ умеренно (более передние зубы) или сильно (задние зубы) изогнут дистально на губной / лингвальной проекции. Он заметно лабиолингвально сжат и прямой (зубы расположены сзади) до лабиально изогнутого в профиле. Выступы большие и хорошо очерченные на передних зубах, но мезиальный бугорок в значительной степени сливается с мезиальным режущим краем бугорка на двух расположенных сзади зубах (Рис. 18E1, G3). Лингвальный вид корня становится все более асимметричным у зубов, расположенных ближе кзади, с увеличенной мезиальной стороной. Срединная выемка базального края широкая и неглубокая. Базальный вид у всех четырех зубов очень похож и показывает широкий, плохо очерченный выступ и слегка V-образный контур корня.
Первый нижний передний зубной ряд: один полный зуб высотой 19 мм относится к ряду a1 (рис. 18D). Основной выступ треугольный, относительно короткий и прямостоячий. В профиль он сильно изогнут в лингвальном направлении. Относительно небольшие треугольные выступы расположены по обе стороны от острия. Корень почти идеально симметричен на губной / язычной проекции и имеет закругленные лепестки на концах. Низкий, почти симметричный базальный вид показывает относительно хорошо разграниченный широкий выступ (рис. 18D2).
Второй нижний передний зубной ряд: Зуб высотой 22 мм (голотип) соответствует левой позиции a2 (рис. 18F). В зубе отсутствует небольшой участок лингвальной поверхности корня, но в остальном он имеет наилучшую сохранность поверхности корня среди всех исследованных зубов из карьера SECAB. Бугорок слегка наклонен дистально на губно-лингвальной проекции и довольно резко сужается в дистальной трети своей длины. Режущие кромки бугорка имеют лингвально изогнутый профиль и слегка сигмовидную форму. В основании куспида имеется пара расходящихся треугольных выступов. Губное основание коронки развито в виде полочки. Асимметричный корень имеет короткую, широкую и угловатую дистальную лопасть, в отличие от более удлинённой, более узкой и округлой мезиальной доли. Основание язычного бугорка хорошо очерчено на базальном виде. При виде в профиль язычный бугор небольшой.
Нижние латерозадние зубные ряды: Один зуб высотой 10,5 мм относится к предполагаемой нижней правой латерозадней части (рис. 18H). Верхушка бугорка отломана, а мезиальная доля корня имеет некоторое повреждение, в первую очередь, затрагивая верхушку доли, которая имеет более округлую форму, чем была бы изначально. Также отсутствует небольшой кусочек корня у мезиального основания язычного выступа.
Таким образом, нижние передние зубы Cretolamna deschutteri имеют в профиль лингвально изогнутый основной выступ, язычный выступ корня хорошо выражен на базальном виде (плохо выражен у Cretolamna appendiculata), но плохо отделён от основного тела корня на виде в профиль (выступает у Cretolamna appendiculata). Верхние латерозадние зубы Cretolamna deschutteri имеют симметричный корень в базальном виде (как у Cretolamna hattini, тогда как он сильно асимметричен по крайней мере в более передних зубах Cretolamna appendiculata) и лабиолингвально сжатый бугорок. Напротив, бугорок толстый и крепкий в сопоставимых верхних латерозадних зубах Cretolamna appendiculata (рис. 18E2).
Зубы Cretolamna hattini наиболее близки к таковым у Cretolamna deschutteri, и оба таксона упоминаются как виды группы Cretolamna hattini. Эта видовая группа характеризуется (дентально) большей шириной корня в базальном виде по сравнению с его высотой в нижних передних зубах, относительно хорошо разграничены, язычный бугорок корня при виде сверху как верхних, так и нижних передних зубов, широкий, но часто плохо разграниченный бугорок при виде снизу в верхних боковых зубах и широкий куполообразный выступ корня на передних зубах при виде в профиль. У других видов рода бугорок более узкий (апикобазально) и более выступающий. Передние зубы Cretolamna deschutteri заметно увеличены, тогда как передние верхние латерозадние зубы Cretolamna hattini почти такого же размера, как и верхние передние зубы (рис. 19А). Это хорошо иллюстрируется простым сравнением размеров зубов между двумя видами.
• † Cretolamna ewelli (Siverson, Lindgren, Newbrey, Cederström & Cook, 2012) [синоним Cretalamna ewelli Siverson, Lindgren, Newbrey, Cederström & Cook, 2012] — коньяк (89,8—86,3 млн лет назад — Коньякский ярус), безымянное обнажение в графстве Трего (Trego County), штат Канзас, США. [Siverson, M. & Lindgren, J. & Newbrey, M.G. & Cederström, P. & Cook, T.D. (2015) Cenomanian–Campanian (Late Cretaceous) mid-palaeolatitude sharks of Cretalamna appendiculata type. Acta Palaeontologica Polonica, 60(2), 339–384]
Зубы Cretolamna из позднеконьякской части мелового слоя Смоки-Хилл (Smoky Hill) в Канзасе были собраны с поверхности и относятся к Cretolamna.
Вид назван в честь палеонтолога Кейта Юэлла (Keith Ewell), собравшего большинство типовых образцов Cretalamna ewelli в 2004 г.
Диагноз. Верхние передние зубы с шиловидным, слегка наклонным дистально остриём или с более широкими загнутым бугорками. Бугорки от мелких до умеренно больших, на передних зубах от прямых до расходящихся. Латерозадние зубы с тонким остриём и глубоким умеренно сильно изогнутым срединным углублением базального края корня. Внутренний передний край бугорков в латерозадних зубах обычно более выпуклый, чем внешний.
Первый верхний передний зубной ряд (см. рисунок 12): зуб высотой 17,5 мм относится к положению A1 (рис. 12A). Зуб цел, за исключением изношенной вершины и небольшого повреждения от атмосферных воздействий на одной стороне бугорка. Основной выступ очень тонкий и слегка наклонен в губно-язычной проекции, предположительно в дистальном направлении. При виде в профиль он изогнут к языку. Боковые бугорки очень маленькие, расходящиеся и относительно тупые. Корень симметричен на губной / лингвальной проекции, но асимметричен на базальном, с одной долей, предположительно дистальной, более сжатой, чем другая доля (Рис. 12A3). Хотя сильно изогнутый к языку основной выступ при виде в профиль является характерной чертой нижних передних зубов группы Cretolamna borealis, хорошо разграниченный язычный выступ на базальном виде и относительно прямая граница лабиального корня / коронки указывают на то, что это зуб из верхнего переднего ряда.
Второй верхний передний зубной ряд: два зуба относятся к положению А2. Один полный и имеет высоту 20,5 мм (рис. 12C). Второй зуб значительно больше другого образца, его высота составляет 25 мм, и на нём отсутствует мезиальный выступ (рис. 12B). Он также несколько моложе, так как был взят из пласта чуть ниже MU4 (другой зуб был собран ниже MU2). Основной выступ имеет шиловидную форму, особенно у более крупного зуба, и умеренно наклонен дистально. Кончик выступа немного загнут. Боковые бугорки прямые (у самого большого зуба) или расходящиеся, от мельчайших (самый большой зуб) до умеренно больших. Бугорок несколько сигмоидально изогнут в профиле (Рис. 12B2, C3). Корень от грацильного (не массивный, небольшой, тонкий, стройный) у меньшего зуба до крепкого (большой зуб) и умеренно асимметричный на губной / язычной проекции, при этом мезиальная доля более удлинена и длиннее, чем дистальная доля.
Третий верхний передний зубной ряд: один полный зуб высотой 18 мм соответствует ряду A3 (рис. 12E). Основной выступ относительно широкий, лабиолингвально сжат (по сравнению с зубами A1 и A2) и наклонен дистально, но с заметно загнутой вершиной. На виде в профиль он сильно лабиально изогнут (рис. 12E2). Относительно небольшие выступы субтреугольные и расходящиеся. Корень асимметричный на губно-лингвальной проекции с более длинной, но более узкой мезиальной долей. Лингвальный бугорок хорошо разграничен на базальном виде. Дистальная доля более лабиолингвально сжата, чем мезиальная.
Верхние латерозадние зубные ряды: неполный зуб (обломанная мезиальная доля корня), высотой 12,5 мм, относится к средней части предполагаемой верхней латерозадней полости (рис. 12D). Основной выступ сильно изогнут в дистальном направлении и тонкий. Срединное углубление базального края корня относительно глубокое и имеет U-образную форму. Более или менее прямоугольный контур характеризует корень в базальном виде (рис. 12D2). Лингвальный бугорок небольшой, но относительно хорошо очерченный при виде с основания. Доли корня сильно лабиолингвально сжаты.
Нижние латерозадние зубные ряды: три зуба относятся к нижней латерозадней полости. Один из них, высотой 19 мм, имеет лабиолингвально сжатый, но сильно асимметричный корень, что указывает на положение lp1 (рис. 12F). Основной выступ тонкий, дистально наклонен, а бугорки расходящиеся. При виде в профиль бугорок несколько лингвально изогнут. Внутренние режущие кромки выступов выпуклые, а внешние кромки вогнутые. Лингвальный бугор очень хорошо разграничен на базальном виде (Рис. 12F4), а срединное углубление базального края глубокое и U-образное (Рис. 12F3). Другой зуб, скорее всего, принадлежит значительно более крупному представителю. Его высота составляет 18 мм, а ширина — 17,5 мм (рис. 12G). Одна из его долей корня отломана. Зуб симметричный, с прямостоячим выступом, слегка изогнутым к языку в профиль. Умеренно расходящиеся боковые выступы очень острые. Низкий и относительно плохо обозначенный язычный бугорок виден на виде снизу (Рис. 12G4). Хорошо сохранившийся зуб высотой 10 мм.
Зубы Cretolamna borealis наиболее близки к зубам двух других видов, отнесённых к группе Cretolamna borealis: Cretolamna gertericorum и Cretolamna ewelli. Для этой видовой группы характерны удлинённые передние зубы типа A3 с бугорком, который загибается на губной / язычной проекции (рис. 12E1) и лабиально изогнут в профиль (рис. 12E2), нижние передние зубы сильно изогнуты по языку в профиль, часто прямоугольный корень в базальном виде в латерозадних зубах с небольшим, но хорошо разграниченным выступом и умеренно расширенными латерально. Первые и вторые передние верхние зубы Cretolamna ewelli отличаются от таковых Cretolamna borealis своими меньшими, менее расходящимися выступами (относительный размер выступов, однако, перекрывается между двумя видами) и шиловидным остриём (рис. 12B1). У Cretolamna borealis основной выступ лабиолингвально сдавлен в первом и втором верхних передних рядах в гораздо большей степени, чем у Cretolamna ewelli. Шиловидный бугорок на первом и втором верхних передних рядах, по-видимому, не просто ювенильный признак Cretolamna ewelli, так как эта форма бугорка наиболее выражена у самого большого переднего зуба (рис. 12В).
• † Cretolamna farishi Siverson, 2010 [синоним Cretalamna farishi Siverson, 2010] — альб (113,0—100,5 млн лет назад — Альбский ярус). [Mikael U. Siversson, 2010, A New Lamniform Shark from the Upper Albian]
• † Cretolamna feldmanni (Hoganson, Erickson & Holland, 2019 [синоним Cretalamna feldmanni Hoganson, Erickson & Holland, 2019] — маастрихт (72,1—66,0 млн лет назад — Маастрихтский ярус), Формация Фокс-Хиллз (Fox Hills Formation), Северная Дакота, США. [Hoganson, J.W. & Erickson, M. & Holland, F.D. (2019) Chondrichthyan and Osteichthyan Paleofaunas from the Cretaceous (Late Maastrichtian) Fox Hills Formation of North Dakota, USA: Paleoecology, Paleogeography, and Extinction. Bulletins of American Paleontology, 398: 1-94]
Оригинальный диагноз по Hoganson et al. (2019): Вид известен только по верхнему левому боковому зубу (см. фото), характеризующегося выдающимся центральным выступом, фланкированным (окружённым) с каждой стороны одиночными хорошо развитыми бугорками, имеющими базальные продолжения режущей поверхности эмали сразу за краем корня. Весь край коронки представляет собой гладкую режущую кромку, имеющую выступы, отделённые от бугорка только глубокой бороздкой, которая почти покрыта эмалью на вершине бугорков. Вся поверхность коронки гладкая. Верхняя треть бугорка плавно сгибается в лингвальном направлении, центральной язычной выпуклости нет. Корень с незначительным центральным отверстием. Корень имеет характерную квадратную форму, образованную чёткими прямолинейными лопастями на переднем и заднем концах, причём передняя часть длиннее, от которой нижние края корня сходятся на мезиальном положении корня, образуя отчетливый прямой V-образный нижний край зуба. Зуб большой, его ширина лишь немногим больше длины.
• † Cretolamna gafsana (White, 1926) [синонимы Lamna gafsana White, 1926; Serratolamna gafsana (White, 1926)] — палеоцен, эоцен — (65,5—33,9 млн лет назад), Северная Америка, Европа и Африка (из Хурибга (Khouribga), Марокко (см. фото); Нигерия). [Errol Ivor White, (1926), Eocene Fishes from Nigeria]
Некоторые учёные относят Cretolamna gafsana к роду Serratolamna Landemaine, 1991.
Этот конкретный вид был, вероятно, трёхметровой акулой, известной по редким ископаемым зубам, что делает её одной из самых редких акул, и, вероятно, предпочитающую более открытую воду.
Необычными находками в формации Нанджемой (Nanjemoy Formation), Вирджиния (эоцен), являются большие зубы Serratolamna gafsana. В прошлом их часто неправильно идентифицировали, как Cretolamna aschersoni (Case, 1994: 114). Дэвид Уорд (David Ward), подтверждая предположение Стива Каннингема (Steve Cunningham), что они могут зубами Serratolamna gafsana, отметил, что иллюстрации у Арамбурга (1952) ясно показывают, что зубы Cretolamna aschersoni имеют неглубокий корень и «причудливые» боковые выступы. Дэвид отметил, что молодые зубы Serratolamna gafsana похожи на зубы Carcharias с множеством боковых выступов и тонкими полосами на губной стороне.
• † Cretolamna gertericorum (Siverson, Lindgren, Newbrey, Cederström & Cook, 2012) [синоним Cretalamna gertericora Siverson, Lindgren, Newbrey, Cederström & Cook, 2012] — турон (93,9—89,8 млн лет назад — Туронский ярус), Франция. [Siverson, M. & Lindgren, J. & Newbrey, M.G. & Cederström, P. & Cook, T.D. (2015) Cenomanian–Campanian (Late Cretaceous) mid-palaeolatitude sharks of Cretalamna appendiculata type. Acta Palaeontologica Polonica, 60(2), 339–384]
Видовой эпитет gertericorum образовано от имён Gert — «Герт», Eric — «Эрик» и латинского суффикса -orum (склонение во множественном числе мужского рода). Производные имена «Герт» и «Эрик» относятся к коллекционерам окаменелостей Герту Де Би (Gert De Bie) и Эрику Коллиеру (Eric Collier), оба из которых собрали большинство зубов Кретоламны из карьера SECAB, Беттречи (Tourtia de Bettrechies), исследованных Микаэлем Сиверсоном (Siversson et al., 2015).
Диагноз. Передние зубы с тонкими, удлинёнными основными выступами. Корень в нижних и верхних передних зубах имеет форму основания треугольной формы. Верхние латерозадние зубы из задней половины предполагаемой латерозадней полости с вогнутым лабиальным профилем и широким, сильно изогнутым дистально бугорком на губно-лингвальной проекции. Боковые бугорки относительно вертикальные на боковых и задних зубах, но расходящиеся на передних зубах. Срединное вдавление базального края корня небольшое и сильно изогнутое в латерозадних зубах.
Второй верхний передний зубной ряд (см. рисунок 11): Целый зуб высотой 27 мм относится к положению А2 (рис. 11А). Основной бугорок удлинён и наклонен дистально в губно-язычной проекции. Режущие кромки имеют слегка сигмовидную форму в профиле. Небольшой субтреугольный и расходящийся бугорок присутствует по обе стороны от основного выступа. Корень асимметричен на губной / язычной проекции, мезиальная доля длиннее и тоньше, чем дистальная доля. Конечности обеих долей образуют острый угол. Корень имеет довольно симметричный базальный вид с треугольным язычным выступом, хорошо отделённым от остальной части корня. Основание корня в профиль вогнутое.
Верхние латерозадние зубные ряды: четыре зуба, включённые в исследование, считаются верхними латерозадними зубами (рис. 11B — E). Скорее всего, они происходят из средней и задней части предполагаемой верхней латерозадней части. Их высота колеблется от 8 до 14 мм. Основной бугорок более или менее сильно изогнут в дистальном направлении и шире у больших зубов, чем у меньших. Губная сторона зуба вогнутая во всех зубах. Боковые бугорки относительно большие, прямые или слегка расходящиеся. Корень у всех зубов асимметричный, мезиальная доля больше, чем дистальная. Срединная выемка базального края корня довольно небольшая и сильно изогнута в двух самых больших зубах. Лингвальный бугорок очень маленький на базальном виде, но хорошо разграничен, за исключением самого большого зуба, у которого эта часть корня несколько стерта (рис. 11В3).
Первый нижний передний зубной ряд: неполный зуб соответствует позиции a1 (рис. 11F). Отсутствует один боковой бугорок и кончик основного выступа. Поверхности поломки выглядят свежими, без каких-либо признаков последующего истирания, связанного с кормлением. Это указывает на то, что зуб был поврежден при удалении из пласта. В сохраненном виде он имеет высоту 24,5 мм. Первоначальная высота оценивается в 27—29 мм. Узкий основной бугорок немного наклонен на губной / язычной проекции и лабиально изогнут в профиле. Основание коронки образует довольно острый угол кнутри на губной стороне. Оставшийся боковой бугорок по морфологии почти идентичен таковому у зуба A2, демонстрируя субтреугольную форму со слегка выпуклыми режущими краями. Корень почти идеально симметричен как в губно-лингвальной, так и в базальной проекции. Как и в случае a1 Cretolamna borealis (рис. рисунок 10A), и в отличие от верхних передних зубов, язычный бугор не разграничен на базальном виде (рис. 11F4). Базальный край корня V-образный.
Нижние латерозадние зубные ряды: одиночный зуб высотой 11 мм относится к предполагаемой нижней латерозадней полости (рис. 11G; голотип). Основной бугорок треугольный, умеренно наклонен дистально. Губная поверхность бугорка морщинистая посередине в базальной части (общая черта у видов группы Cretolamna borealis). Субтреугольные, расходящиеся бугорки с выпуклыми краями расположены по обе стороны от основного выступа. Губная сторона зуба в профиль очень слабо вогнута. Корень глубокий и сильно асимметричный при лингвальном / губном взгляде, при этом мезиальная доля намного больше, чем дистальная доля. Наружные края корня прямые и вертикальные, а срединное углубление базального края небольшое и U-образное.
Зубы Cretolamna gertericorum наиболее близки к таковым у Cretolamna borealis и Cretolamna ewelli. Различия между первыми двумя видами изложены выше при описании Cretolamna borealis. Приписанный к A2 Cretolamna gertericorum (Рис. 11A) отличается от соответствующих зубов Cretolamna ewelli (Рис. 12B, 12C) его значительно более лабиолингвально сжатым остриём. Верхние латерозадние зубы отличаются от аналогичных по размеру зубов Cretolamna ewelli их более широким основным бугорком (рис. 11C3, 12D3), более вогнутым лабиальным профилем и более узким, более сильно изогнутым срединным углублением базального края корня.
• † Cretolamna hattini (Siverson, Lindgren, Newbrey, Cederström & Cook, 2012) [синоним Cretalamna hattini Siverson, Lindgren, Newbrey, Cederström & Cook, 2012] — сантон (86,3—83,6 млн лет назад — Сантонский ярус) — кампан (83,6—72,1 млн лет назад — Кампанский ярус). Географический и стратиграфический диапазон: раннесантонская формация Тамаяма (amayama Formation), Япония, и поздний сантон или самая ранняя кампанская часть мела Смоки-Хилл (Smoky Hill), Канзас, США. [Siverson, M. & Lindgren, J. & Newbrey, M.G. & Cederström, P. & Cook, T.D. (2015) Cenomanian–Campanian (Late Cretaceous) mid-palaeolatitude sharks of Cretalamna appendiculata type. Acta Palaeontologica Polonica, 60(2), 339–384]
Вид назван в честь покойного геолога Дональда Э. Хаттина (Donald E. Hattin) «за его работу по стратиграфии формации Ниобрара (Niobrara Formation), западный Канзас».
Голотип: LACM 128126, частичный скелет, состоящий из нёбно-квадратичных отростков (palatoquadrates), меккелевских хрящей (Meckel's cartilages), 35 позвонков и около 120, в основном разрозненных, зубов. Поздний сантон или ранний кампан, поздний мел, округ Логан, штат Канзас, США.
«Меккелевский хрящ», представляет собой кусок хряща, из которого произошли нижние челюсти позвоночных. Первоначально это был нижний из двух хрящей, который поддерживал первую жаберную дугу у ранних рыб. Затем он стал длиннее и сильнее и приобрел мышцы, способные закрывать развивающуюся челюсть. У ранних рыб и у хондрихтианов (хрящевых рыб, таких как акулы) меккелевский хрящ продолжал оставаться основным компонентом нижней челюсти. Но у взрослых форм остеихтианов (костистых рыб) и их потомков (земноводных, рептилий (включая птиц), млекопитающих) хрящ был покрыт костью, хотя у их эмбрионов челюсть изначально развивалась как меккелевский хрящ. У всех четвероногих хрящ частично окостеневает (превращается в кость) на заднем конце челюсти и становится суставной костью, которая является частью челюстного сустава у всех четвероногих, кроме млекопитающих. У некоторых вымерших групп млекопитающих, таких как эвтриконодонты, хрящ Меккеля все ещё соединен с челюстью, в остальном, это совершенно современные ушные кости.
У Cretolamna hattini верхняя и нижняя челюсти похожи на челюсти Cretoxyrhina mantelli. Челюсти также напоминают челюсти современных алопиид (акулы-молотилки — семейство Alopiidae) и ламнид (семейство Lamnidae). Ограниченные ископаемые свидетельства предполагают, что верхняя челюсть простиралась над нижней челюстью, давая кретоламне Хаттина субтерминальный рот.
Диагноз. Квадратный отросток более чем в два раза дорсовентрально выше самой узкой части нёбно-квадратичного отростка, кпереди от квадратного отростка, но кзади от области, в которой находятся ряды передних зубов. Отсутствие уменьшения размера зубов через переднюю / латерозаднюю границу верхней челюсти. Градационное изменение морфологии корня в базальной проекции от вторых верхних передних зубов к боковым задним зубам. По всем аспектам морфология второго зуба верхней челюсти ближе к таковым соседних верхних латерозадних зубов, чем к первому верхнему переднему зубу. В зубном ряду нет зубов с загнутым бугорком. Апикальные две трети мезиального переднего края бугорка во всех зубах заметно выпуклые. Центры позвонков с умеренно прочными, плотно упакованными радиальными пластинками, иногда раздваивающимися дистально, рядом с известковым телом. Круглые ламели отсутствуют. Отверстия нервной дуги умеренно узкие, шиловидные, проходят на всю длину центра. Толщина известкового тела составляет 1/7 длины центра.
Описание. См. Шимада (Shimada, 2007) — описание нёбно-квадратичных отростков, меккелевского хряща и зубов. Его описание разъединённых зубов, относящихся к LACM 128126, было основано в основном на неоптимально подготовленном материале. Последующая подготовка позволила получить важную дополнительную информацию о морфологии корня в базовом виде.
Позвоночный столб (см. рисунок 14): Сохранившаяся часть позвоночного столба состоит из 35 позвонков среднего ствола (обозначенных Shimada 2007 от «v1» до «v35») размером от 38 до 48 мм в диаметре. Ширина позвонков примерно в 2,5 раза больше, чем их длина (v30 sensu Shimada 2007: рис. 1D, ширина 45 мм и длина 19 мм). Как указывает Шимада (2007), многие позвонки сильно деформированы, что не позволяет проводить много значимых измерений. Дорсальные отверстия выпуклые с очень закруглёнными углами и примыкают к переднему и заднему известковому телу (рис. 14В). Межфораминальная ширина нервной дуги 10—13 мм, внутрифораминальная ширина 7—8 мм. Вентральные отверстия проходят на всю длину позвонков, упираясь в каждое известковое тело (рис. 14C), и являются узкими (4,25 мм; v11 и v32 у Shimada, 2007) относительно своей длины (14 мм; v10); хотя большинство из них сильно искажены. Область вокруг вентрального отверстия сильно укреплена кальцинированным хрящом, что придает ему гладкий и выпуклый вид (рис. 14С). Позвонки округлые, с прямыми умеренно прочными радиальными пластинками, которые могут раздваиваться рядом с известковым телом. Радиальные ламели имеют толщину 0,9—1,2 мм и обычно расположены на равном расстоянии 1,2—1,7 мм друг от друга. Концентрических ламелей нет. Кайма известкового тела узкая (рис. 14С). Известковое тело всегда тонкое (v8 в Shimada 2007: рис. 1) и не сильно утолщается к краю. На латеральном крае толщина известкового тела составляет примерно 1/14 диаметра позвонка. Толщина известкового тела составляет примерно 1/7 длины позвонка. Рядом с известковым телом отсутствуют поры или радиальные каналы.
Зубы Cretolamna hattini наиболее близки к таковым Cretolamna deschutteri (см. рисунок 18), и оба таксона упоминаются как виды группы Cretolamna hattini. Эта видовая группа характеризуется (дентально) большей шириной корня в базальном виде по сравнению с его высотой в нижних передних зубах (сравните рис. 17A2, 18D2 с рис. 7A3, 10A2, 11F4; все вероятны a1), язычный выступ корня на виде сверху как верхних, так и нижних передних зубов, широкий, но часто плохо разграниченный выступ на базальном виде в верхних латерозадних зубах и широкий куполообразный выступ корня на передних зубах при виде в профиль. У других видов рода бугорок более узкий (апикобазально) и более выступающий. Передние зубы Cretolamna заметно увеличены, тогда как передние верхние латерозадние зубы Cretolamna hattini почти такого же размера, как и верхние передние зубы. Это хорошо иллюстрируется простым сравнением размеров зубов между двумя видами. Самые большие передние зубы Cretolamna deschutteri из Беттречи (Tourtia de Bettrechies) немного больше, чем эквивалентные передние зубы LACM 128126. Напротив, передние латерозадние зубы у Cretolamna deschutteri значительно меньше, чем соответствующие зубы LACM 128126. Хотя лингвальный и базальный виды корня одинаковы у латерозадних зубов этих двух видов, бугорок более узкий у Cretolamna deschutteri лабиолингвально намного тоньше, чем у Cretolamna hattini.
Шимада (2007: рис. 6C) проиллюстрировал правый нёбно-квадратичный отросток LACM 128126, что, по его мнению, представляло первые семь зубов верхней челюсти (с использованием разрозненных зубов LACM 128126 и исключая парасимфизы) и отнесённых им к первому и второму переднему зубному ряду (A1 – A2), промежуточный ряд (I1 = самый дистальный верхний передний ряд, Sensu Siverson 1999) и первые четыре зуба в верхних латерозадних полостях (обозначены как L1–4 по Shimada 2007). Все несартикулированные зубы у Шимады (2007: рис. 6С), за исключением зуба, назначенного в положение А1, были связаны с частично обнажёнными бугорками на месте замещающих зубов на нёбно-квадратичном отростке. Разъединённый зуб, рассматриваемый Шимадой (2007) как верхний промежуточный зуб (I1), меньше по размеру по сравнению с соседними зубами в реконструкции Шимады (2007: рис. 7B) и имеет сильно изогнутый дистально бугорок. Как показано на рис. 13C – E, существует небольшое сходство в форме бугорка между зубом на месте, обозначенным Шимадой как верхний «промежуточный» (2007: рис. 6C), и зубом с разнесёнными боковыми выступами, отнесённым им к тому же ряду (Рис. 13C).
Отдельные зубы, отнесённые к верхнему «промежуточному» зубному ряду Шимадой (2007), не показывают каких-либо специализированных особенностей корня, которые характеризуют верхние «промежуточные» зубные ряды современных ламнид (семейство Lamnidae). Вместо этого базальная поверхность корня имеет ничем не примечательные очертания (рис. 16A1, B1), очень похожие на контуры других зубов верхней челюсти из средней части предполагаемой латерозадней области (рис. 15H1, 16C1). У ламнид (Lamnidae) дистальная доля корня, обращённая к промежуточной перемычке, обычно лабиолингвально сдавлена и может быть лингвально изогнута при взгляде снизу.
Shimada (2007: 586) указал на наличие до 15 углублений на медиальной поверхности правого нёбно-квадратичного отростка, каждая из которых потенциально соответствует отдельному ряду зуба (хотя он назначил 13 зубных рядов в верхней челюсти, за исключением парасимфизов, LACM 128126 в Shimada 2007 : рис. 5C). В реконструкции зубного ряда LACM 128126 Сиверсон и его коллеги назначили 12 зубных рядов на верхней челюсти (в дополнение, по крайней мере, к одному ряду парасимфизов) и 10 зубных рядов на нижней челюсть. Некоторые из зубов, отнесённых к верхней челюсти Shimada (2007), вероятно, являются зубами нижней челюсти, на что указывают их более узкие, более вертикальные бугорки и прямой или выпуклый профиль лабиальной стороны коронки.
• † Cretolamna lata (Agassiz, 1843) [синоним Otodus latus Agassiz, 1843; Lamna lata Agassiz, 1843; Cretolamna lata (Herman, 1977); Cretolamna appendiculata lata (Herman, 1977); Lamna appendiculata lata Herman, 1977] — кампан (83,6—72,1 млн лет назад — Кампанский ярус) — маастрихт (72,1—66,0 млн лет назад — Маастрихтский ярус). [L. Agassiz. 1843. Recherches Sur Les Poissons Fossiles. Tome III (livr. 15-16). Imprimérie de Petitpierre, Neuchatel]
Этот вид был описан Агассисом под таксоном Otodus latus, но был понижен в звании до разновидности Cretolamna appendiculata в 1908 году и переведён в подвид в 1977 году французским палеонтологом Жаком Германом. [Herman, J. (1977) Les Sélaciens des terrains néocrétacés et paléocènes de Belgique et des contrées limitrophes. Eléments d'une biostratigraphie intercontinentale. Mémoires pour servir à l'explication des Cartes géologiques et minières de la Belgique, 15: 401 p., 25 fig., 21 pl.]
Повышен до уровня вида как Cretolamna lata Германом и палеонтологом Ван Вайсом Хильде в 2012 году. [Jaques Herman; Van Waes Hilde (2012). Observations concernant l'Évolution et la Systématique de quelques Euselachii, Neoselachii et Batoidei (Pisces - Elasmobranchii), actuels et fossiles (PDF) (in French). Géominpal Belgica 2. pp. 55–56.],
Название вида lata происходит от женской формы латинского слова latus — «широкий», ссылаясь на заметно широкие зубы у этого вида (см. фото).
Otodus latus (Герман, 1977) Агассис (1843 г.) явно представляет собой особый маастрихтский вид. Однако его необходимо определять на основе однородного материала, собранного в узком стратиграфическом интервале на северо-западе Европы. Его зубы в абсолютном выражении больше, а в относительном — шире, чем у последних раннекампанских Cretolamna sarcoportheta. Бугорки ниже и в среднем меньше у маастрихтских видов, и у них может отсутствовать четко выраженная вершина (Герман 1977: табл. 9: 4c). У Cretolamna sarcoportheta мезиальная и дистальная доли корня латерозадних зубов разделены довольно заметной срединной выемкой, тогда как срединное углубление у Cretolamna lata невысокое. Хотя вероятно, что Cretolamna lata принадлежит к группе Cretolamna appendiculata, она нуждается в доработке.
Передние зубы среднего размера, обычно 20 мм высотой и симметричной формой. Они состоят из
приподнятого центрального выступа треугольной формы с аналогичным расходящимися бугорками с обеих сторон. Верхние зубы более сжаты с боков, чем нижние, и имеют более широкие дополнительные бугорки. Лингвальная грань коронки выпуклая, с почти плоской губной стороной. Корень приподнят в латеральном аспекте на язычной поверхности, почти плоский на губной. Корень не имеет медиальной бороздки на
язычной стороне.
Зубы позднего кампана / раннего маастрихта из Нью-Джерси, США, представленные как Cretolamna appendiculata lata Каппеттой и Кейсом (1975: табл. 3: 10–27), близки по морфологии и типу зубов к Cretolamna sarcoportheta. Ввиду отсутствия тщательного пересмотра дентально сходных Cretolamna lata и точных стратиграфических данных для материала Нью-Джерси, вероятно, будет непродуктивно уточнять дальнейшую идентификацию. То же самое относится и к подавляющему большинству предыдущих записей о Cretalamna типа Cretolamna appendiculata, поскольку в этих отчетах неизменно отсутствуют зубы с нетронутой сохранностью корня, показанные на базальных изображениях и изображениях в профиль (наиболее диагностические изображения в этой группе акул, если корень остается неизменным).
Зубы Cretolamna appendiculata lata обладают рядом характеристик, которые отличают его от других видов. Их зубы имеют очень широкие и крепкие корни, которые имеют почти прямоугольную форму и перпендикулярны главному бугорку. Корень также сдавлен и, если смотреть сбоку, не имеет заметного медиального выступа. Треугольные и гладкие главные выступы сопровождаются двумя большими коренастыми выступами треугольной формы. Боковые и задние зубы похожи на передние, за исключением того, что главный бугорок уменьшен с небольшим изгибом к задней части челюсти. Их зубы обычно имеют длину от 1 до 2 см.
• † Cretolamna maroccana (Arambourg, 1935) [синонимы Lamna biauriculata maroccana Arambourg, 1935; Cretolamna biauriculata var. maroccana (Arambourg, 1935); Serratolamna (Cretalamna) maroccana (Arambourg, 1935)] — мел — эоцен (145,5—33,9 млн лет назад), Северная Африка, Ближний Восток, Северная Америка. [Arambourg, C. (1935) Note préliminaire sur les vertébrés fossiles des phosphates du Maroc. Bulletin de la Société géologique de France, 5(5), 413–439]
В 1935 году французский палеонтолог Камилла Арамбург (Camille Arambourg) описал новый подвид Cretolamna biauriculata из зубов, обнаруженных в марокканских фосфатах под таксоном Lamna biauriculata maroccana, который Каппетта выделил в отдельный вид в 1997 году. [Gerard Case; Henri Cappetta (1997). «A new selachian fauna from the Late Maastrichtian of Texas (Upper Cretaceous/Navarro Group; Kemp Formation)». Münchner Geowissenschaftliche Abhandlungen Reihe A: Geologie und Paläontologie. 34 (1): 134–189.]
Название вида maroccana — это женская форма латинского слова «maroccanus» (марокканский), отсылка к местности в Марокко, где был найден типовой образец.
Cretolamna maroccana (см. рисунок) была широко распространена в Марокко (месторождения в Марокко обычно имеют эоценовый возраст) и Иордании (отложения в Иордании относятся к меловому и эоценовому возрасту). Реже, чем Cretolamna appendiculata, зубы Cretolamna maroccana находят в позднемеловых отложениях от Западной Атлантики и побережья Мексиканского залива до Техаса. Их можно отличить по двойным боковым выступам и слабовогнутому краю базального корня (см. фото).
• † Cretolamna nigeriana (Cappetta, 1972) [синоним Lamna appendiculata var. nigeriana Cappetta, 1972] — маастрихт (72,1—66,0 млн лет назад — Маастрихтский ярус). [Cappetta, H. (1972) Les poissons crétacés et tertiaires du bassin des Iullemmeden (République du Niger). Palaeovertebrata, 5(5), 179–251]
В 1972 году французский ихтиолог Анри Каппетта описал зубы из маастрихтских отложений недалеко от колодца Ментес (Mentès well) в административном центре Тахуа (Tahoua), Нигер, которые он отнёс к подвиду Lamna biauriculata nigeriana. Этот подвид будет возведён в категорию собственных видов в 1991 году. [R.T.J. Moody; P.J.C. Sutcliffe (1991). "The Cretaceous deposits of the Iullemmeden Basin of Niger, central West Africa". Cretaceous Research. 12 (2): 137–157.]
Название вида nigeriana происходит от названия страны Niger с префиксом -iana, женского варианта латинского суффикса -anus — «относящийся к», что вместе означает «относящийся к Нигеру» и ссылается на место, где был найден типовой образец.
• † Cretolamna pachyrhiza Herman, 1977 [синоним Cretolamna appendiculata pachyrhiza Herman, 1977] — маастрихт (72,1—66,0 млн лет назад — Маастрихтский ярус), палеоцен. [Herman, J. (1977) Les Sélaciens des terrains néocrétacés et paléocènes de Belgique et des contrées limitrophes. Eléments d'une biostratigraphie intercontinentale. Mémoires pour servir à l'explication des Cartes géologiques et minières de la Belgique, 15: 401 p., 25 fig., 21 pl.]
Жак Герман (Jaques Herman) в 1973 году описал зуб из Монтиана (Montian), (нижний палеоцен), Европа. Cretolamna appendiculata pachyrhiza имеет большой ареал, который включает Европу (Монтиан), Африку (Марокко), Азию и Северную Америку (Формация горы Лорел — Mount Laurel Formation, Нью-Джерси). На ферме Биггс (Biggs Farm), Делавэр, Cretolamna pachyrhiza заменяет вид Cretolamna lata. Это считается первым сообщением о появлении окаменелостей Cretolamna pachyrhiza на Атлантическом побережье.
Зубы большие, обычно наклонные, высота 25 мм (см. фото). Передние зубы прямостоячие, симметричные, состоящие из приподнятого, выпуклого в лингвальном направлении, треугольного центрального бугорка без бороздок и зазубрин, и сильно расходящихся треугольных бугорков с обеих сторон. Двудольный корень крепкий, с выступающей срединной бобышкой без бороздок. В корневые доли урезаны на концах. Боковые зубы более сжатые с боков и наклонены кзади. Боковые зубчики по сторонам от центрального бугорка широкие, треугольные и сильно расходящиеся. Корень умеренно выпуклый лингвально и усечён на концах. Зубы Cretolamna pachyrhiza отличаются от зубов вида Cretolamna lata более крупными размерами, более крепкими строением и сильно отклоняющимися боковыми куспидами. В общем виде он внешне напоминает Lamna obliqua палеоцена / эоцена.
• † Cretolamna sarcoportheta (Siverson, Lindgren, Newbrey, Cederstrom & Cook, 2012) [синоним Cretalamna sarcoportheta Siverson, Lindgren, Newbrey, Cederstrom & Cook, 2012] — кампан (83,6—72,1 млн лет назад — Кампанский ярус). [Siverson, M. & Lindgren, J. & Newbrey, M.G. & Cederström, P. & Cook, T.D. (2015) Cenomanian–Campanian (Late Cretaceous) mid-palaeolatitude sharks of Cretalamna appendiculata type. Acta Palaeontologica Polonica, 60(2), 339–384]
Название вида sarcoportheta происходит от древнегреческих слов σαρκος (sarkos) — «плоть» и πορθητης (porthitis) — «победитель», «завоеватель», «разрушитель», что вместе означает «победитель плоти» или «разрушитель плоти», имея в виду необычно мощные зубы этой акулы.
Голотип: Второй верхний левый передний зуб, LO 10888T (рис. 4D). Паратипы: LO 6347t, 6348t, 6351t, 10889t — 10920t (рис. 4A — C, E, F, 5A — H, 6A — E, 7A — C, 8A — J, 19C: lp7; Siverson 1992a: pl. 1: 1—4, 11). Типовой горизонт: Неизвестный горизонт в толще слабоконсолидированных известковистых песков мощностью до 2 м последнего раннего кампана (неофициальная зона белемнитов Belemnellocamax mammillatus), поздний мел. Типовая местность: Осен (Åsen), котлован Кристианстад (Kristianstad Basin), юг Швеции. Дополнительный материал — 216 зубов из неформальной зоны Belemnellocamax mammillatus (поздний ранний кампан), местонахождение Осен, котлован Кристианстад, южная Швеция (LO 6352t, 6354t и 10921t — 11134t).
Географический и стратиграфический ареал. Кампан Швеции (провинция Скания — Scania Province), Бельгии (провинция Эно — Hainaut Province) и Франции (карьер Боваль — Beauval quarry, провинция Пикардия — Picardie Province; коллекция Питера де Шуттера — Pieter De Schutter).
Диагноз. Крепкие передние зубы с толстым треугольным основным бугорком средней высоты; либо прямые, либо несколько наклоненные дистально, либо слегка изогнутые / загнутые назад. У передних зубов корень асимметрично субтреугольный в основании, одна доля значительно шире другой. Основной выступ очень широкий, треугольной формы в передних боковых зубах с плоской губной стороной бугорка в профиль. Основной выступ более узкий, обычно более вертикальный у латерозадних зубов со слегка лингвально изогнутым при виде в профиль. Корень заметно асимметричный на базальном виде у расположенных впереди верхних латерозадних зубов, при этом дистальная доля уже мезиальной доли во всех типах зубов, кроме одного. Последний, отчетливый тип зуба с апикально изогнутым основным бугорком и субпрямоугольным корнем в базальном виде, с корнем-лопастью на той же стороне вертикальной средней линии зуба, и вершина бугорка, более сжата, чем остальная доля.
Парасимфизные зубы: Из местностей Осен (Åsen ) и Игнаберга (Ignaberga) нет полностью парасимфизных зубов, поскольку корень корродирован в различной степени во всех образцах. Основной бугорок толстый, треугольной формы и поставлен под углом (Siverson 1992a: табл. 1: 16). Некоторые зубы имеют по крайней мере один маленький треугольный выступ (Siverson 1992a: pl. 1: 13; рис. 19C). Корень асимметричный при виде с основания, одна сторона тоньше другой. Более тонкая доля корня находится на той же вертикальной стороне зуба на лингвальной / губной проекции, что и вершина наклонного бугорка (Siverson 1992a: рис. 12).
Первый верхний передний зубной ряд (см. рисунок 4): к этому зубному ряду относятся три полных зуба (в дополнение к нескольким неполным образцам). Их высота составляет 17,5, 18,5 и 19 мм. Основной бугорок лабиолингвально очень толстый, даже больше, чем у первого нижнего переднего зуба. Бугорок прямой (Рис. 4B1) или умеренно изогнутый дистально (Рис. 4A1). Боковые бугорки маленькие на всех первых верхних передних зубах и расходятся до субпараллельности вертикальной оси бугорка. Шейка довольно узкая, равномерной апико-базальной ширины по всей границе корень-коронка. Корень симметричен в губной / язычной проекции, но различается как по общей ширине, так и по ширине долей. Дистальная доля корня лабиолингвально сжата относительно предполагаемой мезиальной доли (рис. 4A3, B2, C1).
Второй верхний передний зубной ряд: к этому зубному ряду относятся три полных зуба (в дополнение к нескольким неполным образцам), размером 16,5—17 мм в высоту. Основной бугорок треугольный, лабиолингвально умеренно сжатый, прямой (рис. 4D3) или несколько дистально наклонен (рис. 4F2) со слегка загнутой вершиной. Боковые бугорки маленькие и более или менее расходящиеся. Корень заметно асимметричен с широкой мезиальной долей и более узкой, более короткой и менее расходящейся дистальной долей. На губной и лингвальной проекциях базальный край мезиальной доли прямой или слегка выпуклый, тогда как базальный край дистальной доли прямой или слегка вогнутый. «Отношение максимальной ширины корня к высоте зуба» у вторых верхних передних зубов больше, чем у первых передних верхних зубов, так как первые в среднем немного короче, но в то же время несколько шире.
Верхний латерозадний зубной ряд (см. рисунок 5): Первый верхний латерозадний зуб имеет отличительную морфологию у этого вида. Основной бугорок вертикальный в своей базовой половине, но имеет слегка или умеренно изогнутую дистально вершину (рис. 5A1). Корень асимметричный, с субвертикальной закруглённой мезиальной долей и более широкой и угловатой дистальной долей. Мезиальная доля заметно сдавлена лабиолингвально (рис. 5А2), предположительно в результате положения рядом с промежуточной перемычкой. Корень также асимметричен во втором верхнем латерозаднем зубе, с субчетырехугольной дистальной долей и более острой мезиальной долей. Предполагаемые вторые верхние латерозадние зубы имеют прямой или слегка вогнутый профиль (рис. 5B4, C2). После реконструкции (рис. 19С) корень становится все более симметричным, продвигаясь кзади через ряды верхней челюсти, и достигает почти идеальной симметрии в средней части латерозаднего ряда (рис. 5G3). Кзади, за этой точкой, корень становится все более асимметричным, снова показывая увеличенную мезиальную долю (Рис. 6D1). Основной бугорок слегка наклонен дистально в мезиальной половине рядов и изогнут дистально, а не наклонен в большинстве зубов в задней половине латеропадних рядов. На виде сверху дистальная доля корня обычно сдавлена по сравнению с мезиальной долей у расположенных впереди верхних латерозадних зубов (рис. 5B2, E1, F1).
Первый нижний передний зубной ряд (см. рисунок 7): почти идеально сохранившийся зуб вместе с неполным образцом, в дополнение к нескольким менее хорошо сохранившимся зубам, относятся к первому нижнему переднему ряду зуба (рис. 7A, B). Зуб хорошей сохранности — симметричный зуб высотой почти 19 мм. Плотно изогнутый базальный край корня и симметрия образца указывают на то, что он происходит из первого переднего ряда. Один из его боковых выступов рудиментарный, а другой маленький, но хорошо сформированный. Зуб отнесён к первому нижнему, а не первому верхнему переднему ряду на основании его массивного корня с большим язычным выступом и несколько лингвально изогнутым бугорком, хотя разница в кривизне бугорка на виде профиля между предполагаемыми верхним и нижним передние зубы у этого вида относительно невелики. Насколько можно судить по его неполной сохранности, другой зуб, относящийся к этой позиции, очень похож на зуб целиком.
Второй нижний передний зубной ряд: к этому ряду относится большой неполный зуб (в дополнение к нескольким другим, еще более неполным зубам) (рис. 7C). В сохранившемся виде зуб имеет высоту чуть более 21 мм и первоначально должен был быть около 22 мм. Дистальная часть мезиальной доли корня неполная, но в остальном зуб хорошо сохранился. Боковые бугорки расходящиеся и больше, чем у других передних зубов. Массивный бугорок дистально изогнут, сохранившаяся дистальная доля корня имеет прямоугольную форму. Шейка относительно узкая, как и у других передних зубов этого вида. Базальный край корня V-образный. Лингвальный бугорок корня заметен.
Нижние латерозадние зубные ряды (см. рисунок 8): расположенные мезиально нижние латерозадние зубы имеют дистально направленный основной бугорок (например, рис. 8C1). Продвигаясь кзади через зубы нижней челюсти, бугорок постепенно опускается и становится более симметричным. Корень заметно асимметричен в первых двух или трёх латерозадних рядах, с короткой, округлой или четырехугольной дистальной долей и удлиненной сужающейся мезиальной долей, но становится все более симметричным в более задних зубах. Нижние латерозадние зубы отличаются от верхних латерозадних зубов своей прямой или лингвально изогнутой губной стороной бугорка в профиле (в отличие от прямой или лабиально изогнутой губной стороной) и, как правило, более вертикальным бугорком.
• † Cretolamna serrata (Agassiz, 1843) [синоним Otodus serratus Agassiz, 1843; Galeus serratus (Agassiz, 1843); Lamna serrata (Agassiz, 1843); Serratolamna serratus (Agassiz, 1843); Sphyrna serrata (Agassiz, 1843); Lamna libyca Quaas, 1902; Serratolamna libyca (Quaas, 1902); Lamna serra Woodward, 1889; Serratolamna serra (Woodward, 1889); Odontaspis oxyprion Egerton, 1844; Serratolamna oxyprion (Egerton, 1844); Otodus divergens Egerton, 1844; Lamna divergens (Egerton, 1844); Serratolamna divergens (Egerton, 1844)] — 70,6—66,043 млн лет назад, меловой период США (Мэриленд, Северная Каролина). [Agassiz, L. (1838) Recherches sur les poissons fossiles, 11th livraison (November 1838). Petitpierre et Prince(text) and H. Nicolet(plates), Neuchatel, vol. 3: [73]-140, pl. 1a, 8a, 8b, 15, 17, 19, 20, 24, 25b, 30, 31, 32, 33, 34, 35, 39, 40.]
Название вида serrata происходит от латинского слова serra — «пила», «зубчатый», «зазубренный», «пилообразный», ссылаясь на форму зубов этой акулы.
Некоторые учёные относят Cretolamna serrata к роду Serratolamna Landemaine, 1991. Эта акула была зарегистрирована во многих частях мира (Herman 1977), и может быть одним из наиболее широко распространённых таксонов меловых акул. Несмотря на такой широкий палеогеографический диапазон, Cretolamna serrata, по-видимому, существовала недолго. Таксон, был ограничен маастрихтом (72,1—66,0 млн лет назад — Маастрихтский ярус) и хорошо известен из маастрихта Северной Америки (Case 1979, Case and Cappetta 1997), Бельгии и Голландии (Leriche 1929, Herman 1977),
Франции (Landemaine 1991), Марокко (Noubhani and Cappetta 1997), Египта (Wanner, 1902), Анголы (Herman, 1977) и Бразилии (Reboucas and Santos, 1956). Недавно присутствие Cretolamna serrata [три зуба из формации Гвинея Корн (Guinea Corn Formation), Центральный выступ (Central Inlier)] было обнаружено в маастрихте Ямайки, что добавляет известное палеографическое распространение этого обширного вида, который ранее ещё не был зарегистрирован в регионе Карибского моря или Центральной Америки.
Кажущееся глобальным распространение Cretolamna serrata предполагает, что она должна быть чрезвычайно космополитическим видом. Такое широкое распространение потребовало бы длительные переходы через океан, но во время маастрихта южная часть Атлантического и Тетийского океанов были относительно узким. Cretolamna serrata, по-видимому, присутствовала в ряде палео сред, в том числе в кратонных меловых морях Северной Европы, в аргиллитах морского побережья США и конденсированных фосфатных аргиллитах на Тетической окраине Северной Африки. Находки было зарегистрированы как в карбонатных, так и обломочных отложениях, как в пределах неритических зон и в более прибрежных районах. Распространение Cretolamna serrata свидетельствует о том, что это была широко распространенная форма, возможно, живущая как активный пелагический хищник в неритических и прибрежных средах.
Cretolamna serrata известна только по изолированным зубам и, как правило, очень редкими ископаемым скелетными материалами (Cappetta 1987b). Маленькие зубы Cretolamna serrata предполагают гораздо меньшие размеры акулы с рвущимися зубами (Cappetta 1987). Сравнение с аналогичными таксонами со схожими зубными рядами, такими, как некоторые из дошедших до нас виды рода Carcharias, предполагает, что, вероятно, Cretolamna serrata достигала менее 1,5 метра в длину. Это
меньше, чем любые существующие ламниды, хотя и находится в пределах диапазона размеров более мелких видов Carcharias.
• † Cretolamna tethysae Zhelezko 1995 [синоним Cretalamna tethysae Zhelezko 1995] — дан (66,0—61,6 млн лет назад — Датский ярус). [Виктор Иванович Железко (1995) Биостратиграфия фосфоритоносных формаций мела, палеогена Зауралья и Северо-Западного Казахстана по эласмобранхиям.]
Зубы Cretolamna tethysae были обнаружены в фосфоритовом слое (0,8 м), залегающем в основании известняков датского яруса в разрезе реки Бодрак (село Скалистое, Бахчисарайский район) в Крыму.
Название вида tethysae происходит от названия древнего океана Тетис, в котором обитали эти акулы. Tethys — немецкоязычная форма имени греческой богини моря Тефиды (Τηθυς). Океан Тетис существовал в эпоху мезозоя между древними континентами Гондвана и Лавразия. Реликтами этого океана являются современные Средиземное, Чёрное и Каспийское моря.
 1f (Leptostyrax macrorhiza).jpg) |
 1f teeth.jpg) |
 2f teeth.jpg) |
 3f teeth.jpg) |
 4f teeth.jpg) |
 5f teeth.jpg) |
 6f teeth.jpg) |
 7f teeth.jpg) |
 8f teeth.jpg) |
 2f teeth.jpg) |
 1f teeth.jpg) |
 1f teeth.jpg) |
 2f teeth.jpg) |
 1f.jpg) |
 2f teeth.jpg) |
 3f teeth.jpg) |
 4f teeth.jpg) |
 1f teeth.jpg) |
 2f teeth.jpg) |
 3f teeth.jpg) |
 4f teeth.jpg) |
 5f teeth.jpg) |
 6f teeth.jpg) |
 1f teeth.jpg) |
 2f teeth.jpg) |
 3f teeth.jpg) |
 1f teeth.jpg) |
 1f teeth.jpg) |
 2f teeth.jpg) |
 3f teeth.jpg) |
 4f teeth.jpg) |
 5f teeth.jpg) |
 1f teeth.jpg) |
 1f teeth.jpg) |
 2f teeth.jpg) |
 1f teeth.jpg) |
 1f teeth.jpg) |
 1f teeth.jpg) |
 1f teeth.jpg) |
 2f vertebra.jpg) |
 3f teeth.jpg) |
 4f teeth.jpg) |
 5f teeth.jpg) |
 6f teeth.jpg) |
 1f teeth.jpg) |
 1f.jpg) |
 2f teeth.jpg) |
 3f teeth.jpg) |
 4f teeth.jpg) |
 5f teeth.jpg) |
 6f teeth.jpg) |
 1f teeth.jpg) |
 1f teeth.jpg) |
 2f teeth.jpg) |
 3f teeth.jpg) |
 4f teeth.jpg) |
 5f teeth.jpg) |
 6f teeth.jpg) |
 7f teeth.jpg) |
 1f.jpg) |
 2f teeth.jpg) |
 3f teeth.jpg) |
 4f teeth.jpg) |
• Род † Dallasiella Cappetta & Case, 1999 (Далласиеллы) — 99,7—94,3 млн лет назад, меловой период, Канада (Саскачеван), США (турон (93,9—89,8 млн лет назад — Туронский ярус) Карлайл Шейл (Carlile Shale), округ Эллис (Ellis County), штат Канзас; штат Техас). [Henri Cappetta & Gerard Ramon Case (1999) Additions aux faunes de sélaciens du Crétacé du Texas (Albien supérieur-Campanien). Palaeo Ichthyologica, 9, 5–111]
Зубы Dallasiella напоминают зубы Cretolamna и в меньшей степени зубы Protolamna, но отделяются от них корнем с хорошо дифференцированной бороздкой.
• † Dallasiella willistoni Cappetta & Case, 1999 [синоним Dallasiella cf. willistoni Cappetta & Case, 1999] — 99,7—94,3 млн лет назад, Bainbridge River Bonebed, Саскачеван, Канада. [Henri Cappetta & Gerard Ramon Case (1999) Additions aux faunes de sélaciens du Crétacé du Texas (Albien supérieur-Campanien). Palaeo Ichthyologica, 9, 5–111]
Вид назван в честь палеонтолога Уиллистона (Williston) за его работу по техасским акулам.
Материал (см. табл. 13): около двадцати зубов. Происхождение: Kiest Boulevard, Даллас. Возраст: турон (93,9—89,8 млн лет назад — Туронский ярус) — коньяк (89,8—86,3 млн лет назад — Коньякский ярус). Голотип: табл. 13, рис. 3.
Описание: лектотип представляет собой нижний латерально-задний зуб (табл. 13. рис. 3). Основной бугорок не очень высокий, прямолинейный в профиль, губная поверхность равномерно выпуклая и гладкая. Есть пара довольно широких боковых зубчиков, чётко прикреплённых к режущим кромкам центрального бугорка. Лопасти корня довольно длинные, дистальный конец у них более широкий, чем мезиальный. Базилярная (относящаяся к основанию) вырезка широкая, глубокая и пористая. Лингвальный выступ умеренно выступает и несёт короткую, но глубокую борозду в том месте, где открывается большое отверстие.
Передний зуб (табл. 13. рис. 1) более узкий, корень не толстый с более длинной бороздой, особенно развитой по направлению к базилярному краю.
Парасимфизный зуб (табл. 13. рис. 2) имеет основной бугорок не выше корня.
Нижний боковой зуб (табл. 13, рис. 4) симметричный, с менее расходящимися в поперечном направлении боковыми зубцами и уплощёнными концами. На язычном выступе, более заметно, чем на переднем зубе, открывается большое отверстие, от которого поверхностная, но острая борозда достигает базилярного края. Лунула (видимая часть корня) широкая и глубокая.
Нижние боковые зубы (табл. 13, рис. 5. 8—9). Основной бугорок расширяется: в средней части губной поверхности корня наблюдается множество эллиптических отверстий. Очень узкие складки могут дифференцироваться в основании латеральных областей лингвальной стороны бугорка. Корень высокий и довольно плоский.
Боковые зубы, вероятно, верхние (табл. 13. рис. 6—7). Основной куспид сильно наклонен к комиссуре, имеет выпуклый мезиальный край и вогнутый дистально. Их губная поверхность практически такая же выпуклая, как и язычная. Корень очень развитый, с широким базилярным вырезом. Базилярное лицо широкое и довольно плоское; язычный бугор остается хорошо заметным и несет четкую борозду.
На некоторых боковых зубах (табл. 13. рис. 10) боковые зубчики могут расщепляться, и, возможно, на боковом зубе (табл. 13. рис. 9) разделились на два зубца.
Обсуждение: Описанные зубы, по-видимому, хорошо соответствуют тем, которые представлены Мейер (Meyer) из турона и нижнего коньяка в Техасе под названием «Odontaspis subulata». Немногочисленные зубы сеномана (100,5—93,9 млн лет назад — Сеноманский ярус), представленные Мюллером и Дидрихом (Muller & Diedrich) как «Cretolamna arcuata» практически идентичны таковым зубам Dallasiella willistoni из Техаса.
По сравнению с Protolamna cacteri из сеномана виды из Kiest Boulevard имеют более крупные зубы; передние ряды полностью гладкие, а из сеномана — слегка складчатые. На уровне боковых зубчиков зубы Kiest Boulevard более плоские лабиально и чаще всего лишены складок.
Некоторые зубы, как передние, так и боковые, у Dallasiella willistoni могут быть похожи на зубы Cretolamna appendiculata; однако они легко отличаются корнем с язычным выступом с хорошо заметной бороздкой и, как правило, более толстым бугорком, особенно в боковых выступах. Кроме того, зубы Daiiasieia имеют бугорок с более округлой губной гранью и довольно многочисленными и развитыми отверстиями в средней области губной поверхности корня.
Среди семейства Cretoxyrhinidae Gluckman, 1958 у рода Sorratolamna также есть зубы с чёткой бороздкой у корня; их дентальная морфология, однако, очень отличается от таковой у Dallasiella, и нет никакой путаницы между этими двумя родами, зубы у которой выше и с более чёткими зубцами.
Зубы Dallasiella willistoni так же очень похожи на зубы Scapanorhynchus raphiodon, которые Майк Эверхарт (Mike Everhart) изначально не точно идентифицировал (см. фото).
 1f teeth.jpg) |
 2f teeth.jpg) |
 3f teeth.jpg) |
 4f teeth.jpg) |
• Род † Paraisurus Glikman, 1957 [синоним Parisurus Glikman, 1957, orth. var.] — 125,0—99,7 млн лет назад. [Glikman, L.S. (1957) (Taxonomic significance of the accessory teeth of sharks of the families Lamnidae and Scapanorhynchidae). «in russian». Trudy geol. Muz. «A.P. Karpinskogo» Akad. Nauk S.S.S.R., 1: 103–109, 2 fig.]
Название рода Paraisurus происходит от латинского слова para — «рядом», «возле», «около» и названия рода ламнообразных акул Isurus Rafinesque, 1810, указывая на их схожесть.
Это чрезвычайно редкий вымерший род. Иногда его относят к incertae sedis («неопределённого положения» — латинское выражение, указывающее на то, что положение таксона в системе неясно). Эти акулы жили в раннем меловом периоде и известны в Европе, Северной Америке и Азии. Они чаще всего известны по отдельным зубам, которые имеют особую форму (см. рисунок). Они мезио-дистально сдавлены, с большим губным бугорком корня. Связанный зубной ряд известен из формации Вено (Weno Formation), штат Техас, США. Первоначально предполагалось, что виды рода Paraisurus вымерли к концу альба (113,0—100,5 млн лет назад — Альбский ярус), но находки из формации нижнего Атко (lower Atco formation) предполагают их проживание в раннем коньяке (89,8—86,3 млн лет назад — Коньякский ярус).
Из верхнего альба Швейцарии, Сент-Круа (Saint Croix), Франсуа Жюль Пикте (François Jules Pictet) и Гюстав Кампиш (Gustave Campiche) описали зубы акул под названием Oxyrhina macrorhiza. Характерные признаки: длинный, раздвоенный на конце корень, обе ветви более или менее сжаты с боков и образуют между собой острый угол. Коронка, рассматриваемая в профиль, очень утолщена у основания; нижняя граница эмали, спускающаяся очень низко с внешней стороны, с внутренней резко поднимается, покрывая менее половины той высоты, какую она занимает с наружной. При рассмотрении с внутренней стороны зуб напоминает туфлю; часть зуба, лишённая эмали, соответствует подъёму ступни. Вершина коронки отогнута внутрь. Наружная сторона у основания обычно несколько сжата с боков; края коронки режущие; по середине внешней стороны наблюдается продольное ребро, особенно хорошо выраженное у основания; это ребро иногда сопровождается с боков двумя или четырьмя базальными бороздками. Характерная особенность по Ф. Пикте и Г. Кампишу — полное отсутствие боковых зубчиков, отличающее род Oxyrhina (Isurtis). Но авторы очень широко понимали этот вид и отнесли к нему ряд зубов очень плохой сохранности, не только видовая, но и родовая принадлежность их сомнительна.
Л. С. Гликман, учитывая своеобразие зубов Oxyrhina macrorhiza, выделил их в новый род Paraisurus. Диагноз по Л. С. Гликману: «Коронка крайне толстая и короткая. По форме напоминает клык. Боковые зубы имеют тенденцию к расширению. Добавочные зубцы у передних зубов отсутствуют. Корни необычайно массивные. Ветви их сливаются с внутренней стороны зуба». К сожалению, описание Л. С. Гликмана отличается не только большой краткостью, но и неопределенностью. Какие боковые зубы, имеющие тенденцию к расширению, подразумевает автор? Если «передние» зубы хорошо описаны Пикте и Кампишем, то зуб, который Л. С. Гликман считает боковым, необходимо было описать более подробно и дать не только изображение с внутренней стороны, но и сбоку. Если судить по изображению, то этот зуб можно отождествить с Megarhizodon macrorhiza (боковой верхнечелюстной зуб). Зубы Paraisurus встречаются очень редко; в моей коллекции — пишет Михаил Иванович Соколов, имеется всего 20 зубов (не считая пяти зубов, полученных мной от Р. Кэси из кембриджских зеленых песков Англии). Но даже имеющиеся в моем распоряжении зубы и описанные Пикте и Кампишем, распределённые по возрасту, позволяют описать четыре вида рода Paraisurus.
Зубы акул из рода Paraisurus являются одними из самых редких находок Пластиножаберных (Elasmobranchii). Для этого рода характерны массивные клыковидные коронки и относительно длинные язычные выступы корней, более или менее сжатые с боков корневых ветви. Род Paraisurus быстро эволюционировал в позднем апте (125,0—113,0 млн лет назад — Аптский ярус), и вымер альбе (113,0—100,5 млн лет назад — Альбский ярус), как раз на границе раннего и позднего мелового периода. Поэтому зубы акул Paraisurus полезны для отслеживания морских и солёных эстуарных отложений, а также для датирования отложений. Изолированные зубы акул указанного рода были известны в верхнем альбе Швейцарии, Франции, Англии и США. Недавно зубы такого же размера были обнаружены в России, Литве, Украине, Узбекистане и Казахстане. Самый старый известный зуб рода Paraisurus с некоторыми примитивными особенностями (описанными ниже) был обнаружен в верхнем апте Каракалпакстана (западный Узбекистан).
Замечания. Все останки Paraisurus были обнаружены в местах с признаками интенсивного шельфового и прибрежного апвеллинга (апвеллинг (англ. upwelling) — подъём глубинных вод океана к поверхности): например фосфоритовые конкреции, отсутствие карбонатного сернента в матриксе, обильные фосфатизированные части скелетов крабов и раков, много фосфатизированных капролитов беспозвоночных и рыб, многочисленные останки морских и устьевых хищников (других акул, актиноптеригов, а иногда и морских черепах, плезиозавров, ихтиозавров). Очевидно, это была высокая биологическая продуктивность в воде и большая биомасса крупных потребляющих веществ, вероятно, съеденных акулами из рода Paraisurus. Строение зубов Paraisurus показывает, что их можно использовать для отрыва больших кусков от тел своей добычи, возможно, от других акул, больших костистых рыб, плезиозавров и, вероятно, ихтиозавров, обитавших в довольно открытых частях морей. Вымирание акул рода Paraisurus, возможно, было вызвано замедлением апвеллинга и снижением биологической продуктивности после окончания последней альбской трансгрессии, после изменения температурного режима океана, связанного с реконструкцией системы обитания.
• † Paraisurus amudarjensis Mertiniene, Nesov & Nazarkin, 1994 — апт (125,0—113,0 млн лет назад — Аптский ярус). [Romualda Mertiniene, Lev Alexandrovich Nesov & Mikhail Valerievich Nazarkin, 1994, Cretaceous Shark of the Genus Paraisurus in Uzbekistan and Lithuania.]
Назван вид amudarjensis в честь реки Амударья.
Голотип. Центральный музей геологоразведки (ЦМГЭ), Санкт-Петербург, Nr. 123/12178. Материал. 1 зуб (см. рисунок).
Типовая местность: северо-восточное побережье высохшего озера Каракуль; Каракалпакстан, Западный Узбекистан, площадка СЗБ-3. Стратиграфия и формация: свита Сулианбобо; Нижний мел, верхний апт (род), зона Acanthoplites nolani.
Диагноз. Зуб маленький. Губная сторона коронки в 2,2 раза больше её ширины. Ямка между корневыми ветвями на губной поверхности переходит в неглубокую борозду коронки. Лингвальный отросток корня широкий (с лингвальной точки зрения). Ветви (выступы) корня высокие, хорошо обособленные, более ориентированы в стороны, чем у других видов рода, и имеют почти одинаковую толщину по всей длине; концы ветвей тупые.
Описание. Высота зуба 11,4 мм; губная сторона коронки в 2,2 раза больше её ширины. Коронка относительно широкая у основания и круто суженная к самой верхней 1/4 их высоты, бугорок резко заострён. Губная поверхность умеренно выпуклая; посередине губной поверхности коронки имеется бороздка. Эта бороздка связана с относительно большой ямкой, расположенной между корневыми ветвями. Корень зуба массивный и очень высокий, язычный выступ широкий (с лингвальной точки зрения) и умеренно длинный. Ветви корня высокие, относительно толстые, хорошо разделённые, ориентированные под относительно большим углом к оси коронки, их концы тупые и почти одинаковой толщины по всей длине.
Сравнение. Новый вид отличается от Paraisurus macrorhiza и всех других известных видов рода Paraisurus, описанных М. Соколовым из разных уровней альба Казахстана, своими примитивными чертами: небольшие размеры, бугорок коронки относительно острый, выпуклость основания язычной стороны коронки умеренная, язычный вырост корня не очень длинный, ветви корня тупые и ориентированы больше в стороны, чем у корней другие виды рода.
Замечания. Образец был найден в обширном комплексе останков позвоночных, включая зубы других ветвей Пластиножаберных: Polyacrodus, Hybodus, Heterodontus, Paraorthacodus, Cretorectolobus, Rhinobatos, Synodontaspis (= Eostriatolamia), Odontaspisilna, Lexatpisilna), зубы голостеевых рыб (Holostei — костные ганоиды) Lepidotes, кости (Teleostei — настоящие костистые рыбы) Albulidae, Blochiidae, панцири черепахи ср. Adocus sp. и ср. Plesiochelys sp., зубы и остеодермы метамезозухиальных крокодилов (metamesosuchian crocodiles), остатки зауроподов с тонкими длинными зубами и мелких (возможно, молодых) плезиозавров (Elasmosauridae), раков и аммонитов. Позднеаптские акулы известны в регионе только в Северо-Восточной Фергане (Кыргызстан), но относятся к склерофагам солоноватоводным видам рода Asiadontus (Asiadontinae, Polyacrodoniidae).
• † Paraisurus compressus Sokolov, 1978 [синоним — Paraisurus macrorhiza Kemp, 1991] — 112,6—99,7 млн лет назад. [Михаил Иванович Соколов, 1978, Зубы акул как руководящие ископаемые при зональном расчленении меловых отложений Туранской плиты.]
Описание (по Соколову). Можно предполагать, что массивные зубы Oxyrhina macrorhiza, изображенные на рис. 6 и 7 табл. X, и зубы, сильно сжатые с боков, изображенные на рис. 8—10 той же таблицы у Пикте и Кампиша, принадлежат одному виду, но занимают различное положение в челюстях. Однако изучение вертикального распределения зубов Paraisurus показывает, что массивные зубы занимают более низкое стратиграфическое положение, сильно же сжатые зубы встречаются только во враконских отложениях (вракон — верхняя часть верхнеальбского подъяруса, иногда выделяемая в особый (четвертый) враконский подъярус альбского яруса), причем ни одного массивного зуба во враконе не было встречено. Пять зубов из кембриджских зеленых песков Англии, т. е. также происходящие из вракона, относятся к тому же типу. Основные признаки, характерные для рода, приложимы и к Paraisurus compressus. Основные отличия этого вида — сильное сжатие коронки с боков. Если у Paraisurus macrorhiza высота зуба превышает ширину (замеренную у основания эмали с наружной стороны) примерно в два раза, то у Paraisurus compressus высота превышает ширину почти в три раза. Корень образует значительно более выдающийся сильно сжатый с боков выступ с внутренней стороны, чем у Paraisurus macrorhiza, образуя как бы седло, на котором сидел зуб, расположенный позади. При рассмотрении зуба в профиль видно, что линия, ограничивающая сверху выступ корня, всегда перпендикулярна к очертанию наружной стороны зуба; последняя обычно описывает очень пологую правильную дугу, поэтому вершина коронки слегка отогнута внутрь, иногда она прямая. Несколько варьирует форма корня, который всегда внизу раздвоен, но ветви его то короткие, сжатые с боков, с желобком, расширяющимся в сторону раздвоенной части корня, то очень длинные, направлены вниз и образуют высокую арку. Между двумя типами корней наблюдаются постепенные переходы. По-видимому, это различие в строении корня связано с положением зубов в челюсти.
Возраст и распространение. Типичные Paraisurus compressus описаны из враконских песков Сент-Круа (Saint Croix) Швейцарской Юры; встречаются в кембриджских зеленых песках Англии, во враконе Восточного Мангышлака, Казахстан, нижняя или средняя часть ходжакульской свиты (см. рисунок). Таким образом, их стратиграфическое положение может считаться твердо установленным враконские отложения Евразии. Найдены так же в меловых отложениях Австралии, Блэк Дам (Black Dam).
Описание. Коронка у основания на губной поверхности более чем в 3 раза больше её ширины. Зубы сравнительно узкие, корни сдавлены с боков. Коронка постепенно сужается от основания к бугорку. Лингвальное лицо гладкое, очень выпуклое. На некоторых зубах есть вертикальные борозды. Губная лицевая сторона слегка выпуклая в основании и вогнутая у бугорка. Корень сильно сжат с боков, язычный выступ узкий (с лингвальной точки зрения) и длинный, с каждой стороны от него расположены относительно глубокие ямки. Ветви корня тонкие, сравнительно длинные, сильно сжатые с боков, концы их острые, иногда лопаточные.
Сравнение. Зубы Paraisurus compressus более узкие и изящные, чем зубы Paraisurus macrorhiza (Pict et Camp.). Корень зубов Paraisurus compressus более сдавлен с боков и значительно уже корня Paraisurus macrorhiza и Paraisurus amudarjensis. Лингвальный отросток корня намного заметнее и сильнее сжат с боков, чем у всех других видов этого рода.
• † Paraisurus elegans Sokolov, 1978 — альб (113,0—100,5 млн лет назад — Альбский ярус). [Михаил Иванович Соколов, 1978, Зубы акул как руководящие ископаемые при зональном расчленении меловых отложений Туранской плиты.]
Описание. Зубы этого вида сильно отличаются от зубов других представителей рода Paraisurus: коронка более высокая, более стройная и более заострённая. Корень массивный, но значительно менее выдающийся с внутренней стороны, чем у описанных видов. Очертание зуба в профиль с наружной стороны образует пологую выпуклую дугу; с внутренней стороны коронка при переходе в корень не образует почти прямого, притупленного угла, как у Paraisurus compressus, а плавно продолжает пологую вогнутую дугу коронки, описывая часть окружности лишь несколько меньшего радиуса. Корень полностью не сохранился, но, восстанавливая его по сохранившимся ветвям, можно заключить, что ветви были короткие и расходились под углом около 100°. Толщина коронки и угол вершины у отдельных экземпляров варьирует в довольно значительных пределах (угол вершины от 25 до 50°). Эта форма более примитивная, чем описанные выше, и характерные особенности рода выражены слабее.
Возраст и распространение. Все шесть экземпляров найдены в отложениях зоны «mammillatum» нижнего альба Мангышлака.
• † Paraisurus lanceolatus Sokolov, 1978 — альб (113,0—100,5 млн лет назад — Альбский ярус). [Михаил Иванович Соколов, 1978, Зубы акул как руководящие ископаемые при зональном расчленении меловых отложений Туранской плиты.]
Описание. В сравнении с Paraisurus macrorhiza зубы более массивные, коронка более толстая, выступ корня более выдающийся, но зуб более стройный и его боковые стороны параллельны и книзу не расширяются. Корень явственно раздвоен, сильно сжат с боков и ветви его почти параллельны. Наружная сторона зуба у Paraisurus macrorhiza очерчивает очень слабо выпуклую наружу дугу, у Paraisurus lanceolatus эта дуга более выпуклая. Но наиболее характерной особенностью описываемого вида является сильная сжатость с боков вершины корня, которая образует высокую узкую пирамиду; с наружной стороны у основания зуба имеется узкий желоб.
Возраст. Единственный зуб найден в отложениях зоны «dentatus» среднего альба Мангышлака.
• † Paraisurus macrorhiza (Pictet & Campiche, 1858) [синоним — Oxyrhina macrorhiza Pictet & Campiche, 1858; Paraisurus macrorhizus (Pictet & Campiche, 1858)] — типовой вид — нижний мел: верхний апт (125,0—113,0 млн лет назад — Аптский ярус) — верхний альб (113,0—100,5 млн лет назад — Альбский ярус) Узбекистана, верхний альб Швейцарии, Франция, Англия, Россия, Литва, альб Казахстана. [François Jules Pictet & Gustave Campiche, 1858-1860, Description des Fossiles du Terrain Crétacé des Environs de Sainte-Croix; Francois Jules Pictet, 1858, Notitz über die Fische der Kreide-Gebilde in der Schweitz und Savoyen in Bibliothèque Universelle de Genève.]
Из верхнего альба Швейцарии, Сент-Круа (Saint Croix), Франсуа Жюль Пикте (François Jules Pictet) и Гюстав Кампиш (Gustave Campiche) описали зубы акул под названием Oxyrhina macrorhiza. У изображенных авторами экземпляров зубов не сохранился полностью корень. По-видимому, две ветви корня были обособлены довольно хорошо. Л. С. Гликман, учитывая своеобразие зубов Oxyrhina macrorhiza, выделил их в новый род Paraisurus.
Распространение. По Ф. Пикте и Ж. Кампишу, зубы Oxyrhina macrorhiza встречаются в верхнем альбе (gault superieur). Вероятно, они приурочены к более низким горизонтам верхнего альба, поскольку во враконе встречается другой вид, Paraisurus compressus. Так же зубы Paraisurus macrorhiza были найдены в экспозиции Варейкяй, Йонавский край, Литва (нижний мел, верхний альб (вракон), Свита Джесиа) и в Лебединском карьере недалеко от города Губкин, Белгородская область, Россия, (верхний альб, нижняя часть секменевской свиты, тонкий слой с массивными конкрециями фосфоритов).
Описание (см. рисунок). Зубы массивные; коронка у основания на губной поверхности примерно в 2 раза больше её ширины. Коронка похожа на клык, толстая и короткая. Коронка постепенно сужается от основания к бугорку, но ближе к бугорку она внезапно сужается. Бугорок заострён и наклонен вперед (лабиально). Губное лицо слегка выпуклое или плоское; продольная вертикальная складка расположена посередине губной поверхности. Иногда по бокам от него встречаются две вертикальные короткие борозды. Лингвальная морда гладкая, у основания макушки выпуклая. Корень массивный, высокий, ветви относительно хорошо разделены. Лингвальный выступ корня очень длинный и большой.
Сравнение. Зубы Paraisurus macrorhiza больше, чем у Paraisurus amudarjensis, и имеют продольную вертикальную складку на губной поверхности коронки. У этого вида более толстая и короткая крона и менее сжатый с боков корень, чем у Paraisurus compressus.
Диагностика. Зубы массивные, мезио-дистально сжатые. Коронка относительно низкая по сравнению с высотой корня и широкая для одних зубов, но относительно узкая для других. Губная грань коронки обычно гладкая или с продольной складкой; иногда по обе стороны от её есть две вертикальные короткие борозды. Лингвальная грань короны заметно выпуклая. Нижняя граница эмелоида на губной поверхности коронки спускается очень низко, покрывает часть боковых краев корневых ветвей и составляет острый угол. Эмелоид лингвального лица покрывает почти половину высоты эмелоида губной стороны. Корень сильно развит, очень высокий. Лингвальный выступ корня длинный, умеренно широкий или узкий, зуб в профиль напоминает сапожок. Ветви корня хорошо разделены. Они относительно длинные губно-язычно, обычно более или менее сдавлены с боков.
Единственный в Поволжье разрез с представительным комплексом рыб нижнего сеномана располагается на севере Волгоградской области (Меловатка-5). В 2010–2011 гг. здесь отобраны две объемные пробы (около 100 кг) из нижнего фосфоритового слоя (Иванов, 1997; Первушов и др., 1999), одна из которых (СГУ, № 329, 2010 г.) обработана. Комплекс эласмобранхий определён предварительно (1321 экз.). Встречены ламноидные акулы (доминируют): Eostriatolamia subulata, Cretolamna appendiculata, Cretoxyrhina cf. denticulata, Leptostyrax cf. macrorhiza., Johnlongia sp., Archaeolamna cf. kopingensis, Anomotodon sp., Palaeoanacorax cf. volgensis, Paraisurus macrorhizus, синеходонтиформы Synechodus dispar, Paraorthacodus recurvus, гибодонты Polyacrodus sp., морские ангелы Squatina sp., ковровые акулы Cederstroemia sp., катраны Protosqualus sp., разнозубые акулы Heterodontus sp., есть редкие скаты (Turoniabatis sp., Squatirhina sp.). В целом, комплекс типично сеноманский, но с отсутствием ряда таксонов (Ptychodus spp. встречены в регионе в верхнем сеномане; нет пока находок кархаринообразных акул). Интересно присутствие в пробе единственного зуба Paraisurus macrorhizus, типичного для альба, что может быть результатом переотложения и/или более позднего существования рода в регионе. Гликман (1957) отмечал находку зуба Paraisurus macrorhizus в недалеко расположенном разрезе Красный Яр, но современная датировка уровня находки соответствует среднему сеноману (100,5—93,9 млн лет назад — Сеноманский ярус).
 1f teeth.jpg) |
 1f teeth.jpg) |
 2f teeth.jpg) |
 1f teeth.jpg) |
 2f teeth.jpg) |
 |
• † [синоним] — дан. []
• Род † Plicatolamna Herman, 1975
• † Plicatolamna arcuata (Woodward, 1894) [syn. Plicatolamna arcuata Woodward, 1894]
• Род † Serratolamna Landemaine, 1991