(Cnidaria Hatschek, 1888) Тип Книдарии, Стрекающие, Phylum Cnidaria Hatschek, 1888 (Cnidarians) 5 классов
Жизнь животных. Том 1. Беспозвоночные Под редакцией профессора Л.А.Зенкевича 1968 г.
ТИП КИШЕЧНОПОЛОСТНЫЕ (COELENTERATA)
Кишечнополостные — самые низко организованные из числа настоящих многоклеточных животных. Тело кишечнополостных состоит из двух слоев клеток — эктодермы и энтодермы, между которыми имеется более или менее развитый неклеточный слой, называемый мезоглеей. Свое название эти животные получили в связи с тем, что у них имеется всего одна полость, называемая кишечной или гастральной полостью. Все кишечнополостные — водные, за исключением немногих видов, морские организмы. Их тело построено по типу радиально-осевой симметрии.
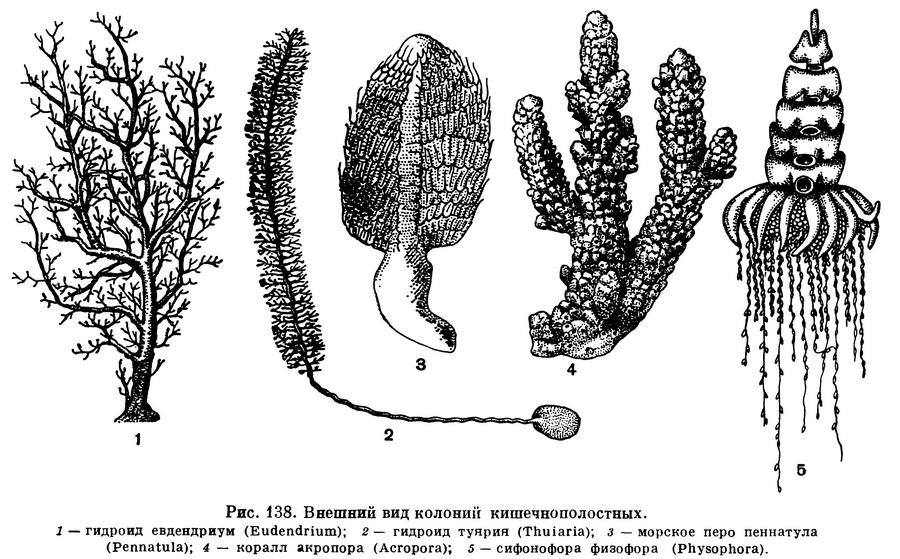
Несмотря на простоту организации, внешне кишечнополостные очень разнообразны. Это зависит от двух причин. Первая причина заключается в способности кишечнополостных образовывать колонии. Как правило, отдельная особь колонии очень невелика, и потому в первую очередь внимание невольно обращается на всю колонию в целом. Одни колонии имеют вид кустиков или маленьких деревьев. Другие похожи на ершик, третьи напоминают фантастические птичьи перья. Кроме таких нежных гибких колоний, есть колонии массивные с мощным известковым скелетом. Они имеют то форму шара, то кубка, то гриба, то колючей елочки. Наконец, встречаются нежные плавающие колонии.
Вторая причина разнообразия внешнего вида кишечнополостных зависит от того, что у этого типа животных отдельная особь имеет форму либо полипа, либо медузы. Тело полипа обычно цилиндрическое, на верхнем его конце расположен рот, окруженный щупальцами. Полипы — малоподвижные или даже прикрепленные животные, часто они образуют колонии. Медузы — одиночные плавающие, подвижные организмы. Их тело имеет форму зонтика со щупальцами по краям. Плавают медузы ртом вниз.
Наличие у кишечнополостных двух форм, двух состояний — полипоидного и медузоидного, способность этих животных образовывать колонии разнообразной формы, а также присущая им яркая окраска делают кишечнополостных очень многообразной по формам группой животных, несмотря на то что их внутреннее строение имеет единый, общий план и устроены они довольно просто.
В настоящее время известно около 9 тыс. видов, относящихся к этому типу. Самые маленькие из них (например, полипы на колониях гидроидов) едва достигают 1 мм, наиболее крупные, такие, как медуза цианея (Cyanea arctica), имеют зонтик до 2 м в диаметре, а щупальца этой медузы растягиваются на 30 м.
Кишечнополостные — самые древние из настоящих многоклеточных животных. За время длительной истории развития этого типа его представители сумели очень хорошо приспособиться к самым разнообразным условиям обитания. Они заселили буквально весь океан от его поверхности до предельных глубин, их можно обнаружить и в полярных областях и в тропиках. Поселяются кишечнополостные на самых разнообразных грунтах, некоторые из них способны выносить значительное изменение солености морской воды, а отдельные виды проникли даже в пресные воды. Почти повсеместно они играют очень важную роль в образовании сообществ морских животных и растений — биоценозов моря. Познакомимся же поближе с этими интересными существами.
ИСТОРИЯ ИЗУЧЕНИЯ КИШЕЧНОПОЛОСТНЫХ
Интерес к изучению фауны морских беспозвоночных возник еще в глубокой древности.
Первые сведения о кишечнополостных животных мы находим в трудах Аристотеля. Правда, в его системе они не представляют собой единой группы животного царства. Отдельные кишечнополостные описывались Аристотелем в разных разделах системы. Основная же часть кишечнополостных объединялась в группу Cnidae, или стрекающих. Аристотель подробно описывает их неподвижность, отмечает наличие в теле стрекающих только одного отверстия — рта, имеющего центральное положение и окруженного кольцом подвижных щупалец. Древнегреческому ученому уже было известно, что в щупальцах находятся особые стрекающие органы, служащие для защиты от врагов и для схватывания добычи. Аристотелю принадлежит и очень интересное утверждение о том, что кишечнополостные и вместе с ними другие просто устроенные беспозвоночные, ведущие прикрепленный образ жизни (губки, асцидии, мшанки), представляют собой организмы, в которых неожиданно смешивается природа животных и растений. Эта двойственность природы кишечнополостных, по его мнению, выражалась в том, что они своей способностью к питанию животной пищей и к восприятию внешних раздражений напоминают животных, а общим планом строения, простотой организации и прикрепленным образом жизни — растения. Вслед за Аристотелем и другой древний философ и натуралист Плиний также усматривал двойственную природу кишечнополостных.
Восемнадцатый и девятнадцатый века были временем накопления новых сведений о строении и образе жизни кишечнополостных. В 1723 г. интересное открытие было сделано французским судовым врачом Пейсоннелем. Он установил, что рифообразующие кораллы — колониальные кишечнополостные животные, обладают массивным известковым скелетом. А ведь до его открытия и мореплаватели и ученые считали, что коралловые полипняки принадлежат к неорганическому миру и представляют собой особый известковый минерал. Пейсоннелю удалось наблюдать, как полипы рифообразующих кораллов захватывали своими щупальцами проплывавших мимо мелких рачков и затем поедали их.
Но самой яркой страницей в истории изучения кишечнополостных явились исследования натуралиста Трамбле, швейцарца по происхождению, увлекавшегося, как и многие другие в то время, наблюдением при помощи микроскопа поведения различных мелких животных — инфузорий и «насекомых» (Так назывались в то время не только настоящие насекомые, но и многие другие мелкие животные — коловратки, черви и т. д.). Случайно он обнаружил прикрепившихся к водорослям пресноводных гидр — мелких одиночных гидроидных полипов, которых принял сначала за водные растения. Трамбле ничего не знал тогда о гидрах. Да это и не удивительно, так как вообще почти ничего не было известно о них и гидры не имели даже зоологического названия. Трамбле назвал гидру «полипом с руками в виде рогов». Результатом длительных наблюдений над гидрами явилась книга «Мемуары о пресноводном полипе», вышедшая в 1744 г. Трамбле с удивительной полнотой и точностью описал строение гидры, ее размножение, питание, способы передвижения и т. д. Но главным достижением натуралиста следует считать открытые экспериментальным исследованием способности гидры к регенерации, т. е. к самовосстановлению. Регенерация у других организмов была известна и раньше, но в предшествовавших экспериментах удавалось наблюдать восстановление лишь отдельных утраченных частей животного, например лапок или усиков у ракообразных. У гидры же Трамбле наблюдал нечто большее — восстановление целого организма из отдельных его частей. Эта способность рассматривалась Трамбле и его современниками как особого рода размножение животных путем рассечения его на части. Работы Трамбле представляли собой большой интерес. С одной стороны, его наблюдения окончательно доказали животную природу гидры и вообще остальных зоофитов. С другой стороны, эти исследования привели к отрицанию мнения об основном различии между животными и растениями, до того времени господствовавшего и заключавшегося в том, что растения могут размножаться черенками и отводками, а животные — нет.
В середине прошлого века зоолог Лейкарт создает самостоятельный тип животных — кишечнополостных, выделяя его из типа Zoophita.
СТРЕКАТЕЛЬНЫЕ КЛЕТКИ КИШЕЧНОПОЛОСТНЫХ И ДЕЙСТВИЕ ИХ ЯДА
Характернейшей особенностью всех кишечнополостных является наличие у них особых стрекательных, или крапивных, клеток. Эти клетки развиваются из недифференцированных промежуточных клеток эктодермы. Каждая стрекательная клетка содержит хитиноидную капсулу овальной или продолговатой формы. Стенка наружного конца капсулы впячивается внутрь ее и имеет вид тонкой спирально закрученной трубочки, называемой стрекательной нитью. Полость капсулы заполнена ядовитой жидкостью.
На внешней поверхности стрекательной клетки имеется чувствительный волосок — книдоциль. Прикосновение к волоску вызывает моментальное раздражение клетки, проявляющееся в стремительном, подобном выстрелу выворачивании стрекательной нити. В зависимости от деталей строения и способа воздействия на врага или жертву различают несколько типов стрекательных капсул. Часть стрекательных капсул имеет длинную, снабженную шипами нить. При «выстреле» такой капсулы нить вонзается в тело жертвы, а через ее полость изливается содержимое капсулы, вызывающее общее или местное отравление. У других капсул нить короткая, лишенная шипов. Такие нити только опутывают жертву. Наконец, имеются липкие нити, которые удерживают добычу приклеиванием. Они же могут служить и для временного прикрепления самих носителей стрекательных клеток при передвижении.

Стрекательные клетки располагаются по всей поверхности тела кишечнополостных, но их наибольшие скопления наблюдаются на щупальцах и вокруг ротового отверстия, т. е. в местах, где они нужнее всего. После «выстрела» стрекательная клетка погибает и на ее месте или поблизости от него развивается новая стрекательная клетка.
Весьма интересно, что стрекательные капсулы могут действовать и после гибели животного. Так, прикосновение к мертвым, выброшенным на берег медузам может вызвать довольно сильные покраснения кожи, сопровождающиеся ощущением жжения.
При поедании кишечнополостных животных некоторыми турбелляриями и заднежаберными моллюсками часть стрекательных клеток переходит в тело хищника. Здесь стрекательные клетки располагаются также в наружных покровах и могут нормально функционировать.
Стрекательные клетки, особенно содержащие ядовитую жидкость в капсулах, являются грозным оружием, которое используется кишечнополостными и для защиты и для нападения. Несомненно, что процветание этих низко организованных животных в значительной мере объясняется наличием у них стрекательных клеток. Мелкие животные, соприкасаясь с кишечнополостными, приклеиваются, опутываются и пронзаются стрекательными нитями, а в их тело попадает яд, вызывающий паралич и смерть. После этого жертва переносится щупальцами к ротовому отверстию и проглатывается. Даже довольно крупные животные часто оказываются пораженными ядом стрекательных капсул, вызывающим к тому же жгучую боль. Сила воздействия яда разных видов кишечнополостных на человека неодинакова: некоторые из них совершенно безвредны, другие представляют собой серьезную опасность.
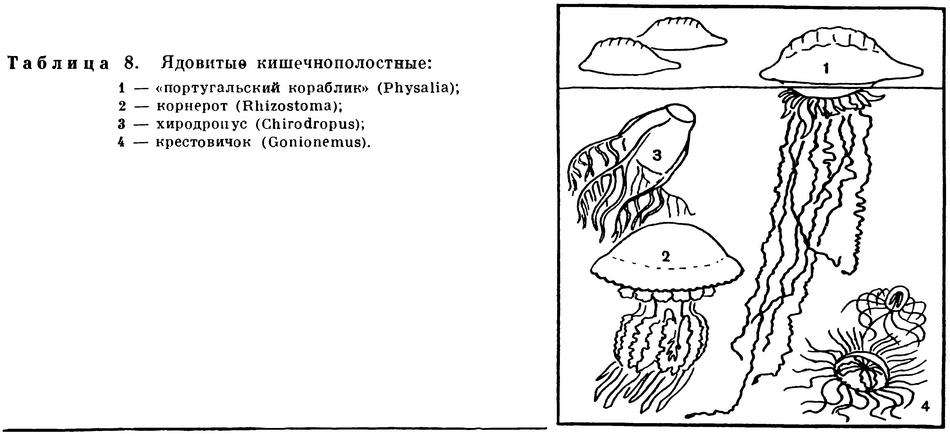
После прикосновения к нашим черноморским актиниям и к медузам цианея, обитающим в арктических и дальневосточных морях, ощущается легкое жжение, особенно там, где кожа нежнее, например между пальцами рук. Несколько сильнее «жжется» корнерот — обычная для Черного моря беловатая медуза с фиолетовым краем зонтика. Тем не менее эти неприятные ощущения, похожие на легкий ожог крапивой, быстро проходят, не оставляя следа. Однако в море имеется много гораздо более опасных кишечнополостных, прикосновение к которым может привести к тяжелому заболеванию. Так, многие актинии, в том числе обитающие в Японском море, вызывают не только «ожоги», но и длительное опухание руки неосторожно дотронувшегося до них человека.
Однако наиболее неприятные последствия заключаются не в «ожогах» и опухолях, а в общем отравлении организма ядом стрекательных клеток. У моряков, плавающих в тропических морях, уже издавна пользуется дурной славой красивая сифонофора физалия. Физалия снабжена большим, до 20 см длиной, плавательным пузырем, поднимающимся над поверхностью воды, от которого свешиваются вниз длиннейшие (до 30 м!) ловчие щупальца, снабженные многочисленными стрекательными клетками. Физалия очень ярко окрашена. Пузырь отливает голубым, фиолетовым и пурпурным цветом, все свешивающиеся вниз придатки ультрамариновые. За яркую окраску физалия получила также название португальского военного кораблика: в средние века португальцы любили пестро окрашивать свои военные суда. Плавающая на поверхности моря физалия вследствие яркой окраски и значительной величины видна издалека, и пловцы всегда остерегаются соприкосновения с ней, так как можно получить очень сильный «ожог», вызывающий сильнейшую боль. Пораженный физалией человек, даже если он хорошо плавает, с трудом держится на воде. Вскоре после этого может наступить и общее тяжелое заболевание с повышением температуры, длящееся несколько дней. Физалии распространены исключительно в тропических морях, и очень редко отдельные их экземпляры заносятся в более теплые участки умеренной зоны.
В последние годы в ряде медицинских и биологических журналов стали появляться сообщения о серьезных отравлениях, вызываемых крупными сцифоидными медузами хиродропус (Chirodropus) из отряда кубомедуз.
Это также обитатель тропических морей. Высота колокола медузы достигает 10—12 см, на краю зонтика сидят 4 щупальца, каждое из которых разветвлено и имеет 9—12 концов. Медуза полупрозрачна и плохо видна в воде. Конечно, ядовитые свойства этой медузы проявились не в последние годы, но прежде на нее просто не обращали внимания в связи с общей плохой изученностью кишечнополостных тропических морей.
Впервые на ядовитость этой медузы обратили внимание в годы второй мировой войны в Австралии, куда было эвакуировано значительное число европейцев. Стало известно несколько случаев загадочной гибели людей во время купания, причем на теле погибших не находили следов зубов акул или других явных признаков повреждения. Долгое время тайна оставалась неразгаданной, но потом удалось установить, что причиной гибели является медуза. Получивший «ожог» человек, едва успев крикнуть от пронизывающей боли, захлебывался и тонул. Было отмечено, что среди пострадавших или погибших были преимущественно вновь прибывшие люди. Местные жители, особенно из числа коренного населения Австралии, могли купаться безбоязненно. По-видимому, у них имелся иммунитет к яду этой медузы.
В фауне наших дальневосточных морей также имеется одна медуза, вызывающая серьезные заболевания при соприкосновении с ней. Местные жители называют эту медузу «крестовичком» за крестообразное расположение четырех темных радиальных каналов, вдоль которых тянутся четыре также темноокрашенные половые железы. Зонтик медузы прозрачный, слабого желтовато-зеленого цвета. Размеры медузы невелики: зонтик отдельных экземпляров достигает 25 мм в диаметре, но обычно они значительно меньше, всего 15—18 мм. На краю зонтика крестовичка (научное название — Gonionemus vertens) имеется до 80 щупалец, способных сильно вытягиваться и сокращаться. Щупальца густо усажены стрекательными клетками, которые располагаются поясками. Посередине длины щупальца имеется небольшая присоска, при помощи которой медуза прикрепляется к различным подводным предметам.
Крестовички обитают в Японском море и вблизи Курильских островов. Они обычно держатся на мелководье. Их излюбленные места — заросли морской травы зостеры. Здесь они плавают и повисают на травинках, прикрепившись своими присосками.Иногда они попадаются и в чистой воде, но обычно невдалеке от зарослей зостеры. Во время дождей, когда морская вода у берегов значительно опресняется, медузы погибают. В дождливые годы их почти нет, зато к концу засушливого лета крестовички появляются массами.
Хотя крестовички могут свободно плавать, обычно они предпочитают подстерегать добычу, прикрепившись к какому-либо предмету. Поэтому, когда одно из щупалец крестовичка случайно коснется тела купающегося человека, медуза устремляется в этом направлении и пытается прикрепиться при помощи присосок и стрекательных капсул. В этот момент купальщик ощущает сильный «ожог», через несколько минут кожа на месте прикосновения щупальца краснеет, покрывается волдырями. Почувствовав «ожог», нужно немедленно выходить из воды. Уже через 10—30 минут наступает общая слабость, появляются боли в пояснице, затрудняется дыхание, немеют руки и ноги. Хорошо, если берег близко, иначе можно и утонуть. Пораженного нужно удобно положить и немедленно вызвать врача. Для лечения применяют подкожные инъекции адреналина и эфедрина; в наиболее тяжелых случаях применяется искусственное дыхание. Болезнь длится 4—5 суток, но и после этого срока люди, пораженные маленькой медузой, долго еще не могут окончательно поправиться.
Особенно опасны повторные ожоги. Установлено, что яд крестовичка не только не вырабатывает иммунитета, а, напротив, делает организм повышенно чувствительным даже к небольшим дозам того же яда. Это явление известно в медицине под названием анафилоксии.
Уберечься от крестовичка довольно трудно. В местах, где обычно купается много людей, для борьбы с крестовичком выкашивают зостеру, огораживают купальни мелкоячеистой сеткой, вылавливают крестовичков специальными сетями.
Интересно отметить, что такими ядовитыми свойствами обладают крестовички, обитающие только в бассейне Тихого океана. Очень близкая форма, относящаяся к тому же виду, но к другому подвиду, живущая на американском и европейском побережьях Атлантического океана, совершенно безвредна.
Наряду с животными, которые легко поражаются стрекательными клетками, имеются такие, на которых яд кишечнополостных совершенно не действует. Обычно эти животные поселяются в сообществе с кишечнополостными, которые служат для них надежной защитой, иногда же подобное сожительство взаимно выгодно для обоих.
ЖИЗНЕННЫЙ ЦИКЛ КИШЕЧНОПОЛОСТНЫХ, ЧЕРЕДОВАНИЕ ПОКОЛЕНИЙ
Из современных нам кишечнополостных животных наиболее просто завершается жизненный цикл коралловых полипов. Оплодотворенное яйцо начинает дробиться. Сначала оно делится надвое, затем каждая из образовавшихся клеточек в свою очередь также разделяется, и так далее. В результате образуется большое количество мелких клеток, расположенных в один слой и имеющих вид маленького полого шарика. Вслед за этим часть клеток начинает погружаться внутрь, в результате чего получается двуслойный зародыш. Из его внутреннего слоя впоследствии формируется энтодерма, а из наружного — эктодерма будущего полипа. Эктодерма покрыта многочисленными мелкими ресничками, при помощи которых зародыш получает способность плавать; с этого момента он превращается в личинку, называемую планулой. Планула не способна питаться и размножаться. Она некоторое время плавает в толще воды, затем садится на дно, прикрепляясь к нему передним концом. Вскоре после этого на заднем (теперь уже верхнем) конце планулы прорывается ротовое отверстие и образуется венчик щупалец. Так возникает первый полип. У колониальных форм этот полип вскоре выпочковывает на себе других полипов, те в свою очередь следующих и т. д. Возникает колония. По достижении колонией определенной степени развития составляющие ее полипы начинают размножаться также и половым путем, образуя яйца. На этом цикл замыкается.
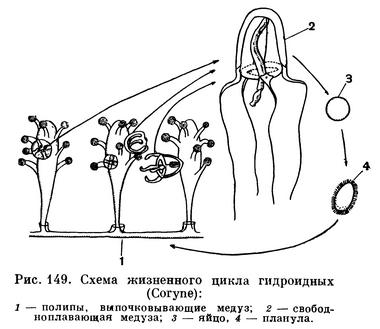
Иначе протекает жизненный цикл у гидроидных и сцифоидных. Дробление яйца и развитие планулы происходит у них так же, как у коралловых полипов, аналогичным способом возникает и первый полип — основатель колонии, а потом и вся колония (у сцифоидных колонии обычно не образуется). Однако полипы гидроидных и сцифоидных совершенно лишены способности к половому размножению, зато они выпочковывают или отделяют не только подобных себе полипов, но совершенно не похожих на них медуз. Медузы и являются половыми особями. Образование медуз у гидроидных и сцифоидных происходит по-разному. У гидрополипов медузы возникают из почек. Вначале такая почка внешне похожа на обычную почку, из которой развивается полип, но затем появляются отличия. Медузоидная почка сильно увеличивается в размере, становится полупрозрачной, внутри ее появляются 4 радиально расположенных канала и отходящий от места их скрещивания ротовой хоботок. Теперь молодая медузка внешне напоминает маленький колокольчик или зонтик. Вскоре она отрывается от колонии полипов и уплывает. Медузы самостоятельно питаются и увеличиваются в размерах, затем у них возникают половые железы. Яйца и сперма выбрасываются прямо в морскую воду, где происходит оплодотворение яиц. Из яиц возникает личинка — планула. На рисунке 149 показан типичный жизненный цикл на примере гидроида из рода Корине (Coryne sarsii).

У сцифоидных, как правило, полипы (их называют сцифистомами) не образуют колоний: возникшие в результате почкования полипчики отделяются от материнской сцифистомы и ведут самостоятельный образ жизни. Вскоре каждый такой полип претерпевает серьезные изменения: его щупальца укорачиваются, а на теле появляется ряд кольцевидных перетяжек, которые становятся все более и более глубокими, как это показано на рисунке 150. Затем начиная с верхнего конца от полипа отделяются дисковидные медузы (на этой стадии их называют эфирами), которые переворачиваются ротовой стороной вниз и уплывают. Эфиры питаются, растут и превращаются в крупных половозрелых сцифоидных медуз, размножающихся исключительно половым путем.
Таким образом, на примере гидроидных и сцифоидных мы имеем возможность познакомиться с весьма своеобразным явлением, называемым чередованием поколений. Действительно, в жизненном цикле этих двух групп кишечнополостных наблюдается правильная смена донного, сидячего, часто колониального полипоидного поколения, размножающегося вегетативно, с поколением свободноплавающим, одиночным, медузоидным, размножающимся половым путем. Разные поколения ведут различный образ жизни и устроены неодинаково, поэтому до открытия явления чередования поколений их считали самостоятельными группами животных.
Чередование поколений имеет очень большое значение для жизни гидроидных и сцифоидных: прикрепленный образ жизни полипов препятствует их расселению, а медузы, покинув колонию, могут далеко уноситься морскими течениями и разбрасывать яйца на очень большом пространстве, заселяя все новые и новые области. Чередование различно устроенных поколений довольно широко встречается в животном мире. Кроме кишечнополостных, оно обнаружено у некоторых плоских, круглых и кольчатых червей, у ряда ракообразных и даже у низших хордовых.
Именно у представителей хордовых — сальп — известный зоолог и поэт Шамиссо в 1819 г. открыл чередование поколений. Это открытие повлекло за собой поиски новых случаев чередования поколений. В 1841 г. М. Сарс описал чередование поколений у двух сцифоидных медуз — аурелии (Aurelia) и цианеи (Cyanea), а годом позже Стенструп установил, что гидроидные также развиваются с чередованием поколений.
Исследования Шамиссо, Сарса и Стенструпа явились значительным событием для зоологов прошлого века, благодаря им пришлось пересмотреть многие старые представления о биологии и систематике.
Описанный выше способ чередования поколений у кишечнополостных называют метагенезом, а животных с этим типом жизненного цикла — метагенетическими. Для метагенеза характерно правильное чередование полового поколения с поколением, размножающимся вегетативно, путем деления или почкования.
Не следует думать, что жизненный цикл у всех видов гидроидных и сцифоидных идет строго по той схеме, с которой мы уже ознакомились. В большинстве случаев от такого типичного проявления метагенеза имеются более или менее заметные отклонения, зависящие от целого ряда причин.
Главной причиной, вызывающей нарушение правильного метагенеза, следует считать различие условий среды, в которых обитают полипы и медузы. В самом деле, придонный, неподвижный образ жизни полипов неизбежно вызывает у них появление целого ряда приспособлений, отвечающих условиям их существования. Так как полипы неподвижно сидят на дне, они не нуждаются в особых органах чувств, позволяющих им на расстоянии узнавать о приближении врага или добычи: ни приблизиться к добыче, ни уйти от врагов они все равно не могут. Поэтому полипы сохранили одно только чувство осязания, позволяющее им ощупью ловить добычу, прикоснувшуюся к расставленным щупальцам. Другой способ защиты полипов заключается в образовании наружного скелета. Чем прочнее и тяжелее скелет, тем надежнее он защищает полипов и лучше удерживает всю колонию на морском дне. Указанные выше особенности полипов характерны не для одних только кишечнополостных — они вырабатываются у многих прикрепленных водных организмов. Общим свойством большинства сидячих животных является также образование колоний. Очень характерно оно и для полипов кишечнополостных и для сифонофор. С образованием колоний часто связано разделение функций ее членами. Особенно ярко это проявляется у гидроидных полипов. Только у очень немногих видов все полипы устроены одинаково и каждый из них может отпочковывать медуз. Гораздо чаще одни полипы, снабженные щупальцами и ртом, ловят для колонии добычу, но не способны размножаться, тогда как другие лишены щупалец и рта, зато приспособлены исключительно к почкованию медуз. Так дело обстоит у широко распространенного гидроида обелии (Obelia geniculata). У таких видов правильность метагенеза нарушается, так как часть полипов (кормящие особи) не дает полового потомства.
Чем сложнее становится колония, тем большее значение приобретает она в жизненном цикле. Роль медуз при этом уменьшается. Большая колония способна прокормить не только полипов, но и медуз. Поэтому медузы долгое время не отрываются от колонии и покидают ее только тогда, когда их половые продукты созревают. Наконец, имеется много таких видов гидроидных, колония которых настолько усложнена, что медузы вообще прекращают отрываться и становятся такими же прикрепленными организмами, как полипы. При этом характерные признаки медузы — зонтик, глазки, каналы гастроваскулярной системы — постепенно утрачиваются и медуза становится похожей на набитый яйцами мешочек. У пресноводных гидр медузы вовсе не образуются.
Но вернемся к типичному метагенезу и проследим за свободноплавающими медузами. Их образ жизни резко отличается от того, который ведут полипы. Медузы — подвижные животные, и потому у них вырабатываются приспособления для движения и ориентировки в пространстве. В первую очередь это касается развития зонтика и мускулатуры, а также органов чувств. У отдельных видов эти приспособления развиты настолько хорошо, что медузы могут далеко уплывать от тех мест, где они начали свою самостоятельную жизнь, отделившись от полипа. Часто они попадают в такие места, где нет подходящих условий для развития полипоидного поколения, например в открытое море, над большими глубинами. Планулы таких медуз, не найдя подходящего места для прикрепления, погибают. Но так обстоит дело далеко не со всеми видами гидроидных и сцифоидных медуз. Часть из них приспособилась к жизни именно в поверхностных слоях воды в открытом море, но при этом изменился и их жизненный цикл.
У гидромедузы бугенвиллии (Bougainvillia platygaster) яйца не выбрасываются в воду, а развиваются в половой железе самки, здесь же происходит и образование маленьких полипчиков, которые выпочковывают новых медуз. Случай этот не единственный, развитие полипов прямо на половой железе медузы характерно также и для одного вида кампанулярии (Campanularia maccrady), и для некоторых других медуз. У этих видов полипоидное поколение хотя и имеется, но оно перестало быть донным и не образует больших колоний. У некоторых видов подавление полипоидной стадии пошло еще дальше и полипы вовсе исчезли из жизненного цикла. Из планулы таких медуз возникают не полипы, а такие же медузы. Так протекает жизненный цикл большой группы гидроидных медуз — трахилид, а из сцифоидных у видов рода пелагия (Pelagia), живущих в открытом море.
Как видно из приведенных примеров, метагенетический жизненный цикл весьма пластичен. Пока он является выгодным для процветания вида, он сохраняется, но как только условия меняются и одно из поколений получает некоторое преимущество, другое поколение подавляется и может совсем исчезнуть.
ОБРАЗОВАНИЕ И СТРОЕНИЕ КОЛОНИЙ КИШЕЧНОПОЛОСТНЫХ
У многих беспозвоночных животных существует особый тип размножения, при котором на поверхности тела материнского организма образуется бугорок в виде разросшейся группы клеток. Этот бугорок увеличивается в размерах и изменяет свою форму до тех пор, пока не станет уменьшенной копией материнского организма. Этот тип размножения называется почкованием. Почки могут образовываться как на внутренней, так и на внешней поверхности тела. Внешнее почкование гораздо шире распространено в природе и служит весьма обычным способом размножения у кишечнополостных. У одиночных полипов почка, достигшая определенной стадии развития, отшнуровывается и переходит к самостоятельному существованию. Именно так размножается обычная пресноводная гидра. На мешковидно-цилиндрическом теле гидры образуется вырост, в образовании которого принимает участие как эктодермальный, так и энтодермальный слой. Вырост, увеличиваясь в размерах, принимает цилиндрическую форму, на его верхнем конце образуется венчик щупалец и прорывается ротовое отверстие. Таким образом, на теле гидры образуется дочерняя гидра, уже способная к самостоятельному образу жизни, но обладающая несколько меньшими размерами. Часто, однако, на теле такой дочерней гидры, еще не успевшей отделиться от материнской, в свою очередь образуется новая почка. Процесс почкования может пойти еще дальше, и тогда возникает уже довольно сложный комплекс из нескольких соединенных между собой гидр с общей для всех них гастральной полостью.
Описанный комплекс гидр обладает большой целостностью и физиологической самостоятельностью. С другой стороны, это сложное целое заведомо делимо, ибо каждая из молодых гидр представляет собой потенциально самостоятельный организм. И действительно, когда наступает какой-то момент, отдельные полипы начинают один за другим отшнуровываться и переходить к самостоятельному образу жизни. Рассмотренный комплекс называют временной колонией.
Гораздо чаще, однако, у кишечнополостных наблюдается образование настоящих, или постоянных, колоний. Колонии кишечнополостных удивительно разнообразны. Среди них могут быть стелющиеся и древовидные, перистые и в форме ершика, изящные и тонкие колонии и колонии с массивным известковым скелетом. Разнообразна и окраска колоний.
Форма колоний зависит прежде всего от характера скелета и степени интеграции колонии. Все полипоидные колонии (за исключением морских перьев) прочно прикрепляются к грунту или к каким-нибудь твердым предметам, лежащим на грунте. Прикрепляются колонии при помощи расширенного основания. У гидроидов таким основанием служит переплетение ползучих нитей, или гидроризы. Оно образуется сразу после оседания личинки гидроида на грунт или иной субстрат. От гидроризы отпочковываются первые полипы колонии. У коралловых полипов образование специальных нитей основания не наблюдается. Колонии их прикрепляются при помощи мясистой мягкой подошвы или при помощи расширенной известковой или роговой базальной пластинки. Среди глубоководных шипастых кораллов (Antipatharia) имеется широко распространенный в океане вид Bathypathes patula, представленный двумя экологическими формами. У одной из них основание в виде базальной пластинки, основание другой крючкообразно загнуто. По этому признаку обе формы раньше рассматривались даже как два самостоятельных вида. Образование той или иной формы основания этих кораллов определяется тем, на какой субстрат осела их личинка. Большую изменчивость формы основания колоний можно найти и среди других кишечнополостных. Например, мягкий коралл Eunephthya, обычно поселяющийся на камнях и скалах мелководных горизонтов северных морей, имеет мясистую широкую подошву. У того же коралла, развивающегося на жидких илистых грунтах, основание разрастается в несколько пузырчатых полостей, захватывающих порции грунта. Эти полости похожи на заполненные песком мешочки и служат колонии тем балластом, который помогает ей сохранять вертикальное положение (рис. 151).

Прикрепительная пластинка не образуется только у представителей отряда морских перьев. У этих животных на нижнем конце стебля образуется мускулистое расширение, при помощи которого колония внедряется в грунт. Морские перья — единственные колониальные коралловые полипы, могущие самостоятельно передвигаться с одного места на другое. Движение по грунту осуществляется волнообразными сокращениями расширенного основания колонии.
Наиболее просто устроены стелющиеся и коркообразные колонии. В таких колониях отдельные полипы отходят непосредственно от сетчатой, нитевидной гидроризы (у гидроидов) или от мембрановидной мягкой пластинки, покрывающей субстрат (у мягких восьмилучевых кораллов или зоантарий). В этих случаях ни нити гидроризы, ни мембрановидная пластинка не принадлежат какому-нибудь одному полипу, а представляют собой общее тело всей колонии — ценосарк. Наиболее яркими примерами таких колоний являются гидроиды Hydractinia или Perigonimus и мягкие восьмилучевые кораллы Clavularia. Гораздо чаще, однако, кишечнополостные образуют древовидные колонии. Тогда от базальной пластинки или гидроризы отходит несущий ветви ствол, состоящий из одной или нескольких соединенных между собой трубок.
Тип ветвления, а следовательно, и общая форма колонии зависят от места расположения почек или зон роста. У некоторых гидроидов, чаще всего у тех, полипы которых не защищены чашечкой, и у морских перьев центральный ствол колонии образуется путем увеличения в длину ножки самого первого после оседания личинки полипа. Он остается всегда на вершине колонии. Непосредственно под ним локализуется зона роста и почкования новых, вторичных, полипов. В свою очередь на ножках вторичных полипов, непосредственно под ними также имеется зона роста и почкования. За счет этих зон образуются ветви колоний и покрывающие их полипы (рис. 152). Ветвление таких моноподиальных, как их называют, колоний может быть в простейших случаях беспорядочным, тогда ветви и полипы отходят от ствола во все стороны.
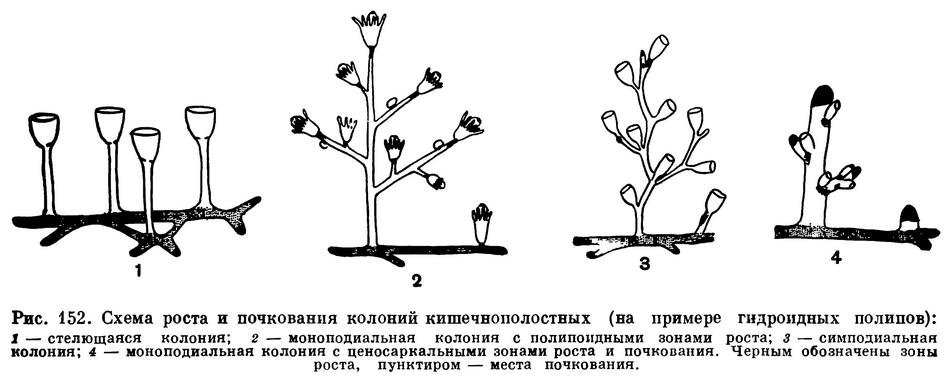
Так ветвятся колонии многих гидроидов и наиболее просто организованные колонии морских перьев. У более сложных колоний морских перьев вторичные полипы отходят от ствола в одной или нескольких плоскостях и на равном друг от друга расстоянии или от так называемых «листьев», т. е. боковых выростов, также расположенных правильными рядами. Такое правильное расположение вторичных полипов и «листьев» колоний обеспечивается тем, что почкование происходит только в одной плоскости.
Некоторые более высокоорганизованные гидроиды и коралловые полипы обладают другим типом ветвления колоний. Зона почкования у них также расположена ниже чашечки полипа, но ножка последнего имеет ограниченный рост. Поэтому, после того как первый полип сформировался, под его основанием начинает образовываться ножка следующего полипа, растущего вверх и в сторону. Иногда в зоне роста одновременно возникает несколько ножек, дающих начало нескольким веточкам. При таком типе ветвления, называемом симподиальным, наиболее старые полипы остаются у основания колоний и их ветвей. Для симподиальных колоний характерен также постоянный перелом продольной оси ствола и ветвей в местах отхождения новых полипов (рис. 152). Так растут и ветвятся многие гидроиды, полипы которых защищены чашечками, и некоторые коралловые полипы. Но высшим типом ветвления колоний гидроидов и коралловых полипов является такой, при котором зона роста и почкования находится не ниже первичного полипа, а на вершине выроста гидроризы или базальной мембраны. В этом случае функция роста и почкования переходит к общему телу колонии, или ценосарку. Основной ствол растет вверх, постепенно отпочковывая от себя полипов или образуя новые ценосаркальные выросты, боковые ветви. Для колоний гидроидов с ценосаркальным ростом очень характерно то, что отдельные полипы лишены ножек и в большей или меньшей степени погружены в толщу ветвей колонии. Ценосаркальный рост колоний мы встречаем, кроме гидроидов, у большинства коралловых полипов. У более высокоорганизованных колоний ветви и полипы, как правило, упорядочены в своем расположении, у некоторых гидроидов ветви расположены группами, в пределах которых они расходятся радиально во все стороны и лежат в одной плоскости, перпендикулярной стволу. Такие колонии называются мутовчатыми. Весьма обычен тип ветвления, при котором боковые веточки отходят от противоположных сторон ствола. При этом основания ветвей могут находиться точно друг против друга, или ветви одного ряда могут быть смещены вверх от оснований ветвей противоположного ряда. У гидроидов в таком случае образуются изящные перистые колонии, а у некоторых горгониевых кораллов и антипатарий, обладающих более крепким известковым или роговым скелетом, вееровидные и пластинчатые колонии. При этом соседние веточки часто сливаются между собой, и тогда эти обычно очень ярко окрашенные вееровидные колонии принимают форму решетчатой пластинки. Иногда такие пластинки волнообразно изгибаются, напоминая складки занавеса. Встречаются колонии, изогнутые пластинки которых образуют еще побочные пластинки. Среди восьмилучевых кораллорых полипов очень любопытные колонии образует так называемый коралл органчик (Tubipora). Скелет органчика состоит из длинных, поднимающихся от базальной пластинки известковых трубок, образованных из слившихся известковых скелетных телец. Эти трубки, напоминающие трубки органа, кроме того, соединены между собой на разных уровнях поперечными пластинками. И трубки и соединяющие их пластинки окрашены в красивый малиново-красный цвет.
При сильном развитии известкового скелета отдельные ветки сливаются друг с другом, и вся колония тогда приобретает форму массового монолита. Такие колонии образуются среди группы гидрокораллов, у которых обычный хитиноидный скелет пропитан известью и значительно утолщен, и у шестилучевых рифообразующих кораллов. Мощные коралловые полипняки образуют рифы и даже целые коралловые острова. Недаром же всего 200 лет назад считалось, что коралловые рифы принадлежат к неорганической природе, представляя собой особые минеральные конкреции! У наиболее массивных мадрепоровых кораллов скелетные чашечки полипов, сближаясь, сливаются между собой, и тогда на поверхности полипняка возникают причудливо извивающиеся линии с красивой поперечной исчерченностью.
Строение полипов и характер расположения их на ветвях колоний тесно связаны с формой самой колонии, а также с питанием. Все кишечнополостные — плотоядные животные. Они питаются планктонными организмами, мелкими беспозвоночными, ведущими придонный образ жизни, взвешенными в воде остатками животных и даже мелкими рыбками и их личинками. Большинство колониальных кишечнополостных питается мелкими планктонными животными. У наиболее примитивных колоний (в виде стелющейся на грунте пластинки) полипы приобретают вытянутую цилиндрическую форму или высоко поднимаются на тонкой ножке. Такое приподнятое над пластинкой положение полипов облегчает им ловлю добычи. С другой стороны, длинная ножка или вытянутая форма полипа делает их более уязвимыми для тех хищников, которые ими могут питаться. Поэтому эволюция колониальных форм шла в направлении приобретения древовидных стволов и одновременно сокращения длины ножки. Тем не менее еще у многих более просто организованных гидроидов с поднимающимися вверх стволами сохраняются тонкие длинные ножки полипов. Многие стелющиеся колонии находят более удобный способ для успешного облова придонных вод, поселяясь на стволах других, более высокоорганизованных колоний кишечнополостных. При этом необходимость образования ножек или собственных прямостоящих стволов у таких кишечнополостных отпадает, а отдаление от грунта осуществляется благодаря использованию стволов и ветвей других колоний. Колонии-эпифиты встречаются среди гидроидов и зоантарий. У наиболее высокоорганизованных гидроидов, образующих крепкие древовидные или простые колонии, ножки полипов исчезают полностью, а чашечки полипов или прирастают одной стороной к стволу, или же полностью погружаются в толщу ветви или ствола. Полипы восьмилучевых кораллов или способны втягиваться в тело колонии, или защищены специальными скелетными образованиями.
Успешность облова водного пространства полипами зависит не только от того, насколько высоко они приподняты над грунтом, но и от общей формы колонии. Представим себе густо и беспорядочно разросшийся куст гидроида и сравним положение полипов, сидящих на концах ветвей и в самой их чаще. Естественно, что те полипы, которые расположены на концах ветвей, находятся в более выгодных условиях и скорее могут поймать добычу, чем те, которые расположены в чаще куста. Поэтому дальнейшее упорядочение ветвей, о котором мы уже говорили, способствует лучшему питанию колоний. Но одного этого тоже недостаточно. Упорядочение ветвей коррелятивно связано с упорядочением положения на них отдельных полипов. Если полипы будут сидеть слишком тесно, они будут мешать друг другу при ловле добычи. Гораздо успешнее должен проходить лов добычи при расположении полипов в продольных рядах и в шахматном порядке. И действительно, большинство колоний кишечнополостных имеет двухрядное положение полипов. У многих гидроидов, кроме того, часто встречается такое положение полипов, при котором отверстия их чашечек попеременно направлены в разные стороны. Путем удвоения рядов могли возникнуть колонии с четырьмя и более рядами полипов. В наиболее высокоорганизованных колониях наблюдается многорядное положение полипов. В таких рядах полипы располагаются в шахматном порядке. Однако известны случаи, когда расположение полипов в один ряд дает колонии значительно большие преимущества, чем многорядное. В частности, такое положение полипов удалось обнаружить на глубоководных колониях антипатарии (Bathypathes lyra). Строение ее будет описано в соответствующем разделе. Среди колоний коралловых полипов мы находим и такие, полипы которых расположены в полном беспорядке. Однако в этом случае наблюдается отчетливая разноэтажность венчиков щупалец, обеспеченная разной длиной ловчих полипов. При этом ни один из полипов, даже полностью расправивший свои щупальца, не мешает своему соседу — в одном и том же этаже полипы сидят на достаточном расстоянии друг от друга.
Итак, исследование строения колоний гидроидных и коралловых полипов показывает, что в природе отчетливо выражено стремление к упорядочению ветвей и полипов и увеличению их числа. Для некоторых групп кишечнополостных обнаружена определенная связь между числом особей в колонии и размерами отдельных полипов. Как правило, чем больше особей в колонии, тем меньше их размеры.
Колонии одного и того же вида, обитающие в полярных районах, обладают меньшим числом ветвей и полипов, чем колонии, живущие в теплых водах. Размер полипов у первых заметно крупнее, чем у вторых. Особенно отчетливо протекает увеличение размеров у видов, завоевывающих большие глубины океана. Используя многие роды морских перьев, имеющих своих представителей как в мелководной, так и глубоководных зонах океанов, можно построить яркие морфологические ряды, иллюстрирующие этот процесс. Так, например, у преимущественно мелководного вида из рода Kophobelemnon — Kophobelemnon stelliberum, высота колоний которого достигает 300—350 мм, на стволе можно насчитать несколько десятков полипов. В расправленном состоянии они достигают длины 6—10 мм. У близкого, но более глубоководного вида Kophobelemnon polyflorum, обитающего на глубине 1000—3500 м, колонии достигают 140 мм в высоту, количество же полипов при этом сокращается в десятки раз. Чаще всего на стволе этих кодоний образуется всего лишь три полипа, длина которых в расправленном состоянии достигает 22 мм. Наконец, экспедицией на судне «Витязь» во многих местах Тихого океана и в Беринговом море ка глубинах от 2843 до 4070 м были найдены многочисленные колонии дотоле неизвестного вида Kophobelemnon biflorum, на стволе которого имелось только два очень крупных полипа. Высота этих колоний достигала 150 мм, а длина полипов в расправленном состоянии — 33 мм. Два года назад, во время плавания «Витязя» в Индийском океане, удалось поднять с глубины 3500 и 4800 м две очень интересные колонии умбеллул, также оказавшиеся представителями нового вида. На вершине ствола этих умбеллул имелся всего лишь один полип, но огромных размеров. В расправленном виде длина его достигает 125 мм! У близких видов умбеллул — Umbellula thomsoni и Umbellula dutissima, обитающих на глубине 1300—5600 и 1000—3000 м, имеется соответственно 4—7 и 3—14 полипов гораздо более мелких размеров. Таким образом, новый вид умбеллул, названный Umbellula monocephalus (одноглавая), представляет собой крайний пример этого ряда. Процесс сокращения количества полипов при одновременном возрастании их размеров прослеживается и на представителях одного и того же вида, обитающего на разных глубинах океана. Различие в строении колоний, числе и размерах полипов отдельных форм, обитающих на мелководье и на больших глубинах океана, бывает настолько велико, что такие сильно отличающиеся друг от друга формы рассматривались многими учеными как самостоятельные виды.
До сих пор говорилось о колониях с точки зрения их общего плана строения. Были рассмотрены способы укрепления колоний и их ветвление, порядок расположения. Теперь нужно остановиться на еще одном очень важном свойстве колоний кишечнополостных. У этих своеобразных животных полипы далеко не всегда бывают одного и того же строения. Очень часто отдельные полипы имеют различную форму и играют различную роль в жизни колонии. Колонии, на которых образуются разные по строению и функциям полипы, называют полиморфными.
Чаще всего полиморфизм наблюдается среди гидроидов. В силу весьма характерного для этих кишечнополостных чередования поколений, или метагенеза, на колониях, кроме питающих полипов (о которых говорили выше), отпочковываются и особи медузоидного поколения. Полипы, которые приспособлены для отпочковывания медуз, значительно отличаются от обычных питающих колонию полипов. Они лишены щупалец и ротового отверстия. Но полиморфизм может идти гораздо дальше. Тогда на ценосарке образуются чрезвычайно сильно видоизмененные полипы, несущие защитную функцию. Они имеют форму простых или разветвленных нитей, также без рта и щупалец. Но у них особое развитие приобретают стрекательные капсулы, с помощью которых полипы могут поражать врага или добычу. Такие полипы, называемые дактилозоидами, располагаются в определенном порядке вокруг питающих полипов и полипов, производящих медуз.
Полиморфизм свойствен и многим восьмилучевым кораллам. Но здесь он имеет ограниченный размах, поскольку функция полового размножения переходит к обычным кормящим полипам, или аутозоидам, амедузоидное поколение вообще отсутствует. В отношении коралловых полипов правильнее говорить не о их полиморфизме, а о диморфизме, так как на большинстве их колоний образуются полипы лишь двух типов: упомянутые выше аутозоиды и очень мелкие полипы, часто совсем лишенные щупалец, носящие название сифонозоидов. У этих зооидов перегородки в кишечной полости, столь характерные для коралловых полипов, значительно редуцированы, зато сифоноглиф развит очень сильно. Сифонозоиды не могут добывать пищу и переваривать ее. Функция сифонозоидов заключается в том, что они при помощи биения ресничек сифоноглифов создают мощный ток воды в системе каналов, связывающих между собой отдельные части колонии. С этим током воды по колонии разносятся питательные вещества и необходимый для дыхания кислород. Таким образом, мы видим, что у восьмилучевых кораллов полиморфизм полипов выражен довольно слабо. У колониальных шестилучевых кораллов полиморфизм и вовсе не наблюдается.
Высшим типом полиморфных колоний являются не гидроиды и коралловые полипы, а очень своеобразная группа свободноплавающих кишечнополостных — сифонофоры. Эти колониальные животные, которые чрезвычайно разнообразны по своему строению, окраске и внешнему виду, состоят из полого ствола, на котором выпочковываются многочисленные полипоидные и медузоидные особи, выполняющие различные функции и отличающиеся друг от друга своей формой и строением.
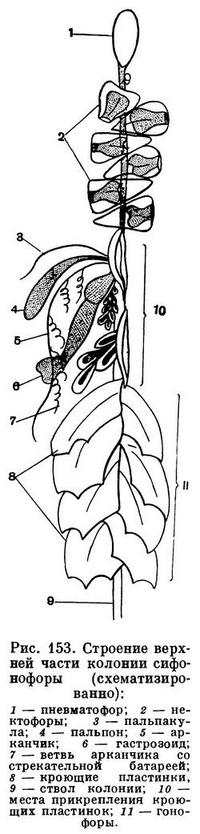
В самой верхней части колонии выпочковывается медузоидная особь — пневматофор, или плавательный пузырь. Он может быть очень маленьким (не более 1 — 20 мм), но иногда достигает 20—30 см. У одних сифонофор пневматофор замкнут, у других он имеет пору. Форма пневматофора варьирует от почти шаровидной, овальной или грушевидной до асимметричной. Иногда на поверхности пневматофора образуются различные выросты, или гребни. Пневматофор служит для большинства колоний сифонофор гидростатическим аппаратом. Животное может регулировать количество газа, содержащегося в пневматофоре, выпуская его через пору, снабженную замыкательной мышцей, либо вновь вырабатывая особыми железистыми клетками. Увеличивая или уменьшая объем пневматофора, сифонофора способна подниматься к водной поверхности либо опускаться в глубь океана. Для некоторых сифонофор, обитающих на поверхности моря, пневматофор служит парусом.
Сразу же под пневматофором у большинства сифонофор выпочковываются другие медузоидные особи — нектофоры, или плавательные колокола (в количестве от 10 до 400 в каждой колонии). У некоторых видов пневматофор отсутствует, и тогда нектофоры располагаются в самой верхней части колонии. Нектофоры, последовательно сокращаясь и расширяясь, передвигают колонию в горизонтальном направлении, но могут, так же как пневматофор, способствовать перемещению сифонофоры вверх или вниз.
В нижней половине ствола размещаются остальные особи колонии. Среди них можно различить питающие полипы, или гастрозоиды, у основания каждого из которых имеется лишь одно длинное щупальце, называемое арканчиком. Арканчик, как правило, ветвится, и на его ветвях находятся скопления стрекательных клеток. Если ветви отсутствуют, то стрекательные клетки располагаются в виде бляшек на самом арканчике.
Кроме гастрозоидов, у многих сифонофор имеются полипы другого сорта — пальпоны и цистозоиды. Они устроены проще гастрозоидов и иногда даже лишены рта. Их щупальца не ветвятся, но, так же как арканчики, снабжены стрекательными клетками. Предполагается, что пальпоны и цистозоиды выполняют чувствительные и выделительные функции.
У большинства колоний сифонофор имеются медузоидные особи, играющие защитную роль. Это кроющие пластинки. Они очень сильно изменены и совсем не похожи на медуз.
Наконец, постоянной составной частью колонии являются особи, несущие функцию полового размножения. Они могут иметь вид хорошо развитых медузок с радикальными каналами и ротовым хоботком, в стенках которого формируются половые продукты. У некоторых сифонофор такие медузоиды даже могут отрываться и свободно плавать, пока половые продукты в них не созреют. Но гораздо чаще зонтик медузоидов редуцируется и даже срастается с ротовым хоботком.
Очень важно отметить, что у наиболее сложно устроенных сифонофор часть из перечисленных особей располагается на стволе небольшими группами, называемыми кормидиями. В каждый кормидий обязательно входят полипы, выполняющие функции питания, и половые медузоиды. Кормидии могут отделяться от основной колонии и вести самостоятельную жизнь. В этом случае кормидии называют эвдоксиями или эрсеями.
Прежде отдельные кормидии считались даже самостоятельными видами сифонофор. Теперь, после того как рассмотрено строение различных колоний кишечнополостных, интересно вновь вернуться к вопросу о подчинении отдельных частей индивидуальности всей колонии, который был уже затронут в начале этого раздела.
В процессе постепенного усложнения строения колоний отдельные полипы все больше и больше теряют свое индивидуальное значение и лишь выполняют ту роль, которая диктуется всей колонией в целом. Особенно отчетливо такая подчиненность отдельных полипов проявляется в полиморфных колониях. Полипы в таких колониях уже и потенциально не способны нести самостоятельный образ жизни. В полиморфных колониях функции ее членов строго распределены между собой и вместе с тем тесно связаны. Постепенная интеграция колоний наблюдается во всех основных группах кишечнополостных. В классе гидроидных наиболее совершенными в этом отношении колониями являются сифонофоры. По сути дела, их полиморфная колония превратилась в настоящий самостоятельный организм, способный к активному направленному передвижению. Среди коралловых полипов высшей степенью подчинения отдельных членов всей колонии в целом обладают морские перья, также превращающиеся в самостоятельный прекрасно слаженный организм. И для морских перьев чрезвычайно характерна способность к самостоятельному, активному передвижению по грунту, способность укрепляться в нем. К сифонофорам и морским перьям уже вполне приложимо понятие особи, ибо эти колонии представляют собой единое морфо-физиологическое целое, в котором индивидуальность отдельных частей полностью отсутствует.
Как же осуществляется общая регуляция деятельности колоний у кишечнополостных, этих наиболее просто устроенных животных? Этот вопрос еще ждет своего решения, так как в настоящее время знания в области анатомии и физиологии кишечнополостных еще недостаточны для этого. Но многие эксперименты и наблюдения говорят о том, что общий регуляторный механизм в колониях существует и оказывается тем сложнее, чем больше степень интеграции колонии. В простейших случаях, например в колониях гидроидов, у которых нервная система находится на очень низкой стадии развития, регуляторная деятельность едва выражена. Это объясняется, видимо, тем, что отдельные полипы колоний связаны между собой ценосаркальными каналами. Во всяком случае, о наличии общеколониальной нервной системы у гидроидов никаких сведений до сих пор нет, раздражение одного из полипов колонии не передается остальным. В наиболее просто устроенных колониях коралловых полипов общеколониальная нервная система также отсутствует и передача раздражений также осуществляется только по ценосаркальным каналам. Но благодаря особенностям проводящей системы передача раздражения осуществляется на более высоком, чем у гидроидов, уровне. Например, раздражение одного из полипов колонии Alcyonium уже передается соседним полипам. Правда, волна сокращения полипов при этом распространяется удивительно медленно и, как правило, захватывает только веточки, расположенные в непосредственной близости от того полипа, к которому было приложено раздражение. Но по мере усложнения строения колоний и их интеграции усложняется и нервная система кишечнополостных. У полиморфных колоний высшего типа уже образуется общеколониальная нервная система, связывающая самые отдаленные друг от друга участки колонии. Нервная система, пронизывающая все тело колонии, наблюдается, как и следовало ожидать, у сифонофор среди гидроидных полипов и у морских перьев среди коралловых. У этих кишечнополостных животных раздражение одного полипа вызывает резкую реакцию всей колонии. Как правило, эта реакция выражается в сокращении колонии. У сифонофор быстро втягиваются щупальца, сокращается пневматофор или плавательные колокола. Организм начинает погружаться или меняет направление своего движения, стремясь уйти от опасности. Раздражение одного из многочисленных полипов морского пера в течение нескольких секунд распространяется по всей колонии. Несколькими сильными сократительными движениями базального расширения колония втягивается глубоко в грунт. Сокращается и вся верхняя часть колонии, остающаяся над грунтом. Полипы быстро исчезают в толще ствола, а по всей колонии при этом пробегают волны фосфоресцирующего света.
СОЖИТЕЛЬСТВО КИШЕЧНОПОЛОСТНЫХ С ДРУГИМИ ОРГАНИЗМАМИ
Среди кишечнополостных довольно много примеров сожительства (синойкии (Синойкия — сосуществование, от греческих слов syn — вместе, oikos — дом.) и симбиоза), тогда как число паразитов очень незначительно.
Объяснить возникновение различных форм сожительства не столь сложно. У таких организмов, как кишечнополостные, ведущих прикрепленное существование и имеющих ветвистую древовидную форму, легко возникают обрастания одних организмов другими. В море всегда можно найти высоко поднимающуюся над грунтом крепкую древовидную колонию, ветви которой покрыты разнообразными колониями гидроидов, альционарий, мшанок или одиночными организмами — актиниями, асцидиями, брахиоподами. Кишечнополостные, как колониальные, так и одиночные, часто обрастают шероховатую грубую поверхность асцидий и их длинных ножек, прикрепляются к поверхности и к отдельным длинным спикулам губок, покрывают слоевища водорослей и листья морских трав и т. д. Но большинство из них предпочитает какой-нибудь один определенный вид, который они и используют в качестве субстрата. Так, например, колонии гидроида Filellum serpens можно встретить на стволах почти всех других гидроидов, но обычно эта стелющаяся форма развивается на нескольких видах морских елочек — сертулярий. Другой гидроид — Obelia flexuosa — обычно живет на слоевищах бурой водоросли Fucus, хотя колонии обелии можно легко встретить и на других водорослях, и на листьях морской травы Zostera. Видимо, таким путем совершается в природе переход от безвыборочного обрастания к постоянным формам сосуществования (синойкии).
Гидроид Perigonimus — очень характерный сожитель некоторых двустворчатых моллюсков. Обычно этот гидроид поселяется на том краю раковины моллюска, который прикрывает его сифоны. Моллюски, служащие хозяином для Perigonimus, по типу своего питания относятся к организмам-фильтраторам, питающимся мельчайшими органическими частичками, взвешенными в морской воде. Для того чтобы обеспечить себя пищей, моллюск должен непрерывно прогонять через свою мантийную полость воду, поступающую через вводный и выходящую через выводной сифон. Создаваемые при этом токи воды, очевидно, облегчают гидроиду улавливание пищи. Другой стелющийся гидроид — Hydractinia echinata — является сожителем брюхоногих моллюсков, чаще всего хищного моллюска Natica. Гидроид Ptilocedium repens, обитающий в Тасмановом море, приспособился к существованию на морском пере Sarcophyllum. Это очень любопытный случай комменсализма, ибо морские перья крайне редко обрастают.
Синойкия имеется и среди мелких восьмилучевых кораллов — альционарий. Например, обитающий в Адриатическом море Parerythropodium coralloides обитает исключительно на ветвях живых колоний горгонарии Eunicella verrucosa. В Ла-Манше эта же горгонария опять-таки служит хозяином, но теперь уже другому комменсалу — актинии Amphianthus dohrnii.
Среди самих же горгонарий настоящей синойкии, видимо, нет, и они могут быть лишь хозяевами для других морских организмов. Так, например, два других вида горгонарий — Primnoa resedaeformis и Paragordia arborea, широко распространенные в умеренной зоне северного полушария, в Норвежском море почти всегда служат хозяевами колоний зоантарии Epizoanthus norvegicus. Желтая Primnoa является, кроме того, хозяином желтоокрашенной формы актинии Ptichodactys patula. Особая форма этой актинии, имеющая синюю окраску, поселяется на синей же горгонарии — Paramuricea placomus.
Очень много синойкий среди зоантарий колониальных шестилучевых кораллов, никогда не образующих собственного скелета, но внешняя поверхность которых бывает часто инкрустирована всевозможными посторонними частицами как органического (раковинами фораминифер и диатомовых водорослей, спикулами губок и коралловых полипов), так и неорганического происхождения (песчинками, мелкими обломками известковых пород и т. д.). Palythoa, один из видов этих кишечнополостных, является сожителем обитающей в японских водах горгонарии Corallium inutile. Коралл-хозяин служит для Palythoa не только субстратом для прикрепления и отдаления от грунта, но, что гораздо любопытнее, и поставщиком материала для строительства внешнего скелета: Palythoa инкрустирует поверхность своей колонии известковыми склеритами горгонарии. Другой вид зоантарий, распространенный в Адриатическом море, — Parazoanthus axinellae — сожитель двух видов губок из рода Axinella хотя, как исключение, и поселяющийся на некоторых других губках.
Синойкия широко распространена и среди актиний. Выше сообщалось уже о том, что желтоокрашенная и синяя формы актинии Птиходактис являются соответственно сожителями горгонарий Primnoa и Paramuricea. Примеров подобного рода можно привести очень много. Поэтому остановимся на наиболее интересных. При глубоководных тралениях, проведенных с советского исследовательского судна «Витязь» в северо-западной части Тихого океана на глубине 3—4 тыс. м, очень часто попадались лопатоногие моллюски Scaphopoda, чьи конические саблевидно изогнутые раковины удивительно напоминают по форме моржовый клык. Почти на всех раковинах собранных моллюсков были найдены бесцветные тонкостенные актинии с небольшим количеством щупалец. Актинии эти поселяются на уровне нижней трети раковины, т. е. чуть выше той части раковины, которая постоянно погружена в грунт. Можно предположить, что при движении моллюска по грунту мельчайшие организмы, живущие в верхнем слое грунта, всплывают потревоженные и становятся жертвой актинии. Сожительство этой актинии с лопатоногим моллюском настолько обычно, что случайные находки раковин, свободных от актиний, каждый раз вызывали удивление. На другом субстрате эти животные, по-видимому, никогда не поселяются.
Известны и еще более интересные случаи синойкии среди глубоководных актиний. В начале XX в. в результате глубоководных тралений в Атлантическом океане, проведенных Монакским океанографическим институтом, было поймано два экземпляра голотурии Pseudostichopus villosus. Нетрудно представить себе удивление ученых, когда они на нижней поверхности этих голотурий обнаружили укоренившихся там двух небольших плотных актиний! Однако сожительство актинии, названной Sicyopus commensalis, видимо, не случайно. На обоих экземплярах голотурий актинии поселились в одном и том же месте — спереди и немного сбоку от окруженного короткими щупальцами ротового отверстия. Эти уплощенные голотурии непрерывно ползают на поверхности грунта, собирая щупальцами частички органического детрита. Sicyopus commensalis при этом оказывается в положении авангарда, первой осваивающей новые «охотничьи угодья». Любопытно, что под снятой с тела голотурии актинией была обнаружена небольшая ямка, в которую актиния была как бы запрессована. В связи с этим интересно отметить наблюдения, проведенные над рядом других актиний, которые показали, что их подошва выделяет особый секрет, легко растворяющий тот органический по происхождению субстрат, на котором они поселяются. Так, например, актиния Cereus pedunculatus, укрепившаяся на раковине моллюска, постепенно растворяет ее непосредственно под своей подошвой и через некоторое время проваливается внутрь раковины через образовавшееся в ней отверстие. Возможно, что ямка в теле голотурии, в которой укрепляется актиния, имеет такое же происхождение. Сожитель в этом случае не очень-то щадит своего хозяина. Но еще более безжалостно обращаются со своим хозяином актинии, поселяющиеся на глубоководном морском пере Kophobelemnon biflorum. Эти морские перья были впервые и в большом количестве найдены экспедиционным судном «Витязь». По имеющимся данным, они обитают вдоль побережья северной части Тихого океана на глубине 3—4 тыс. м, предпочитая заиленные склоны глубоководных впадин. На каждом из собранных экземпляров Kophobelemnon biflorum были обнаружены одна или несколько актиний, близких к Sicyopus. Так же как и в случае сожительства с голотурией, эти актинии разрушают своей подошвой мягкие ткани морского пера, все глубже и глубже проникая в его тело. Наконец, их подошва достигает известкового осевого стержня колонии и плотно его обволакивает. Комменсализм подобного типа оказывается губительным для морского пера. Все его мягкие ткани отмирают и отваливаются. И тогда комменсал-победитель оказывается сидящим на гладком осевом стержне отмершей колонии на высоте 7—10 см над уровнем грунта. В просмотренной нами коллекции Kophobelemnon biflorum, собранной экспедицией «Витязя», можно найти все стадии описанного процесса. Имеются великолепные экземпляры пера, на котором молодые актинии только что поселились, есть и колонии, уже приходящие в упадок. На двух экземплярах Kophobelemnon biflorum от мягких его тканей сохранилась лишь самая верхняя часть с двумя крупными аутозоидами, а нижняя полностью разрушена актиниями, ставшими уже очень крупными.
Наконец, в коллекции есть и несколько стержней отмерших колоний, плотно охваченных подошвой бывших сожителей.
Не только взрослые актинии могут быть сожителями других животных. Личинки обитающих в северных морях Edwardsia, Peachia, Haloclava и некоторых других родов проникают под колокол медуз и при помощи своего рта прикрепляются к ротовому конусу, гонадам или внутренней поверхности колокола. Прикрепившись к медузе, личинки плавают вместе с ней до момента, когда наступает время превращения их во взрослые стадии. Если личинка по какой-либо причине теряет своего хозяина, она способна самостоятельно плавать на поверхности воды, пока вновь не проникнет под колокол проплывающей мимо медузы. Среди личинок актиний, кроме сожителей медуз, существуют и сожители гребневиков. Так, например, некоторые личинки в Северном море часто поселяются на гребневике Bolinopsis infundibulum. Личинки одной австралийской актинии, однако, не ограничиваются жизнью под колоколом медузы, а проникают даже в радиальные каналы. Собственно говоря, такой образ жизни скорее напоминает паразитизм, чем синойкию. Для того чтобы выйти затем на свободу, этим личинкам приходится разрушать часть колокола. Да и питание этих сожителей скорее всего происходит за счет хозяина.
Высшим типом сожительства двух организмов, несомненно, является симбиоз, приносящий обоим партнерам взаимную выгоду. Симбиоз значительно реже встречается в природе, чем синойкия, а тем более паразитизм. Справедливо это и для кишечнополостных животных. Если сожительство можно найти почти во всех систематических группах кишечнополостных, среди обитателей любых морей и любых глубин, то этого уже нельзя сказать об организмах, ведущих симбиотический образ жизни.
Так же как и синойкия, симбиоз возник из простого обрастания одних организмов другими. В случае обоюдной выгодности такого сожительства в дальнейшем мог выработаться инстинкт или определенные приспособления в строении, облегчающие встречу организмов-симбионтов. Наконец, симбиоз становится уже необходимостью для таких организмов: существование каждого из них настолько тесно зависит от существования другого, что жизнь их в отдельности оказывается уже затруднительной или невозможной.
Сказанное можно проиллюстрировать рядом примеров. Так, гидроид Hydractinia echinata, как уже было упомянуто выше, обычно обрастает раковины хищных брюхоногих моллюсков, являясь их комменсалом. Но часто Hydractinia echinata обрастает и раковины, служащие домиком ракам-отшельникам. Разрастающаяся со временем гидрориза гидроида постепенно превращается в сплошную пластинку, причем край ее при этом перерастает устье раковины и нависает над телом рака-отшельника. Такое сожительство становится выгодным как для гидроида, так и для рака-отшельника. Постоянное передвижение рака по дну водоема в поисках пищи способствует лучшему газообмену гидроида, а взмученные остатки пищи, пожираемой раком, легко улавливаются и поедаются полипами гидрактиний. Вместе с тем поросль гидроида маскирует раковину рака-отшельника, что для него, безусловно, полезно. Кроме того, раку не приходится часто менять свой домик, ибо по мере роста рака разрастается и пластинка гидроризы, служащая раку-отшельнику достаточно надежной защитой. Легко заметить, что описанное сейчас сожительство представляет собой простейший случай симбиоза. Здесь нет активной заинтересованной стороны: гидроид случайно поселяется на раковине-домике, занятой раком-отшельником. Последний же не ищет для себя раковины, уже обросшей колонией гидрактинии. Каждый из партнеров, и рак, и гидроид, может существовать вполне самостоятельно.
Классические примеры симбиоза двух организмов дает сожительство раков-отшельников и актиний. Но и в этих случаях можно различить три ступени постепенного осложнения отношений между партнерами. В простейшем варианте рак-отшельник еще не в состоянии снять поселившуюся на каком-нибудь субстрате актинию и перенести на свой домик. Он лишь бросает свой старый домик, если случайно найдет подходящую пустую раковину с уже поселившейся на ней актинией. Так ведет себя, например, рак-отшельник Eupagurus excavatus, если он случайно натолкнется на подходящую для него пустую раковину, на которой поселилась актиния Calliactis parasitica. Совершенно так же поступает рак, найдя пустую раковину с сидящей на ней Hormathia corenata или даже Actinia equina, хотя обе эти актинии ведут вполне самостоятельный образ жизни и не вступают в симбиоз с другими животными.
Совместная жизнь актинии и рака-отшельника дает несомненные преимущества каждому из партнеров. Актинии теперь не нужно находиться в постоянном и напряженном ожидании поимки пищи. Она без труда улавливает щупальцами взмученные остатки пищи рака-отшельника, в поисках которой он непрерывно движется по дну водоема. Рак же находится теперь под надежной защитой стрекательных капсул актинии — грозного оружия для большинства врагов раков-отшельников.
Более высокую ступень в развитии симбиотических взаимоотношений показывает сожительство той же Calliactis parasitica с другим раком-отшельником — Pagurus arrosor. Рак-отшельник не должен искать для себя пустую раковину с уже сидящей на ней актинией. Он способен снять ее с любого субстрата, перенести на свой домик и укрепить на нем. Когда раку-отшельнику старый домик оказывается мал и он вынужден сменить его на новый, он снимает актинию со старого и переносит на свой новый домик. Происходит это следующим образом. Своими ходильными ногами первой и третьей пар рак-отшельник начинает поглаживать и похлопывать актинию. Казалось бы, актиния, которая в ответ на любое раздражение отвечает сокращением своего тела и выбрасыванием стрекательных нитей, должна была бы поступить аналогичным образом, когда рак начинает ее похлопывать. В данном же случае актиния ведет себя не совсем так. При первом прикосновении рака она, правда, начинает сжиматься, но затем вновь распускается и стрекательных нитей не выбрасывает. Даже полностью сжавшаяся актиния начинает распускаться, если ее начинает поглаживать рак-отшельник. После того как актиния полностью распустилась, ее будущий спутник начинает поглаживать подошву актинии. Этим поглаживанием рак добивается сокращения подошвы и отделения ее от субстрата. Тогда раку остается только подставить свою раковину поближе к подошве актинии и перекатить на нее актинию.
Впрочем, имеются указания, что после того, как произошло отделение подошвы, актиния сама наклоняется в сторону раковины отшельника и охватывает ее своими щупальцами. Затем она вновь переворачивается и прикрепляется к раковине уже подошвой. В тех случаях, когда рак-отшельник находит не вполне плотно укрепившуюся актинию, ему даже не приходится ее похлопывать и поглаживать — описанный процесс пересадки актинии протекает без его помощи.
Актиния выделяет своей подошвой слизистую перепонку, которая способствует более надежному укреплению актинии на домике рака-отшельника. Однако эта тонкая мембрана не может перерасти за край раковины и тем самым увеличивать ее объем. Отшельнику приходится через некоторое время менять свой старый домик, ставший ему тесным, на новый, более просторный, и вновь пересаживать на него свою спутницу. Если на раковине остается достаточно места, предприимчивый рак может обзавестись еще одной актинией. Описаны случаи, когда Pagurus arrosor носил на своей раковине до 8 актиний.
Лучшим примером высшего типа симбиоза актинии и рака-отшельника служит сожительство Adamsia palliata с Eupagurus prideaxi. Адамсия, распространенная в Средиземном море и в умеренных водах восточной части Атлантического океана, ведет самостоятельное существование только в самом начале своей жизни. Молодые адамсии имеют обычное для актиний цилиндрическое тело с довольно широкой подошвой, при помощи которой они прикрепляются к камням или пустым раковинам моллюсков. Очень редко они успевают дорасти до 1 см в высоту: как правило, еще до этого их захватывают раки-отшельники. До сих пор еще не вполне установлено, способна ли эта актиния и во взрослом состоянии вести самостоятельный образ жизни, если она не вступает по какой-нибудь случайности в симбиоз с отшельником. Однако представить себе адамсию-одиночку очень трудно, зная повадки отшельника Eupagurus prideaxi. Среди своих сородичей он самый подвижный и драчливый. Поэтому он и предпочитает самые мелкие раковины в качестве своего домика. Раковины хищных брюхоногих моллюсков Nassa и Natica, которыми он обычно пользуется, едва прикрывают его мягкое брюшко. У взрослого, крупного рака такой домик защищает только самый задний отдел брюшка. Что же касается головогруди рака, то она никогда не прикрывается раковиной. Защитить себя от врагов и хищников Eupagurus может только одним способом — найти и вступить в симбиоз с адамсией. Молодой рак находится в постоянных поисках адамсии. В поиске актинии ему помогает не столько зрение, сколько осязание и способность улавливать определенные химические раздражения, исходящие от актинии. В аквариуме этот рак-отшельник обнаруживает молодую адамсию и в том случае, если она прикрыта колпачком из ткани. Как только он обнаруживает одиночную молодую актинию, он стремительно бросается к ней. В это время его, кажется, ничто уже не может остановить. Подойдя к адамсии, Eupagurus крепко схватывает ее своими клешнями и не отпускает в течение 10 минут. Казалось бы, что такое обращение должно заставить адамсию сжаться. На самом же деле этого не происходит. Напротив, подошва молодой адамсии начинает отделяться от субстрата. После этого рак, не разжимая клешней, переносит ее на свой домик.
Когда молодой рак-отшельник, еще не успевший обзавестись спутницей, встречается со своим более удачливым сородичем, он моментально вступает с ним в бой за обладание адамсией. Если обороняющийся оказывается слабее, он бросает раковину и пускается в бегство, оставляя агрессору и свой дом и свою актинию.
Рак отшельник Eupagurus prideaxi укрепляет адамсию на раковине так, чтобы ее ротовой диск был направлен вперед, а все ее тело находилось сзади и чуть ниже ротового аппарата самого рака. При таком положении актиния без труда может улавливать и поедать остатки пищи, пожираемой раком. Вместе с тем такое положение актинии заставляет ее со временем изменить форму тела. Дело в том, что рост ее не может идти равномерно во все стороны. Распространению подошвы по раковине дальше вниз и назад препятствует постоянное трение раковины о грунт. Не может актиния расти и вперед — этому мешают находящиеся в движении ноги рака. Актиния разрастается вбок и вверх, вдоль края устья раковины. Тело ее поэтому приобретает полулунную форму. Если рак-отшельник не обзавелся второй актинией, то боковые лопасти подошвы, поднимаясь все выше и выше по раковине, могут сомкнуться, окольцовывая устье раковины. Если на раковине находятся две актинии, то края их подошв также смыкаются друг с другом на верхней и нижней поверхностях раковины. У адамсии есть и другое свойство, крайне полезное для рака-отшельника: подошва адамсии, так же как и подошва калиактис, выделяет плотную слизистую пластинку, которая, однако, у адамсии быстро твердеет и принимает роговую структуру. Эта пластинка по мере роста актинии перерастает край устья раковины и нависает над телом рака. Объем домика тем самым все время увеличивается, и быстрорастущему раку не приходится менять своего домика. Крупные взрослые адамсии достигают в ширину 5—7 см. Домик же рака-отшельника имеет гораздо более скромные размеры. Получается, что адамсия в конце концов сидит не столько на поверхности первоначального убежища рака, сколько на роговом кольце вокруг его тела, которое сама и построила.
Такой домик очень удобен и для рака-отшельника, ибо он несоизмеримо легче толстостенной раковины моллюска и достаточно прочен. В то же время он эластичен и нисколько не затрудняет движений рака.
После укрепления адамсии на раковине меняется не только форма ее тела, но и ее окраска. Молодые адамсии окрашены в яркий розово-красный цвет. Но у взрослых адамсий, вступивших в симбиоз с пагурусом, окраска тела меняется и становится желто-розовой, различных оттенков, но такой же, как и окраска головогруди и верхних члеников ходильных ног рака. Наиболее ярко окрашены спинные лопасти подошвы актинии; ближе к бокам они светлеют. Ротовой диск и щупальца, невидимые сверху, почти совсем лишены окраски (см. цветную таблицу 9).
Взаимная выгодность симбиоза Adamsia palliata и Eupagurus prideaxi очевидна. Рак-отшельник, почти все тело которого оказывается не прикрытым раковиной, давно стал бы жертвой какого-либо хищника, если бы он не был надежно защищен актинией. Даже такие свирепые хищники, как головоногие моллюски, не смеют нападать на отшельника, боясь страшного оружия адамсии — ее стрекательных нитей. Адамсия в свою очередь не заботится о пище — это прямая обязанность ее хозяина. Постоянное передвижение рака по грунту также весьма выгодно для актинии, так как это улучшает условия газообмена. Не обязательно, однако, укрепление актинии на раковине или панцирях ракообразных. В природе вырабатываются и другие формы симбиоза между актиниями и раками. Актиния может быть использована тогда не только как оружие защиты, но и как орудие для добывания пищи. Один из средиземноморских крабов известен тем, что он постоянно держит в своих клешнях двух актиний, которые служат ему одновременно и защитой от врагов и орудием добывания пищи. Своими щупальцами, вооруженными стрекательными капсулами, актиния схватывает проплывающую добычу. Но ее хозяин-краб беззастенчиво отбирает часть добычи для себя. Единственное преимущество актинии, ведущей такой образ жизни, заключается в том, что она все время перемещается с одного места на другое. Симбиоз в данном случае носит несколько однобокий характер.
Во всех до сих пор описанных сообществах активной стороной были ракообразные и пассивной — актинии. Но бывает и наоборот. Так, например, актиния Autholoba reticulata, обитающая на западном побережье Южной Америки, сама стремится вступить в симбиоз с ракообразными, предпочитая другим краба Hepatus. Эта актиния, как и другие ее сородичи, вначале поселяется на камнях, прикрепляясь к ним своей подошвой. Но затем она переворачивается ротовым диском вниз и держится за субстрат своими щупальцами. Подошва ее отрывается от субстрата. В таком необычном для актинии положении она ожидает приближения своего будущего спутника. Через некоторое время терпение ее может быть награждено, и тогда ее подошва крепко схватывает лапку проползающего мимо краба. Затем актиния переползает на его панцирь.
Существует и не менее интересный симбиоз актинии и рыб. На мелководье западного побережья Австралии обитают самые крупные в мире актинии Stoichactis, ротовой диск которых достигает в диаметре 1,5 м. На поверхности диска несколькими концентрическими рядами располагаются бесчисленные щупальца этих актиний. Среди щупалец или в непосредственной близости от них держится парочка очень ярко окрашенных рыбок — амфиприонов. По наблюдениям ряда авторов, невозможно найти ни одной актинии, возле которой бы не держались эти рыбки, так же как и рыбок вдали от актиний. Даже в поисках добычи рыбки никогда не удаляются от актиний более чем на метр. При малейшей опасности они прячутся в лесу длинных щупалец актиний. Если в аквариум с актинией и амфиприонами посадить хищных рыб, то они могут жить вместе очень долго, но стоит убрать актинию, как амфиприоны становятся жертвой хищных рыб. Амфиприоны не умеют ни быстро плавать, ни находить защиту в водорослях, а их очень яркая окраска и красивый рисунок из белых и черных полос на красном или золотистом фоне слишком заметны для врага.
В естественных условиях эта же яркая окраска идет на пользу рыбкам, привлекая к ним добычу. Так как рыбки хватают добычу в непосредственной близи от щупалец и ротового отверстия актинии, здесь же поедают ее и, естественно, теряют ее остатки, они тем самым снабжают пищей и актинию. Кроме того, держась среди щупалец, рыбки постоянным биением своих плавников создают токи воды, улучшающие условия газообмена актинии.
Кроме этих рыбок, среди щупалец актиний часто держится несколько видов креветок и небольшой краб, подражающий в окраске рыбкам.
О симбиозе кишечнополостных с одноклеточными водорослями зооксантеллами будет сказано в главе о коралловых постройках.
Среди кишечнополостных имеется небольшое число настоящих паразитов. У гидроидных медуз купана (Cunina) и куноктанта (Cunoctantha) образовавшаяся из яйца планула оседает не на дно, а на край зонтика некоторых видов других гидроидных медуз из отряда лептолид. Здесь планула превращается в двущупальцевую личинку, которая пробирается к ротовому хоботку хозяина. Вскоре у личинки появляется вторая пара щупалец и ротовой хоботок. Придерживаясь щупальцами за нижнюю сторону зонтика, личинка запускает свой ротовой хоботок в желудок медузы-хозяина и питается за ее счет. Личинка быстро растет и, размножаясь почкованием, образует подобных себе паразитических четырехщупальцевых личинок. Вскоре внутри зонтика медузы-хозяина оказывается целая группа паразитов. После этого все личинки начинают превращаться в медуз. У них развивается зонтик, укорачивается ротовой хоботок; они покидают своего хозяина и ведут свободноплавающий образ жизни.
Еще более яркий пример паразитизма дает своеобразное кишечнополостное животное из гидроидных — полиподиум (Polypodium hydriforme).
Полиподиум — единственный представитель кишечнополостных, паразитирующий в теле позвоночного животного, он поселяется в икре некоторых осетровых рыб.
Впервые полиподиум был обнаружен академиком Ф. В. Овсянниковым в 1871 г. в икре волжской стерляди. Овсянников обратил внимание на то, что, кроме нормальных икринок, попадаются также больные, несколько более крупные и светлее окрашенные. Внутри такой икринки сидит полиподиум. Самая ранняя из известных стадий развития паразита — это маленькая двуслойная личинка, имеющая червеобразную форму. Личинки поселяются в развивающейся икринке. По мере роста икринки и накопления в ней желтка развивается и личинка; она вытягивается в длину, и на ней образуются многочисленные выросты — почки. Вскоре в каждой почке появляется по 12 щупалец, обращенных внутрь. На этой стадии паразит обладает весьма своеобразным строением — он как бы вывернут наизнанку. Его пищеварительный слой — энтодерма — расположен снаружи и непосредственно соприкасается с желтком икринки, служащим полиподиуму пищей. Покровный же слой — эктодерма — обращен внутрь.
Перед наступлением периода нереста рыбы-хозяина слои клеток у паразита принимают нормальное положение. Почки полиподиума при этом вворачиваются, захватывая внутрь гастральной полости остатки желтка из икринки. Когда рыба выметывает икру в реку, вместе со здоровыми икринками в воду попадают и зараженные. В воде паразит выходит из икринки, разорвав ее оболочку, и вскоре после этого распадается на части по числу почек — из каждой развивается двенадцатищупальцевый полип около 1 мм высотой. Из одной зараженной икринки выходит 60—90 полипов.
Первое время полипы лишены рта и питаются за счет запасов желтка, но на 4—5-й день самостоятельной жизни у них появляется рот и полипы при помощи щупалец, усаженных стрекательными клетками, начинают ловить малощетинковых червей, коловраток и других маленьких пресноводных животных. Четыре коротких щупальца полипа служат для прикрепления ко дну и для передвижения, более длинными щупальцами полип ловит добычу.
В течение всего лета полипы размножаются делением. Перед делением число щупалец у полипа удваивается, а затем на его теле появляется продольная перетяжка, и вскоре он разделяется надвое.
К осени в теле полипа развиваются половые железы. Полиподиумы раздельнополы, но изредка попадаются гермафродитные особи. Весь дальнейший цикл развития полиподиума пока еще неясен. Известно только, что мужские полипы способны оседать на молоди осетровых рыб и на их тело прикреплять свои гонады. Способы оплодотворения и пути проникновения паразита в икринку половозрелой рыбы пока совершенно непонятны.
Полиподиум приносит значительный ущерб, так как зараженная им дорогостоящая икра осетровых рыб теряет свою товарную ценность. Кроме того, паразит значительно снижает продуктивность осетровых, вследствие того что зараженные икринки погибают, не дав потомства.
Поселяется полиподиум в икре стерляди, осетра, севрюги и аральского шипа. Этот паразит был обнаружен в рыбах из Волги и ее притоков, Дона, Днепра, Днестра, Дуная, Кубани, Северной Двины, Сырдарьи и Амура.
Рыбы, зараженные полиподиумом, лечению не поддаются, но имеются широкие возможности по проведению профилактических мероприятий с целью снизить зараженность рыб в данном бассейне и препятствовать проникновению паразита в другие реки. Для этого необходимо тщательно просматривать икру при ее заготовке для засола и на рыбоводных станциях. Ни в коем случае не следует выбрасывать зараженную икру в воду, где из нее разовьются полипы, которые будут заражать здоровых рыб. При акклиматизации осетровых в новых бассейнах нельзя ввозить в них взрослых рыб, так как они могут быть заражены полиподиумом. Лучше всего для целей акклиматизации использовать тщательно просмотренную здоровую икру осетровых.
ЗНАЧЕНИЕ КИШЕЧНОПОЛОСТНЫХ В ПАЛЕОНТОЛОГИИ
Среди наиболее древних кишечнополостных преобладали организмы, лишенные хорошо развитого скелета. Известны находки нескольких сцифоидных медуз из кембрийских и меловых отложений.
Только остатки вымерших рифообразующих кораллов, обладавших массивным известковым скелетом, часто встречаются в самых различных геологических отложениях. Описано более 5000 видов ископаемых кораллов. Находили их в виде отдельных отпечатков, в виде крупных ископаемых рифов. Наиболее древними кораллами были археоциатиды из нижних кембрийских и гелиотиды из силурийских отложений.
Начиная с силурийского периода становится чрезвычайно распространенной группа очень своеобразных четырехлучевых кораллов (Tetracorallia). Это были одиночные или колониальные кораллы конической формы, с расширенной верхней чашечкой. На ее поверхности отчетливо различимы четыре первичные перегородки. Перегородки (или септы) делили чашечку на четыре сектора, каждый иа которых в свою очередь делился вторичными перегородками.
Настоящие мадрепоровые рифообразующие кораллы появились только в меловом периоде, а современные нам семейства этих кораллов — только в четвертичном периоде.
Несмотря на многочисленные и разнообразные находки ископаемых кораллов, группа кишечнополостных в целом составляла весьма небольшую часть накопленного палеонтологического материала и не могла поэтому играть сколько-нибудь существенной роли в палеонтологических реконструкциях. Однако недавние исследования, проведенные в долине Эдиакара, в Южной Австралии, заставили ученых изменить такое мнение о значении кишечнополостных в палеонтологической летописи.
Еще совсем недавно никто не знал, какими были предки тех беспозвоночных животных, которые достигли значительного развития уже в начале кембрийского периода.
И вот при раскопках докембрийских пород в долине Эдиакара удалось наконец собрать более 6000 образцов ископаемых беспозвоночных, населявших моря в протерозойскую эру. Среди собранных образцов были отпечатки медуз, принадлежавших к шести различным родам, мягких восьмилучевых кораллов и коралловых полипов, напоминавших современные морские перья, и множество других животных. Среди многочисленных отпечатков кораллов не было ни одного, который свидетельствовал бы о наличии известкового массивного скелета. Ткани древнейших коралловых полипов поддерживались лишь простыми известковыми тельцами — спикулами.
Все эти находки еще требуют пристального изучения. Однако уже сейчас можно сказать, что предположение о том, что докембрийские животные были лишены скелета или не прикрывались панцирем и потому и оставались до сих пор неизвестными, по-видимому, оказалось справедливым. Кроме того, изучение собранных остатков позволяет предположить, что отсутствие раковин или внутреннего скелета у докембрийских животных объясняется не какими-нибудь особыми условиями обитания в то удаленное от нас на сотни миллионов лет время и не каким-нибудь резким отличием образа жизни этих животных. Отсутствие скелета, видимо, должно объясняться тем, что у древнейших беспозвоночных животных еще не выработался механизм кальциевого обмена, в результате которого организмы оказались в последующем способны к образованию массового известкового скелета, панцирей и раковин.
Теперь благодаря трудам зоологов русской школы — И.И. Мечникова, В. Н. Беклемишева и А. А. Захваткина — стали понятны самые ранние периоды в эволюции кишечнополостных. Можно считать установленным, что предками современных нам кишечнополостных были примитивные многоклеточные животные, напоминавшие планул современных видов.
К сожалению, среди палеонтологических находок нет остатков этих крошечных, лишенных скелета предков многоклеточных, и поэтому представить себе картину ранней эволюции кишечнополостных можно, лишь проводя аналогии и детально изучая индивидуальное развитие.
Мы вправе предположить, что следующим за планулой этапом эволюции кишечнополостных был полип.
Для многих сидячих организмов очень характерна способность к вегетативному размножению и образованию колоний. Эта особенность позволяет им лучше бороться за существование: полипы сообща ловят и усваивают добычу, сообща защищаются от врагов.
На ранних ступенях развития кишечнополостных произошло разделение их на две крупные ветви. Появились полипы, гастральная полость которых была разделена перегородками на камеры. Это способствовало более интенсивному пищеварению. Впоследствии от этих полипов произошли коралловые полипы и сцифоидные. Те же полипчики, у которых перегородок в кишечной полости не было, дали гидроидных. Однако прикрепленный образ жизни имел и свои недостатки: потомство сидячих животных всегда поселялось рядом с родителями. Это создавало перенаселенность и препятствовало животным завоевывать новые жизненные пространства.
Появление подвижных половых особей медуз было очень существенным шагом на пути эволюции кишечнополостных, эта их особенность наряду с развитием стрекательного аппарата является одной из главнейших причин процветания типа.
Мы уже убедились, что кишечнополостные сумели приспособиться к самым различным условиям жизни в воде. Их находят среди бурных волн прибоя на берегу моря и в спокойных водах океанских глубин. Они поселяются в холодных морях Арктики и Антарктики и под палящими лучами тропического солнца. Кишечнополостные живут на дне, в толще воды и плавают на ее поверхности. Некоторые из них перешли к паразитизму, и их жизненной средой стал организм животного-хозяина. Кишечнополостные проникают даже в пресные воды, и только воздушная среда, наземное существование оказались для них совершенно недоступными.
Эта очень древняя группа организмов, несомненно, является в то же время одной из наиболее процветающих. Конечно, нельзя ожидать, что в дальнейшем из кишечнополостных возникнут какие-то высокоорганизованные животные, но их способность приспосабливаться к разнообразным условиям жизни позволит им и впредь успешно конкурировать с гораздо более высокоорганизованными обитателями моря.