(Perccottus Dybowski, 1877) Род Перккоттусы, Головешки, Genus Perccottus Dybowski, 1877 (Chinese sleepers, Amur sleepers) 1 вид
Материал подготовил Фактистов И. Е., 2022 г. Сайт www.fishbiosystem.ru
Новый род Perccottus впервые научно описал в 1877 году польский учёный Бенедикт Дыбовский (Бенедикт (Венедикт) Иванович Дыбовский, имя при рождении — Бенедикт Тадеуш Дыбовский, польск. Benedykt Tadeusz Dybowski, 1833 — 1930, польский, белорусский и российский географ, зоолог, медик, лингвист и общественный деятель, изучал медицину и естественные науки в университетах Дерпта (Тарту), Бреславля (Вроцлав) и Берлина, где и получил в 1860 году степень доктора медицины, с 1862 года экстраординарный профессор зоологии и палеонтологии в Императорском Варшавском университете, за участие в Польском восстании 1863 года был сослан в Сибирь, где исследовал озеро Байкал, реку Амур, благодаря усилиям Петербургской Академии наук в 1877 году вернулся на родину, но вскоре, желая изучить Камчатку, взял место окружного врача в Петропавловске-Камчатском, где подробно изучал природу Командорских островов, в 1883 году эмигрировал во Львов, был руководителем кафедры зоологии Львовского университета, создал зоологический музей, в 1928 году избран иностранным членом-корреспондентом Академии наук СССР за значительный вклад в изучение Сибири и Дальнего Востока России, автор описаний 34 новых видов рыб, в его честь названо 5 видов рыб).
Перккоттусы, Головешки (Perccottus) — монотипический род лучепёрых рыб из отряда окунеобразные (Perciformes Bleeker, 1859) семейства одонтобутовые (Odontobutidae Hoese & Gill, 1993), обитающих в пресных и солоноватых водах северной Азии и Европы.
Название рода Perccottus происходит от соединения названий родов Perca (Пресноводные окуни) и Cottus (Подкаменщики), ссылаясь на внешнее сходство единственного его представителя — ротана-головешки с окунями и подкаменщиками. Английские названия «Chinese sleeper» и «Amur sleeper», в дословном переводе — «китайские сони» и «амурские сони», указывают на места естественного обитания и на неторопливый и размеренный образ жизни этих рыб, которые обычно просто лежат на дне пресноводных водоёмов. Русское название «головешка» указывает на схожесть окраски и внешнего вида ротана, особенно в брачном наряде, с обгорелой деревяшкой. Иногда используется ошибочное название рода Percottus.
Найденные окаменелости (fossils) ранних одонтобутид, ближайших родственников Perccottus, были датированы средним эоценом (48,6—40,4 млн. лет назад). Обнаруженные отолиты представителей вымершего рода † Odontobutidarum Hoese & Gill, 1993, были отнесены к виду † Odontobutidarum triangularis n. sp. и датированы средним эоценом (47,8—41,3 млн. лет назад — Лютетский ярус). Местонахождение ископаемых останков: песчаный карьер Остероден (Osteroden sand pit), Нижняя Саксония (Lower Saxony), Германия (52,5° с.ш., 7,8° в.д.; палеокоординаты 49,4° с.ш., 3,8° в.д.). Окружающая среда — морская; литология — нелитифицированный мелкозернистый песчаник и конгломерат. Над базальными слоями гравия лежат мелкозернистые, рыхлые пески толщиной до 1,5 м как в песчаных карьерах Далум (Dalum), так и в Остероден, но только в последнем имеются участки, в которых арагонит не растворился и, таким образом, сохранились отолиты. [W. Schwarzhans. 2007. The otoliths from the middle Eocene of Osteroden near Bramsche, north-western Germany. Neues Jahrbuch für Geologie und Paläontologie 244(3):299-369]
Более поздние найденные отолиты представителя рода Micropercops Fowler & Bean, 1920, были датированы поздним олигоценом (27,82—23,03 млн лет назад — Хаттский ярус, Duntroonian — 27,3—25,2 млн лет назад) и отнесены к виду † Micropercops pomahaka Schwarzhans, Lee & Gard, 2017. Местонахождение ископаемых останков: формация Помахака, река Помахака (Pomahaka Formation, Pomahaka River), Южный остров (South Island), Новая Зеландия (46,0° ю.ш., 169,2° в.д.; палеокоординаты 52,1° ю.ш., 176,6° з.д.). Окружающая среда — морская; среднезернистый, серый алевролит (среднезернистый серый ил с пнями). [W. Schwarzhans, D. E. Lee, and H. J. L. Gard. 2017. Otoliths reveal diverse fish communities in Late Oligocene estuarine to deep-water paleoenvironments in southern Zealandia. New Zealand Journal of Geology and Geophysics 60(4):433-464]
Головешки имеют относительно короткое, плотное тело, удлинённо-овальной формы, почти круглое в сечение, в передней части немного вальковатое, к хвостовому плавнику сжатое с боков. Плавники мягкие, без острых шипов. Два спинных плавника, расположенных очень близко друг к другу, при этом первый спинной плавник ниже и короче второго. D VI—IX, I—II 9—11. Второй спинной плавник по величине и форме похож на анальный. А I—III 7—10. Грудные плавники крупные, округлой формы. Брюшные плавники не соединены в диск, слабо развиты (V I 5). Хвостовой плавник округлой формы. Голова приплющена. Лобные кости с гребнем, начинающимся от места прикрепления posttemporale и идущим вперед почти до глаза, где правый и левый гребень соединяются тонким поперечным гребнем. Место прикрепления posttemporale ближе к затылочному гребню, чем к краю черепа (см. рисунок). Расстояние между точками прикрепления posttemporalia меньше, чем довольно узкое межглазничное пространство. Нижнеглоточные кости закруглены. Чувствительных каналов и пор нет (генипоры есть). Рот широкий, нижняя челюсть выдаётся вперёд. Верхнечелюстная кость у взрослых достигает вертикали заднего края глаза. Зубы на челюстях в несколько рядов, щетинковидные, не исключая внешнего ряда подвижные, несколько изогнутые. Клыковидных зубов нет. Зубы на сошнике в виде двух совершенно отделённых одна от другой групп, правой и левой, каждая с 6—7 довольно крупными зубами. На нёбных костях и на языке нет зубов. Предкрышка без шипа. Межжаберный промежуток узкий, жаберные отверстия простираются вперёд почти под передний край предкрышки. Жаберные тычинки бугорковидные, далеко одна от другой, 8—10. Существует зачаточная наджаберная полость. Чешуя умеренной величины, на боках неясно ктеноидная, на спине циклоидная, squ. 33—43, в поперечном ряду 17—22 чешуи. Голова покрыта чешуёй вплоть до середины лба, бока головы тоже покрыты чешуёй.
Распространение (карта): пресные и солоноватые воды Евразии.
Естественный ареал Perccottus расположен на Дальнем Востоке России (Амурская область, юг Хабаровского края, Приморский край, северо-запад Сахалинской области), в северо-восточном Китае и на севере Кореи. Большая часть естественного ареала ротана приходится на бассейн Амура, где ротан преимущественно населяет пойменные водоёмы этой реки и её притоков, включая такие крупные, как Зея, Сунгари и Уссури. На севере ареал головешки доходит до бассейна реки Тугур. На юг от бассейна Амура ротан известен из рек Японского моря, включая Суйфун, Тумень-улу, Ляохэ и из района города Люйшунь (бывший Порт-Артур). В бассейне реки Сунгари этот «бычок» (под таким названием был известен в Харбине) водится повсеместно, предпочитая, однако, стоячие воды или болота. В верховьях реки Сунгари ротан, по-видимому, отсутствует. На западе, вероятно, эта рыба не поднимается по Амуру выше Джалинды (Никольский, 1956). Таранец (1937) указывает местообитание ротана на северо-западе Сахалина, напротив лимана Амура, высказывая предположение, что он исторически недавно пересёк Татарский пролив. В литературе есть упоминания о его появлении в морской воде (Солдатов, Линдберг, 1930; Каредин, 1966; Дмитриев, 1971). Еловенко (1981) отмечает, что встречали ротана в море лишь после паводков. Вероятно, он выносится из озёр большой водой и в солёной воде долго не живёт. Недавно подтверждено обитание этого вида в пресноводных водоёмах Северо-Запада острова Сахалин (Иванов, Иванова, 2002).
В 1916 — 2008 гг. ротан широко распространился в Северной Евразии, вызывая угнетение многих популяций аборигенных видов беспозвоночных, рыб и амфибий. Современное географическое распространение ротана существенно шире, чем это было принято считать ранее. К настоящему времени за пределами своего нативного ареала этот вид обнаружен в 48 субъектах Российской Федерации (в 36 областях, 9 республиках, 3 краях), а также в Литве, Латвии, Эстонии, Белоруссии, Молдавии, Украине, Казахстане (на севере страны), Монголии, Польше, Словакии, Венгрии, Сербии, Болгарии и Румынии. Первые сведения о возможном появлении ротана поступили из Италии. Инвазийный ареал ротана простирается более чем на 100° с запада на восток и на почти 20° с севера на юг.
История первых интродукций ротана в водоемы западных районов Евразии началась в 1916 г. Этот вид распространился на обширных территориях вследствие перевозок людьми на значительные расстояния и последующего саморасселения в пределах водных бассейнов, а также вследствие вторичных (местных) перевозок людьми. В нативной и инвазийной частях ареала ротан населяет преимущественно старицы рек, мелководья озёр и разнообразные пруды. Этот вид поедает молодь других видов рыб, что ведёт к снижению экономической эффективности рыбоводных хозяйств [Залозных, 1984]. В природных водоёмах хищничество ротана может быть причиной угнетения и исчезновения популяций некоторых аборигенных видов рыб. Особенности биологии ротана позволяют ему населять водоёмы, недоступные для большинства местных видов рыб. Присутствие ротана в таких водоёмах ведёт к сокращению видового разнообразия макробеспозвоночных и амфибий [Reshetnikov, Manteifel, 1997; Manteifel, Reshetnikov, 2002; Reshetnikov, 2003].
Анализ данных показал, что инвазийный ареал ротана много шире, чем это было принято считать ранее (Reshetnikov, 2009). К настоящему времени за пределами своего нативного ареала (Рис. 1) ротан отмечен в бассейнах рек Висла, Преголя, Неман, Даугава, Великая, Нева, Онега, Северная Двина, Обь, Енисей, Дунай, Днестр, Днепр, Дон, Волга, Урал, а также в верховьях бассейна р. Амур (Рис. 2). Форма современного инвазийного ареала ротана определяется ландшафтными и климатическими факторами, а также существованием нескольких очагов инвазии, давших начало нескольким новым участкам ареала на значительном удалении один от другого. Некоторые из этих участков до сих пор обособлены, но некоторые другие расширились и слились. По-видимому, до настоящего времени московский, плесецкий, томский, байкальский и верхнеамурский участки ареала независимы, в то время как львовский (Украина), московский, нижегородский (илевский) и некоторые другие слились и представляют собой единый участок ареала (Рис. 2), простирающийся к востоку до Тюменской и Курганской областей России [Решетников, Чибилев, 2009] и далее до Омской области вблизи недавно расширившегося Томского очага инвазии. Новые очаги инвазии продолжают появляться и расширяться. Например, в 1990 г. ротан был зарегистрирован в городских прудах г. Томск, а в июле 2005 г. в результате специальной экспедиции было выявлено широкое распространение этого вида на несколько сотен километров вниз по течению р. Обь [Решетников, Петлина, 2007].
В Евразии инвазийные популяции ротана были найдены с 44° по 63° с. ш. и с 17° по 121° в. д. (без учёта неподтвержденной информации из Италии). Наиболее северная находка этого вида приурочена к оз. Плесцы в Архангельской области России [Шляпкин, Тихонов, 2001] и расположена на 8° севернее самой северной находки в пределах нативного ареала (оз. Бокус в бассейне р. Уда Хабаровского края России). В настоящее время наиболее южная находка ротана в западной Евразии расположена в Болгарии [Jurajda et al., 2006]. Западная граница основной части инвазийного ареала проходит по рекам Висла, Сан, Бодрог и Тисса. Очевидно, все названные реки были заселены ротаном вследствие расширения западно-украинского (львовского) очага инвазии, достоверно известного с 1980 г. Самая западная из подтверждённых находок расположена в окрестностях оз. Балатон в Венгрии [Harka et al., 2008]. Ещё более западная изолированная находка ротана в Италии требует дополнительного подтверждения. Наиболее восточные находки популяций ротана, относящиеся к приобретённому ареалу, расположены в Забайкальском крае России. В будущем возможно слияние верхнеамурского инвазийного участка с нативной частью ареала ротана. В настоящее время Perccottus glenii быстро расселяется по рекам Дунай, Иртыш и Обь. Ожидается его появление в р. Енисей.
Мясо ротана-головешки белоснежное, с приятным, чуть сладковатым вкусом. Этот диетический продукт отвечает всем требованиям качества. Но особого внимания заслуживает состав рыбного продукта. В филейной части рыбы в большом количестве содержится никотиновая кислота (витамин РР). Кроме этого активного компонента, в продукте также присутствуют: витамины А и D; легкоусвояемые белки; омега-кислоты; целый комплекс аминоксилот; токоферол или витамин Е. А, также минеральные элементы: молибден, цинк, сера, никель, хром и ещё целая группа минералов, в которых нуждается организм человека. Отдельно стоит отметить, что состав мяса ротана представляет собой идеально сбалансированную формулу веществ, которые в едином комплексе оказывают положительное действие на течение практически всех биохимических процессов. Но главная ценность рыбного продукта заключается в его низкой калорийности. Пищевая ценность стограммовой порции отварного мяса составляет всего 88 ккал. Конечно, у жареной рыбы этот показатель будет выше, но не настолько, чтобы отказаться от вкусного блюда даже при похудении. Дело в том, что мясо ротана очень быстро усваивается пищеварительной системой, поэтому жиры впрок организм просто не успевает отложить.
В кулинарном отношении травянка считается универсальным рыбным продуктом, отчего ценность этой рыбы возрастает в 2 раза. Из этой скромной рыбешки можно готовить: легкие салаты; супы и другие первые блюда; домашние консервы; омлет; запеканку; котлеты; начинки для пирогов; тушеные и жареные блюда (см. фото). Белое с розовым оттенком мясо головешки имеет нежную структуру, в нём практически отсутствуют крупные кости. Впрочем их удаление не вызывает проблем. Знатоки рыбной кулинарии уверяют, что жареный ротан по вкусовым качествам превосходит даже карася, который считается своеобразным «эталоном» рыбного вкуса.
Аквариум для рыб подойдёт от 100—150 литров. На дно насыпают мелкий песок серого или жёлтого цвета. Из декораций подойдут ветвистые коряги. Растениями засаживают 60% от площади дна. Не помешают и плавающие на поверхности растения, типа водокраса. Молодых ротанов можно содержать группой, но следует понимать, что доминирующее количество самцов может привести к постоянным потасовкам (особенно в небольших емкостях), вплоть до выбивания глаз. Парное содержание тоже не совсем удачно. Вечно готовый к спариванию самец не даёт прохода самке. Соседями могут быть серебряные караси, обыкновенные караси, вьюны, лини и карпы. Едят ротаны любой живой корм и пищу животного происхождения — кусочки рыбы, мяса, креветки. Очень прожорливы. Поэтому кормят рыб не чаще одного раза в день, устраивая на недели 1—2 разгрузочных дня. При содержание ротанов можно обходится без технических средств, но маленький внутренний фильтр не помешает, так как он будет препятствовать образованию застойных зон в аквариуме. Температура содержания 15—25 °С, pH 6,5—7,5, dH 2—15.
Словарь названий пресноводных рыб СССР. Г.У.Линдберг и А.С.Герд 1972 г.
29.1.0. Perccottus Dybowski, 1877 — Головешки.
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
Очень близки
друг к другу заходящие дальше других
на север пресноводные роды
головешковых — мексиканский Гобиоморус и
восточноазиатская Головешка.
Род Головешка (Perccottus) представлен тремя видами в реках
Китая и одним из них в Советском
Союзе.
Рыбы пресных вод СССР и сопредельных стран. Л.С.Берг 1948 г.
1. Род PERCCOTTUS DYBOWSKI
Perccottus Дыбовский, Изв. Сиб. отд. Русск. геогр. общ., VIII, 1877, стр. 28 (тип: Perccottus glenii). — Берг, Зап. Акад. Наук (8), XXIV, № 9, 1909, стр. 207.
Брюшные плавники не соединены в диск, с I 5 лучами, слабо развиты. На нёбных костях и на языке нет зубов. Зубы на сошнике в виде двух совершенно отделенных одна от другой групп, правой и левой, каждая с 6—7 довольно крупными зубами. Зубы на челюстях в несколько рядов, щетинковидные, не исключая внешнего ряда подвижные, несколько изогнутые; клыковидных зубов нет. Чешуя умеренной величины, на боках неясно ктеноидная, на спине циклоидная. Голова покрыта чешуей вплоть до середины лба; бока головы покрыты чешуей. Предкрышка без шипа. Межжаберный промежуток узкий, жаберные отверстия простираются вперед почти под передний край предкрышки. Жаберные тычинки бугорковидные, далеко одна от другой, 8—10. Тело не очень удлиненное, спереди вальковатое, сзади сжатое. Голова приплющена, рот широкий, нижняя челюсть выдается вперед; верхнечелюстная кость у взрослых достигает вертикали заднего края глаза. D VI—IX, I—II 9—11, А I—III 7—10, squ. 33—43, в поперечном ряду 17—22 чешуи. Лобные кости с гребнем, начинающимся от места прикрепления posttemporale и идущим вперед почти до глаза, где правый и левый гребень соединяются тонким поперечным гребнем. Место прикрепления posttemporale ближе к затылочному гребню, чем к краю черепа. Расстояние между точками прикрепления posttemporalia меньше, чем довольно узкое межглазничное пространство. Нижнеглоточные кости закруглены. Существует зачаточная наджаберная полость. Чувствительных каналов и пор нет (генипоры есть, см. рис. 775 и 776).
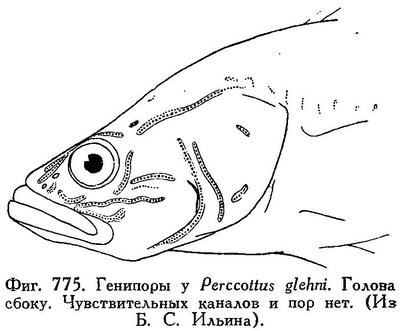
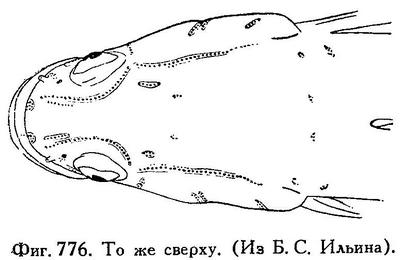
Близок к роду Philypnus Valenciennes 1837 (nomen conservandum, = Gobiomorus Lacepède 1800), у которого зубы на сошнике в виде сплошной поперечной полоски, жаберные тычинки палочковидные, место прикрепления posttemporale иное; род Philypnus с двумя видами свойствен Центр. Америке и прилегающим местам (Калифорния, Техас, Эквадор, Суринам, Антильские острова и проч.).
1—2 вида в бассейне Амура и в Китае.