Carcharhinus melanopterus (Quoy & Gaimard, 1824)
Чернопёрая рифовая акула, Чернопёрая рифовая серая акула, Чёрноплавниковая рифовая акула, Чёрноплавниковая рифовая серая акула, Мальгашская ночная акула, Мальгашская рифовая акула, Мальгашская ночная серая акула, Чёрноконечная рифовая акула, Чёрноконечная рифовая серая акула, Кархаринус меланоптерус (Rus),
Blacktip reef shark, Blacktip shark, Blackfin reef shark, Black finned shark, Black tips nilow, Reef blacktip shark, Guliman, Requien shark
(Eng).
Синонимы:
Carcharhinus melanoptera (Quoy & Gaimard, 1824)
Carcharhinus melanoptures (Quoy & Gaimard, 1824)
Carcharhinus melanoterus (Quoy & Gaimard, 1824)
Carcharias commersonii (Blainville, 1816)
Carcharias elegans Ehrenberg, 1871
Carcharias marianensis Engelhardt, 1912
Carcharias melanopterus Quoy & Gaimard, 1824
Carcharias playfairii Günther, 1870
Carcharinus melanopterus (Quoy & Gaimard, 1824)
Hypoprion playfairii (Günther, 1870)
Mapolamia spallanzanii (non Péron & Lesueur, 1822)
Squalus carcharias minor Forsskål, 1775
Squalus commersonii Blainville, 1816
Squalus ustus Duméril, 1824
Материал подготовил Фактистов И. Е., 2021 г. Сайт www.fishbiosystem.ru
Новый вид впервые в соавторстве научно описали как Carcharias melanopterus в 1824 году в ходе исследовательского путешествия на корвете «Урания»:
• французский учёный Жан-Рене-Констан Куа (Жан-Рене-Констан Куа — фр. Jean René Constant Quoy, 1790 — 1869, французский врач, зоолог и анатом, в 1806 году начал обучение в школе морской медицины в Рошфоре (Rochefort), после чего служил вспомогательным хирургом в поездке на Антильские острова (Antilles, 1808—1809), в 1814 году получил докторскую степень в Монпелье (Montpellier), был главным хирургом (1814—1815) в поездке на Реюньон (Réunion), с 1817 по 1820 годы служил корабельным врачом во время кругосветного плавания французского корвета «Урания» («L'Uranie») под командованием Луи де Фрейсине (Louis de Freycinet), с 1822 по 1825 годы вместе с Жозефом-Полем Гемаром (Joseph Paul Gaimard) служил натуралистом во время кругосветного плавания на борту «La Coquille» под командованием Луи Исидора Дюперре (Louis Isidore Duperrey) и на борту «Астролябии» («L'Astrolabe») с 1826 по 1829 годы под командованием Жюла Дюмона-Дюрвиля (Jules Dumont d'Urville), в 1830 году избран членом Французской Академии наук (Académie des sciences), с 1848 по 1858 годы служил генеральным инспектором Морского бюро медицины и хирургии (Naval Bureau of Medicine and Surgery), вместе с Гемаром описали около 80 новых видов рыб, в его честь названо 4 вида рыб);
• французский учёный Жозеф-Поль Гемар (Жозеф-Поль Гемар — фр. Joseph Paul Gaimard, 1796 — 1858, французский хирург, естествоиспытатель, ихтиолог и зоолог, изучал медицину в военно-морской медицинской школе в Тулоне (Toulon), вместе с Жан-Рене-Констан Куа (Jean René Constant Quoy) работал натуралистом на кораблях «Урания» («L'Uranie») под командованием Луи де Фрейсине (Louis de Freycinet) 1817—1820 гг. и «Астролябии» («L'Astrolabe») под командованием Жюла Дюмона-Дюрвиля (Jules Dumont d'Urville) 1826—1829 гг., был командиром экспедиции (1835—1836) в Арктическом море (Arctic Sea) на корвете «La Recherche» с посещением прибрежной Исландии (Iceland) и Гренландии (Greenland), с 1838 по 1840 год, снова на борту «La Recherche», был руководителем научной экспедиции в Лапландию (Lapland), Шпицберген (Spitzbergen) и на Фарерские острова (Faroe Islands), в 1839 году избран членом-корреспондентом Петербургской академии наук, вместе с Куа описали около 80 новых видов рыб, в его честь названо 2 вида рыб).
В 1824 году их записки были опубликованы в отчёте о кругосветном путешествии на корвете «Урания», написанном Луи де Фрейсине. [Quoy, J.R.C. & Gaimard, J.P. (1824) Description des Poissons. Chapître IX. In: Freycinet, L. de, Voyage autour du Monde…exécuté sur les corvettes de L. M. «L’Uranie» et "La Physicienne, " pendant les années 1817, 1818, 1819 et 1820. Paris. Description des Poissons. Description des Poissons. Chapter IX.: 192—401 [1-328 in 1824; 329—616 in 1825], Atlas pls. 43-65] Лейкотипом был назначен самец длиной 59 см, пойманный в ходе экспедиции в водах Индонезии у берегов острова Вайгео (Waigeo Island). Более поздние исследователи отнесли вид к роду серых акул.
Чернопёрая рифовая акула, Чернопёрая рифовая серая акула, Чёрноплавниковая рифовая акула, Чёрноплавниковая рифовая серая акула, Мальгашская ночная акула, Мальгашская рифовая акула, Мальгашская ночная серая акула, Чёрноконечная рифовая акула, Чёрноконечная рифовая серая акула (Carcharhinus melanopterus) — вид хрящевых рыб из отряда кархаринообразных (Carcharhiniformes Compagno, 1977) семейства серые акулы (Carcharhinidae Jordan & Evermann, 1896), обитающих в тропических и субтропических водах Индо-Тихоокеанского региона.
Название рода Carcharhinus происходит от греческих слов καρχαρος (karcharos) — «острозубый», «зубастый» и ρινος (rhinos) — «нос», указывает на то, что это зубастая акула с длинным рылом, по другим данным καρχαρος (karcharos) — «острый», «зазубренный» и ρινη (rhine) — «акула» (древнее имя для акулы, полученное от греческого слова рашпиль), оба слова указывают на колючую, шершавую кожу акулы. Видовой эпитет происходит от древнегреческого слова μελανος (melanos) — «чёрный» и греческого слова πτερον (pteron) — «крыло» и ссылается на чёрные кончики спинных и хвостового плавников, характерные для этого вида.
В 1965 году Международная комиссия по зоологической номенклатуре назначила его типовым видом рода. В ранней литературе научное название этой акулы было ошибочно присвоено виду Carcharhinus spallanzani, позднее признанному синонимом Carcharhinus sorrah.
Подобно прочим членам рода серых акул, филогенетическая позиция мальгашской ночной акулы остается до конца не определённой. На основании морфологии Джек Гэррик (Джон Эндрю Фрэнк Гаррик, «Джек» Гаррик — John Andrew Frank Garrick, «Jack» Garrick, 1928 — 1999, выдающийся ихтиолог из Новой Зеландии) в 1982 году предположил, что самым близкородственным видом является пугливая акула (Carcharhinus cautus). [Garrick, J.A.F. «Sharks of the genus Carcharhinus». NOAA Technical Report, NMFS Circ:1982. — p.445]
Проведя в 1988 году морфологический анализ, Леонард Компаньо (Леонард Джозеф Виктор Компаньо — Leonard Joseph Victor Compagno, 1943 г.р., мировой авторитет в области таксономии акул) сделал предположение о том, что существует близкая связь не только с пугливой акулой (Carcharhinus cautus), но и ещё с четырьмя видами, однако их взаимоотношения так и не было точно установлены. Проведённый в 1998 году аллозимный анализ принёс неоднозначные результаты, так как было обнаружено, что мальгашская ночная акула образует политомию с десятью видами рода серых акул. [Naylor, G.J.P. The phylogenetic relationships among requiem and hammerhead sharks: inferring phylogeny when thousands of equally most parsimonious trees result // Cladistics. — 1999. — Vol. 8, № 4. — P. 295—318.]
У мальгашских ночных акул характерное для серых акул торпедообразное, обтекаемое, умеренно толстое тело. Крупные грудные плавники имеют форму в виде изогнутого узкого серпа и сужаются к закруглённым кончикам. Длина их переднего края составляет 17—19 % от общей длины тела. Первый спинной плавник тоже значительного размера (умеренно крупный), с узким, закруглённым или заострённым концом, его свободный задний кончик образует изгиб в виде буквы «S», основание расположено позади свободных кончиков грудных плавников. Второй спинной плавник меньше первого, он расположен напротив анального плавника или чуть ближе к голове. Его высота составляет 3,4—4,1 % от общей длины тела. Длина внутреннего края равна 0,8—1,1 % от высоты. Гребень между спинными плавниками отсутствует. Хвост сильно асимметричный, с хорошо развитой верхней лопастью, волнообразным гребнем вдоль дорсальной поверхности верхней лопасти и выемкой под вершиной этой лопасти. Пять жаберных щелей, последние две расположены над основанием грудного плавника. Рыло короткое, широкое и закруглённое. Овальные, довольно крупные глаза вытянуты по горизонтали (см. фото). Ноздри обрамлены спереди кожными складками, образующими сосковидные лопасти (см. фото). Дыхальце отсутствует. Складки верхней губы очень короткие. Симфизальные зубы отсутствуют. Во рту с каждой стороны челюстей имеются по 11—13 верхних и 10—12 нижних зубных рядов (см. фото). Верхние зубы имеют форму треугольника, их края зазубрены, постав от вертикального до наклонного. С каждой стороны от основания основной вершины имеются маленькие острия (см. фото). Нижние зубы похожи на верхние, однако зубчики по краям у них тоньше (см. фото). По сравнению с зубами самок зубы взрослых самцов изогнуты сильнее.
Окраска дорсальной поверхности серо-коричневого или жёлто-коричневого цвета, брюхо белое. Первый спинной плавник и нижняя доля хвоста с яркими, широкими чёрными кончиками, ограниченными более светлыми участками. Остальная часть хвостового плавника и другие плавники с отчетливо чёрными краями. Длина редко превышает 1,6 м, максимальная зарегистрированная длина 1,8 м, по некоторым данным 2 м. Максимальный зарегистрированный вес 13,6 кг.
Распространение (карта): прибрежные районы тропической и субтропической зоны Индийского и Тихого океанов (35 ° с. ш. — 25 ° ю. ш., 7 ° в. д. — 134 ° з. д.). После открытия Суэцкого канала акулы проникли и адаптировались в Средиземном море. В Индийском океане они встречаются от ЮАР до Красного моря, включая воды Мадагаскара, Маврикия и Сейшельских островов. На востоке их ареал простирается вдоль Индийского субконтинента до Юго-Восточной Азии, включая Шри-Ланку, Андаманские острова и Мальдивы. В Тихом океане они распространены от южного побережья Китая, Филиппин и Индонезии до севера Австралии, Новой Каледонии и многочисленных островов (Маршалловы острова, Острова Гилберта, Острова Общества, Гавайи и Туамоту). Несмотря на информацию в публичных источниках, нет подтверждённых данных, свидетельствующих о присутствии этого вида в водах Японии, а японские образцы, скорее всего, прибыли из Тайваня.
Мальгашские ночные акулы предпочитают мелководье, не бояться заходить на рифовые платформы (см. Видео). В то же время, этот вид был отмечен на глубине до 75 м. Иногда можно наблюдать, как эти акулы патрулируют риф, выставив над водой спинной плавник. Молодые акулы держатся на небольшой глубине, тогда как взрослые встречаются на обрывах у края рифа. Мальгашские ночные акулы попадаются в солоноватых водах эстуариев рек и в озёрах Мадагаскара, а также в пресноводных водоёмах Малайзии, хотя они не столь терпимы к пониженной солёности по сравнению с акулой-быком (Carcharhinus leucas). На Альдабре во время отлива они скапливаются в каналах между рифовыми платформами, а когда наступает прилив, заплывают в мангровые заросли. По некоторым данным, мальгашские ночные акулы, обитающие в северном и южном пиках своего ареала, совершают миграции.
Наряду с рифовыми (Triaenodon obesus) и темнопёрыми серыми (Carcharhinus amblyrhynchos) акулами мальгашские ночные акулы являются самыми многочисленными акулами, обитающими на коралловых рифах индо-тихоокеанской области. Мальгашские ночные акулы предпочитают держаться на мелководье, тогда как два других вида населяют более глубокие воды. Стремительные и активные мальгашские ночные акулы встречаются как по одиночке, так и небольшими группами, изредка они собираются в крупные стаи (см. фото). В целом неполовозрелые и взрослые акулы не разделяются по половому признаку. У некоторых особей наблюдается привязанность к индивидуальному участку обитания, который они не покидают в течение нескольких лет.
Исследования путём мечения акул, проведённые на атолле Пальмира в центральной части Тихого океана, показали, что площадь индивидуального участка обитания мальгашских ночных акул приблизительно равная 0,55 км2, и является наименьшей среди всех видов акул. Размер и расположение участка в течение суток не меняются. От 3 до 17 % всей площади занимают излюбленные места охоты, непропорционально занимаемые постоянно живущими на участке акулами. Большую часть времени мальгашские ночные акулы плавают взад-вперёд вдоль края рифа, предпринимая периодические вылазки на песчаные пустоши (см. фото). Ночью с приливом средняя скорость движения снижается, возможно, из-за притока прохладной воды, снижающего их метаболизм, или же вслед за замедлением движения рыб, на которых они охотятся. Мальгашские ночные акулы, живущие в водах Альдабры, в целом более подвижны по сравнению акулами, обитающими в атолле Пальмира, и преодолевают за 7 часов более 2,5 км.
Мальгашские ночные акулы, особенно мелкие особи, могут стать добычей крупных рыб, таких как груперы (род Epinephelus), темнопёрые серые акулы (Carcharhinus amblyrhynchos), тигровые акулы (Galeocerdo cuvier) и крупные сородичи. На атолле Пальмира они избегают столкновения с тигровыми акулами, держась подальше от центральных, более глубоких лагун.
Зачастую мальгашские ночные акулы являются преобладающими хищниками в своей экосистеме и играют ведущую роль в формировании структуры синэкологии. Их рацион в основном состоит из костистых рыб, включая кефалей (семейство Mugilidae), груперов (род Epinephelus), ворчунов (семейство Haemulidae), каранксов (род Caranx), мохарровых (семейство Gerreidae), губанов (семейство Labridae), хирурговых (семейство Acanthuridae) и силлаговых (семейство Sillaginidae). В Индийском океане группы мальгашских ночных акул преследуют вдоль берега стаи кефалей, что облегчает им охоту. Кроме того, они поедают кальмаров, осьминогов, креветок, каракатиц, а также падаль и мелких акул и скатов. У северного побережья Австралии этот вид акул охотится на морских змей, включая бородавчатых змей Acrochordus granulatus, Hydrelaps darwiniensis, ластохвостов и ластохвостых змей. Акулы, обитающие на атолле Пальмира, нападают на птенцов морских птиц, упавших из гнезда в воду. В желудке мальгашских ночных акул обнаруживают также водоросли, кораллы, гидроидных, крыс, мшанок и камни.
Исследователи, работавшие на атолле Эниветок, обнаружили, что мальгашских ночных акул привлекает плеск или звук, издаваемый при ударе металла о твёрдый предмет под водой, а также запах как раненой, так и целой рыбы. Как и у большинства акул, у мальгашских ночных акул в сетчатке нет колбочек, поэтому они плохо различают цвета и мелкие детали. Их зрение адаптировано распознавать малейшие движения и контраст в условиях плохой освещённости, чему способствует наличие отражающего свет тапетума. Эксперименты показали, что эти акулы способны засечь небольшие объекты на расстоянии 1,5—3 м, но не могут чётко различить их форму. Кроме того, мальгашские ночные акулы обнаруживают добычу с помощью электрорецепции: их ампулы Лоренцини обладают чувствительностью около 4 нановольт/см в диапазоне 25 см. У них хорошо развито обоняние — они реагируют на экстракт рыбы, разведённой в воде в пропорции 1:10 млрд. Подобно темнопёрым рифовым акулам (Carcharhinus amblyrhynchos), в присутствии сородичей они смелеют и приходят в возбуждение, а в экстремальной ситуации могут впасть в пищевое безумие. Этот вид акул более активен в ночное время.
Подобно прочим представителям семейства серых акул, мальгашские ночные акулы размножаются путём живорождения (см. Видео), хотя между популяциями наблюдаются существенные различия жизненного цикла. У акул, обитающих у северного побережья Австралии, воспроизводство имеет годичный цикл, спаривание происходит с января по февраль, тогда как у в водах Муреа брачный сезон длится с ноября по март. У побережья Альдабры цикл воспроизводства длится два года, вероятно, высокая межвидовая конкуренция за добычу ограничивает способность самок ежегодно вынашивать потомство. Ранние отчёты о мальгашских ночных акулах, обитающих в Индийском океане (1978), у берегов Мадагаскара (1961) и в Красном море (1964), свидетельствовали о том, что цикл воспроизводства у них был двухгодичный, сезон размножения длился с июня по июль и с декабря по январь. Если говорить точнее, укороченный цикл воспроизводства у этого вида может быть связан с повышенной температурой воды.
Будучи готовой к спариванию, самка мальгашской ночной акулы начинает, опустив голову, плавать у дна по синусоидной траектории. Наблюдения в дикой природе дают основание предположить, что самки испускают химические сигналы, которые позволяют самцам их засечь. После обнаружения готовой самки самец приближается к ней приблизительно на расстояние 15 см и начинает следовать за ней, устремив рыло по направлению к её клоаке. В процессе ухаживания самец может укусить самку в жаберную область или за грудные плавники, нанесённые им раны заживают через 4—6 недель. После завершения стадии преследования самец толкает самку и поворачивает её на бок, так чтобы её голова оказалась напротив дна, а хвост приподнялся, и вставляет один из птеригоподий в клоаку. Совокупление длится несколько минут, затем акулы разъединяются и продолжают вести себя как обычно. В водах Муреа некоторые взрослые самки спариваются и приносят потомство в одно и то же время каждый год с небольшим отклонением в несколько недель, тогда как у молодых акул такого постоянства не наблюдается. Кроме того, они чаще беременеют после совокупления.
В Индийском океане и у тихоокеанских островов беременность длится 10—11 месяцев и 7—9 месяцев у северного побережья Австралии. Ранее продолжительность беременности оценивалась в 16 месяцев, хотя обоснованность этих данных неоднократно оспаривалась. У самок имеется один функциональный яичник, расположенный справа, и два функциональных яйцевода, разделённые на камеры, содержащие по одному эмбриону. Только что отложенные яйца, заключённые в капсулу, имеют размер 3,9 х 2,6 см. После вылупления на первой стадии развития эмбрионы питаются желтком, содержащимся в желточном мешке. Спустя два месяца длина эмбрионов достигает 4 см, у них уже хорошо развиты внешние жабры. На четвёртом месяце желточный мешок начинает трансформироваться в плацентарное соединение, которое прикрепляется к стенке матки. К этому времени у эмбрионов кончики плавников приобретают характерную чёрную окантовку. Пятимесячные эмбрионы имеют длину 24 см, внешние жабры исчезают, плацента полностью сформирована. Питание желтком сохраняется до седьмого месяца беременности.
Роды происходят с сентября по ноябрь. В качестве природных питомников самки используют мелководье внутри рифов. В Индийском океане и у северного побережья Австралии длина новорожденных составляет 40—50 см, тогда как в водах тихоокеанских островов самая маленькая свободно плавающая мальгашская ночная акула имела длину около 33 см. В помёте 2—5 новорожденных (обычно 4), численность в помёте не коррелирует с размером самки. Молодые мальгашские ночные акулы образуют обычно большие группы у берега на мелководье с песчаным дном или в мангровых зарослях, где вода едва покрывает их спину. Во время прилива они могут заплывать на затапливаемые коралловые плато. Сначала они растут быстро, прибавляя в среднем по 23 см в год за первые два года жизни. Подростки и взрослые акулы вырастают в год в среднем на 5 см. Самцы и самки данной акулы достигают половой зрелости при длине 95 см и 97 см (северное побережье Австралии) и 105 см и 110 см (Альдабра).
На мальгашских ночных акулах паразитируют ленточные черви Anthobothrium lesteri, Nybelinia queenslandensis, Otobothrium alexanderi, и Platybothrium jondoeorum, слизистые споровики рода Unicapsula, и моногенеи Dermophthirius melanopteri. Был зарегистрирован случай инфекционного заболевания геморрагической септицемией с летальным исходом, вызванной бактерией Aeromonas salmonicida.
Паразиты (список Юргена Поллерспёка — Jürgen Pollerspöck):
• Myxosporea (класс Миксоспоридии, Слизистые споровики — Myxosporea): Ceratomyxa carcharhini GLEESON & ADLARD, 2011; Ceratomyxa melanopteri GLEESON & ADLARD, 2011.
• Cestoda (класс Цестоды, Ленточные черви — Cestoda): Anthobothrium lesteri WILLIAMS, BURT & CAIRA, 2004; Callitetrarhynchus gracilis (RUDOLPHI, 1819) PINTNER, 1931; Dasyrhynchus basipunctatus (CARVAJAL, CAMPBELL & CORNFORD, 1976); Dasyrhynchus giganteus (DIESING 1850); Dollfusiella sp.; Eutetrarhynchus sp.; Floriceps minacanthus CAMPBELL & BEVERIDGE, 1987; Mixonybelinia edwinlintoni (DOLLFUS, 1960); Nybelinia africana DOLLFUS, 1960; Nybelinia queenslandensis JONES & BEVERIDGE, 1998; Otobothrium alexanderi PALM, 2004; Otobothrium australe PALM, 2004; Otobothrium carcharidis (SHIPLEY & HORNELL, 1906); Otobothrium crenacolle LINTON, 1890; Otobothrium cysticum (MAYER, 1842); Otobothrium penetrans LINTON, 1907; Otobothrium sp.; Phyllobothrium minutum WILLIAMS, 1968; Phyllobothrium pammicrum SHIPLEY & HORNELL, 1906; Platybothrium jondoeorum HEALY, 2003; Prochristianella sp.; Pterobothrium heteracanthum DIESING, 1850; Scyphophyllidium angustum (LINTON, 1889) RUHNKE, 2011; Tentacularia coryphaenae BOSC, 1797; Tetrarhynchobothrium sp..
• Nematoda (тип Нематоды, Круглые черви — Nemata): Pulchrascaris chiloscyllii (JOHNSTON & MAWSON, 1951).
• Copepoda [надотряд Копеподы, Веслоногие ракообразные — класса Челюстеногие ракообразные (Maxillopoda)]: Kroyeria longicauda CRESSEY, 1970; Nemesis sp..
• Isopoda (класс Высшие ракообразные — Malacostraca): Gnathia trimaculata COETZEE, SMIT, GRUTTER & DAVIES, 2009.
• Hirudinea (класс Клителлаты — Clitellata): Pontobdella macrothela (SCHIMARDA, 1861).
Обычно мальгашские ночные акулы ведут себя довольно робко и уплывают при появлении человека. Однако известны случаи нападения акул данного вида на людей, но со смертельным исходом инцидентов отмечено не было. В присутствии раненой рыбы (например, на подводной охоте) эта акула легко приходит в возбуждение и может укусить дайвера. Отмечены также случаи, когда акула кусала слишком назойливых пловцов. До 2013 года было зафиксировано 11 неспровоцированных атак, приписываемых этому виду акул. Чаще всего акулы кусают людей за ноги, спутав их со своей естественной добычей, но не наносят тяжёлых повреждений. На Маршалловых островах местные жители избегают укусов, предпочитая преодолевать мелководье, вплавь, а не вброд.
Эти акулы регулярно попадают в рыболовные сети в прибрежных водах Таиланда и Индии. Этот вид не представляет интереса для коммерческого рыболовства. Мясо используют в пищу, в продажу оно поступает в свежем, замороженном, вяленом и копчёном виде. Международный союз охраны природы присвоил этому виду статус сохранности «Близкий к уязвимому положению». Несмотря на повсеместное распространение, численность некоторых локальных популяций снижается из-за перелова. У мальгашских ночных акул медленный цикл воспроизводства. Этот вид акул часто содержат в аквариумах зоопарков и океанариумах.
Акулы Мирового океана: Справочник-определитель. Е. П. Губанов, В. В. Кондюрин, Н. А. Мягков 1986 г.
Carcharhinus melanopterus (Quoy et Gaimard, 1824) — черноперая рифовая акула
[Carcharias melanopterus Quoy et Gaimard, 1824; Carcharias (Hypoprion) playfairii Günther, 1870]
Окраска спины темно-серая или буроватая, брюхо беловатое. Края лопастей хвостового плавника и верхушки всех плавников окрашены в черный цвет (см. рис. 52, б). Рыло короткое, равное минимальному расстоянию между ноздрями или чуть превышающее его. Тело прогонистое, стройное. Зубы на челюстях сходны по форме, но на верхней челюсти несколько шире и крупнее, чем на нижней, незначительно скошены к углам рта. Зубная формула 12-14 — 1-2 — 12-14 / 10-14 — 1-3 — 10-14 (в единичных случаях на верхней челюсти может быть по 11 зубов на каждой стороне).
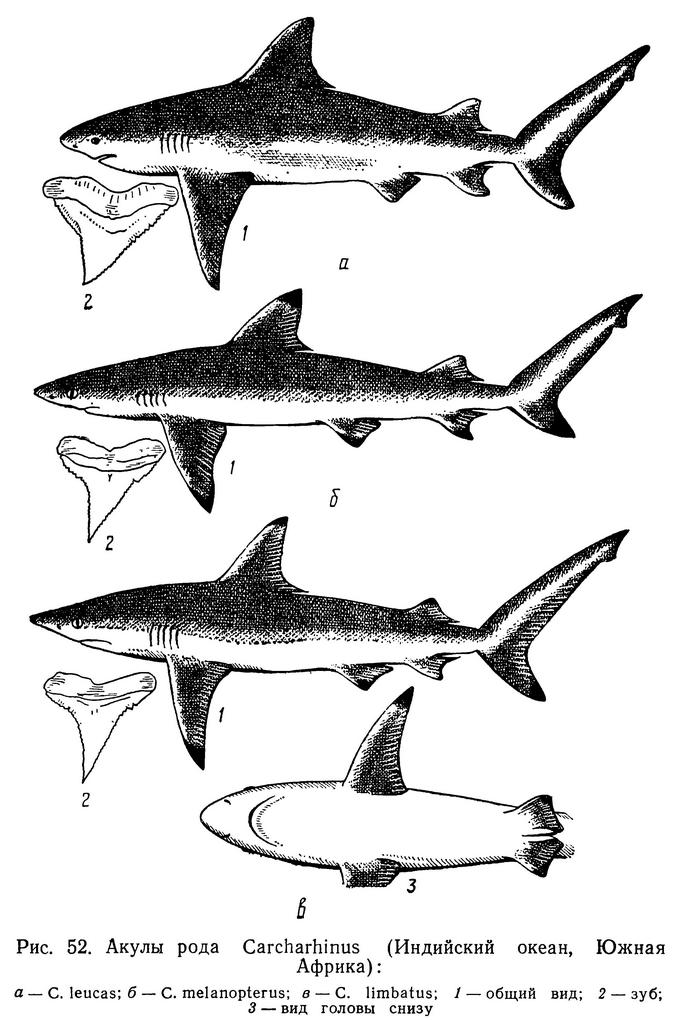
Биология. Максимальная длина тела 1,8 м. Половозрелость наступает при длине около 1 м. Длина молоди при рождении не превышает 50 см. Живородящие. Число молоди от одной самки 10—13 (обычно 4—7). Хищники. Питаются стайными костистыми рыбами и головоногими моллюсками.
Распространение. Индийский (включая Красное море) и Тихий океаны (как в открытых водах, так и у берегов) на глубинах не более 150 м.
Хозяйственное значение. Объекты ярусного промысла. Для пищевых целей.
Гюнтером (Günther, 1870) по голотипу, пойманному у берегов Мадагаскара, описан четвертый вид рода Hypoprion playfairii, который отличается от Hypoprion hemiodon наличием черных пятен на плавниках и формой зубов нижней челюсти. Отмечено небольшое отличие и в относительной длине основания второго спинного плавника. Уровень этих различий невелик. Вероятно, в данном случае имеет место географическая или индивидуальная изменчивость, что не позволяет пока говорить о самостоятельности вида Hypoprion playfairii (Günther, 1870), тем более что ни литературные данные, ни материалы многочисленных советских научно-исследовательских и поисковых рейсов в данный район не подтверждают его существования.