Ксенакантообразные (Xenacanthiformes) — отряд вымерших хрящевых рыб (Chondrichthyes Huxley, 1880), похожих на акул. Его относят к надотряду или инфраклассу ксенакантоморф (Xenacanthimorpha), точное положение которого дискутируется — это подчинённый таксон либо хрящевых рыб (Chondrichthyes Huxley, 1880), либо пластиножаберных (Elasmobranchii Bonaparte, 1838). Представители отряда жили с девонского по меловой периоды (407,6—66,0 млн лет назад). Характерным отличием данного отряда являлся зубчатый шип у основания шеи. Также стоит отметить строение зубов. Основание каждого зуба имело бугорки. Наиболее хорошо изученными представителями ксенакантообразных являются Xenacanthus и Orthacanthus. Xenacanthus, возможно, достигали в длину 4 метров. Рыбы обитали в пресной воде.
В отряде выделяют 3 семейства (не считая устаревшего Orthacanthidae Heyler & Poplin, 1989):
• Семейство † Diplodoselachidae Dick, 1981.
• Род † Dicentrodus Traquair, 1888
• Род † Diplodoselache Dick, 1981
• Род † Hagenoselache Hampe & Heidtke, 1997
• Род † Lebachacanthus Soler-Gijon, 1997
• Род † Orthacanthus Agassiz, 1843
• Семейство † Xenacanthidae Fritsch, 1889.
• Род † Acanthopleurus Agassiz, 1843
• Род † Aganodus Owen, 1867
• Род † Compsacanthus Newberry, 1856
• Род † Diacranodus Garman, 1885
• Род † Fayolia Renault & Zeiller, 1884
• Род † Lepacanthus Arthaber, 1896
• Род † Lophacanthus Stock, 1880
• Род † Mooreodontus Hampe & Schneider, 2010
• Род † Ochlodus Owen, 1867
• Род † Phricacanthus Davis, 1879
• Род † Platyacanthus Fritsch, 1889
• Род † Pleuracanthus Agassiz, 1837
• Род † Plicatodus Hampe, 1995
• Род † Pternodus Owen, 1867
• Род † Triodus Jordan, 1849
• Род † Wurdigneria Richter, 2005
• Род † Xenacanthus Beyrich, 1848
• Семейство † Xenosynechodontidae Gluckman, 1980.
• Род † Xenosynechodus Gluckman, 1980
• Род † incertae sedis («неопределённого положения» — латинское выражение, указывающее на то, что положение таксона в системе неясно).
• Род † Anodontacanthus Davis, 1881
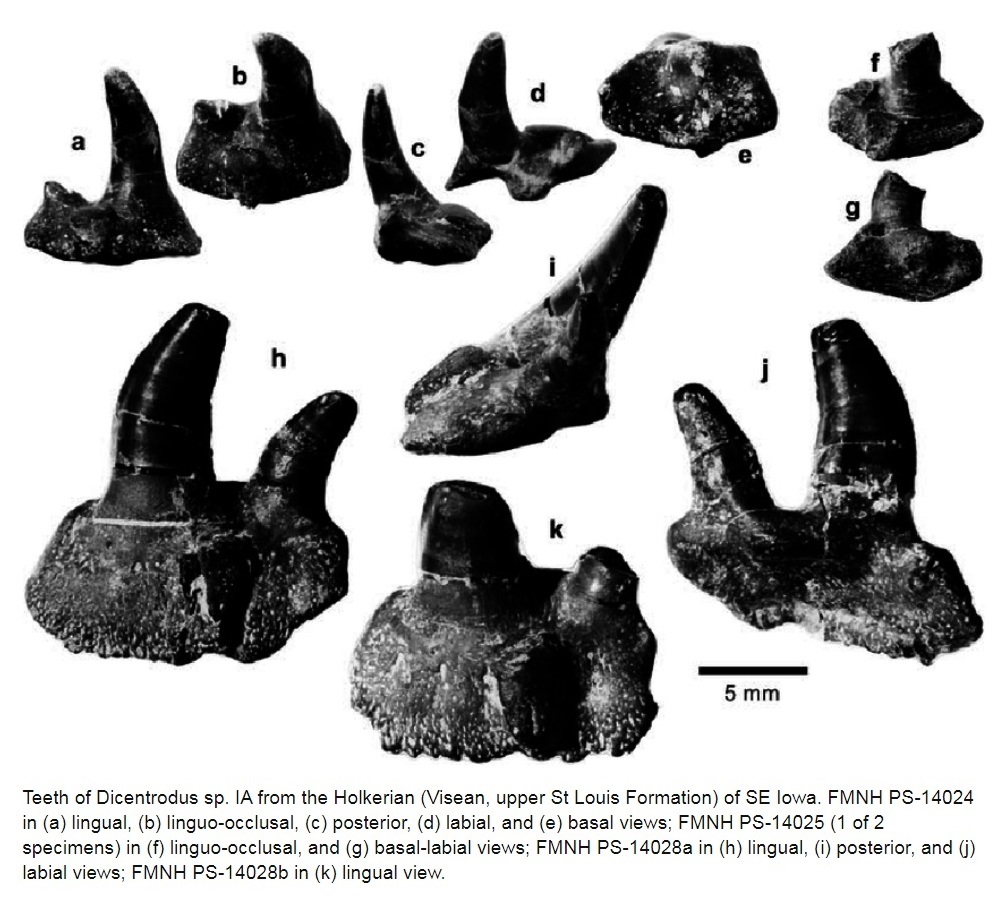
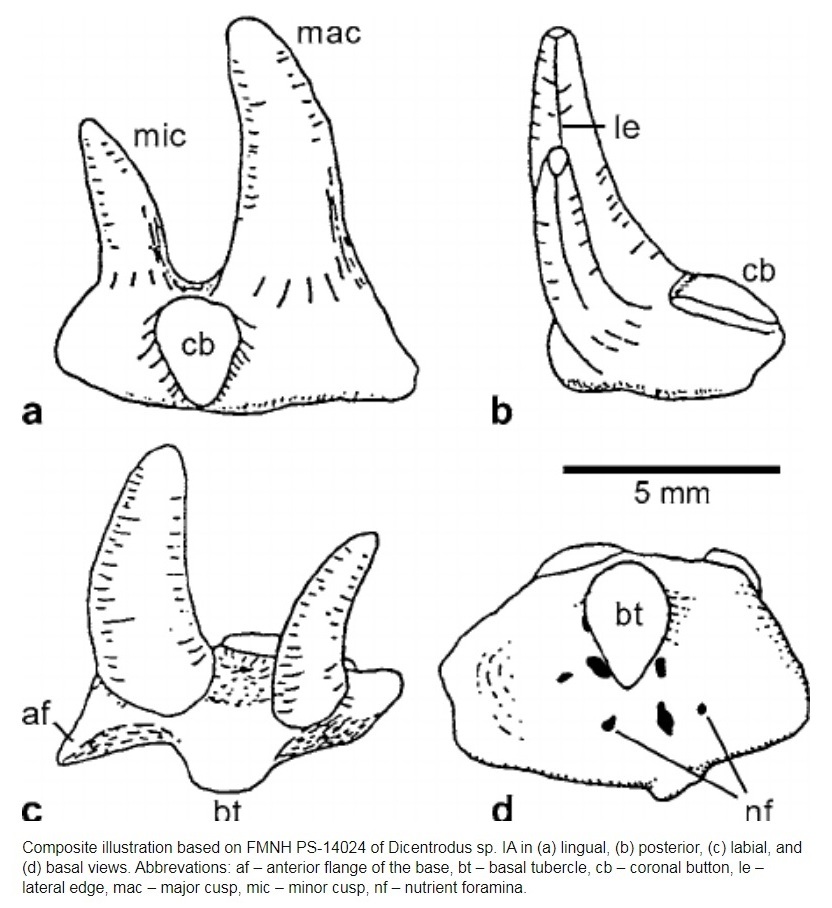
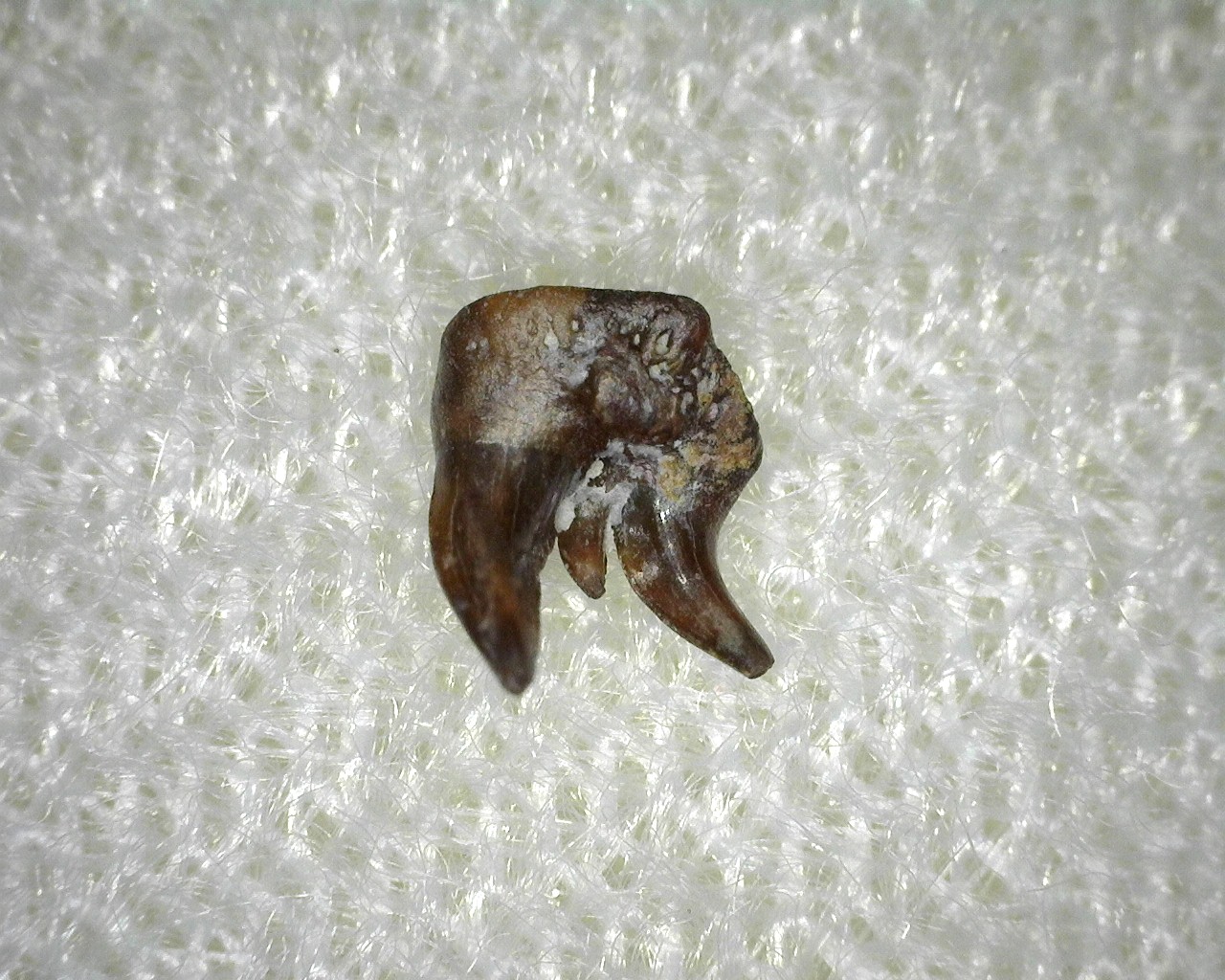
Семейство Диплодоселахиды † Diplodoselachidae Dick, 1981, Род Дицентродусы — † Dicentrodus Traquair, 1888 (известен из каменноугольного периода) состоит из одного вида — Dicentrodus bicuspidatus Traquair, 1881.
У Dicentrodus были двустворчатые зубы с плоским основанием и беззубчатыми или мелко-зубчатыми, лабиолингвально сжатыми, сильно неравными бугорками. Первоначально известный из поздневизейского — раннего серпуховского яруса Шотландии (Dicentrodus bicuspidatus), теперь известно, что он встречается также в среднем визейском веке в Северной Америке. Это самое раннее обнаружение ксенакантид, зарегистрированное в Северной Америке; их наличие основано на неполных зубах, которые легко отличить от других родов ксенакантид. Межконтинентальное распространение Dicentrodus, как и других родов ксенакантид, можно объяснить растущим объемом данных, которые подтверждают, что ксенакантиды были эвригалинными акулами и не ограничивались пресноводной средой обитания. Dicentrodus bicuspidatus — нектобентосное плотоядное животное. [Dicentrodus (Chondrichthyes: Xenacanthida) from the Early Carboniferous (Visean: upper St Louis Formation) of Iowa, USA]
Семейство Диплодоселахиды † Diplodoselachidae Dick, 1981, Род Диплодоселахи — † Diplodoselache Dick, 1981 (известен из каменноугольного периода) состоит из одного вида — Diplodoselache woodi Dick, 1981.
Новые ископаемые остаки были обнаружены из групп горючих сланцев нижнего карбона (Viséan) в районе Эдинбурга. Они показывают мозаику характеристик, типичных как минимум для двух групп ранних акул. Их самая отличительная особенность — маленькие двустворчатые зубы типа «Diplodus». Вместе с длинным спинным плавником и структурой единственного невысокого спинного шипа образует набор знаков, ранее считавшихся уникальными для ксенакантовых акул позднего палеозоя. Другие особенности, такие как большой равнодолевой хвост и строение грудных плавников, возможно, были унаследованы от предка более типичного палеозойского двужаберного вида, обнаруженного у анакантовых и ктенакантовых акул. Предполагается, что ксенаканты произошли от предков в анакант-ктенакантовом комплексе в начале каменноугольного периода и адаптировались к пресноводной среде. Этот предок, вероятно, обладал сложным кожным скелетом, включающим растущие чешуйки с утолщенным и тонким вогнутым основанием.
Фауна конкреций с рыбой, среди которых была обнаружена акула Diplodoselache woodi, интерпретируется как лагунная по своему происхождению. Акула, вероятно, была нектобентосным плотоядным животным среднего размера (около 0,75–2,0 м в длину), обитавшим в поверхностных водах лагуны.
Семейство Диплодоселахиды † Diplodoselachidae Dick, 1981, Род Гагеноселахи — † Hagenoselache Hampe & Heidtke, 1997 (известен из каменноугольного периода) состоит из одного вида — Hagenoselache sippeli Hampe & Heidtke, 1997. [Oliver Hampe und Ulrich H.J. Heidtke, 1997, Hagenoselache sippeli n. gen. n. sp., an early xenacanthid elasmobranch from the Upper
Carboniferous (Namurian B) of Hagen-Vorhalle (NW-Sauerland/Germany)]
В 1997 был описан новый ксенакантид — Hagenoselache sippeli из намюрского периода северо-западного Зауэрланда (NW Sauerland). Пока только относительно полный, расчленённый экземпляр голотипа. Hagenoselache sippeli во многом отличается от предыдущих известных родов ксенакантообразных, например, по строению плавников или зубному ряду. К сожалению, ни спинной шип, характерный для ксенакантид, ни его положение не возможно определить. Поэтому остается неясным, является ли шип головным или вставленным в область плечевого пояса.
Череп небольшой, длина всего лишь от 1/8 до 1/9 длины всего тела. Зубы трёхвершинные высотой от 2 до 3,5 мм; средний выступ от 3/4 до 4/5 длины боковых выступов. Очертания кончиков примерно закруглённые, с 3—6, реже 8 короткими скульптурными полосами в верхней части. Основание зуба плоское и продольно-овальное, от треугольной до грушевидной формы с лингвадической ориентацией конуса. Верхушка корня обычно с 3—6 отверстиями, иногда сбивает с толку зернистая поверхность и морфологически очень изменчивый венечный бугорок с возможным стержнем. От 3 до 7 отверстий на нижней стороне корня. Базальный бугорок округлый, четко углубленный, округлой формы с возможным стержнем. Боковой угол между коронкой и корнем от 120 ° до 125 °. Коронка и корень из ортодентина, коронковый бугорок из трабекулярного дентина. Дентинные канальцы параллельны с слегка зигзагообразными изгибам на периферии. Плавниковый край спины длинный и равномерно высокий. Analis короткий и широкий, предположительно с 8 просто разделёнными лучевыми костями и без значительного сокращения за счет слияния. Caudalis heterocercus, четко отделённый от дорсальной кости большим эпикаудальным выступом и задним. Hypochordal lebus (ипохордовый лебус) почти вдвое длиннее анального отверстия, поддерживаемый серией длинных двояковыпуклых радиалиев.
Семейство Диплодоселахиды † Diplodoselachidae Dick, 1981, Род Лебахакантусы — † Lebachacanthus Soler-Gijon, 1997 (известен из пермского периода) состоит из одного вида — Lebachacanthus colosseus Heidtke, 2007. [HEIDTKE, U.H.J. (2007) Neue Erkenntnisse über Lebachacanthus (Chondrichthyes: Xenacanthida) aus dem Unteren Rotliegend (Unteres Perm) des südwestdeutschen Saar-Nahe-Beckens. [New discoveries on Lebachacanthus (Chondrichthyes: Xenacanthida) from the Lower Rotliegend (Lower Permian) of the southwest-german Saar-Nahe basin]. Mitteilungen der POLLICHIA, 93: 7–14, 5 fig., 2 tab]
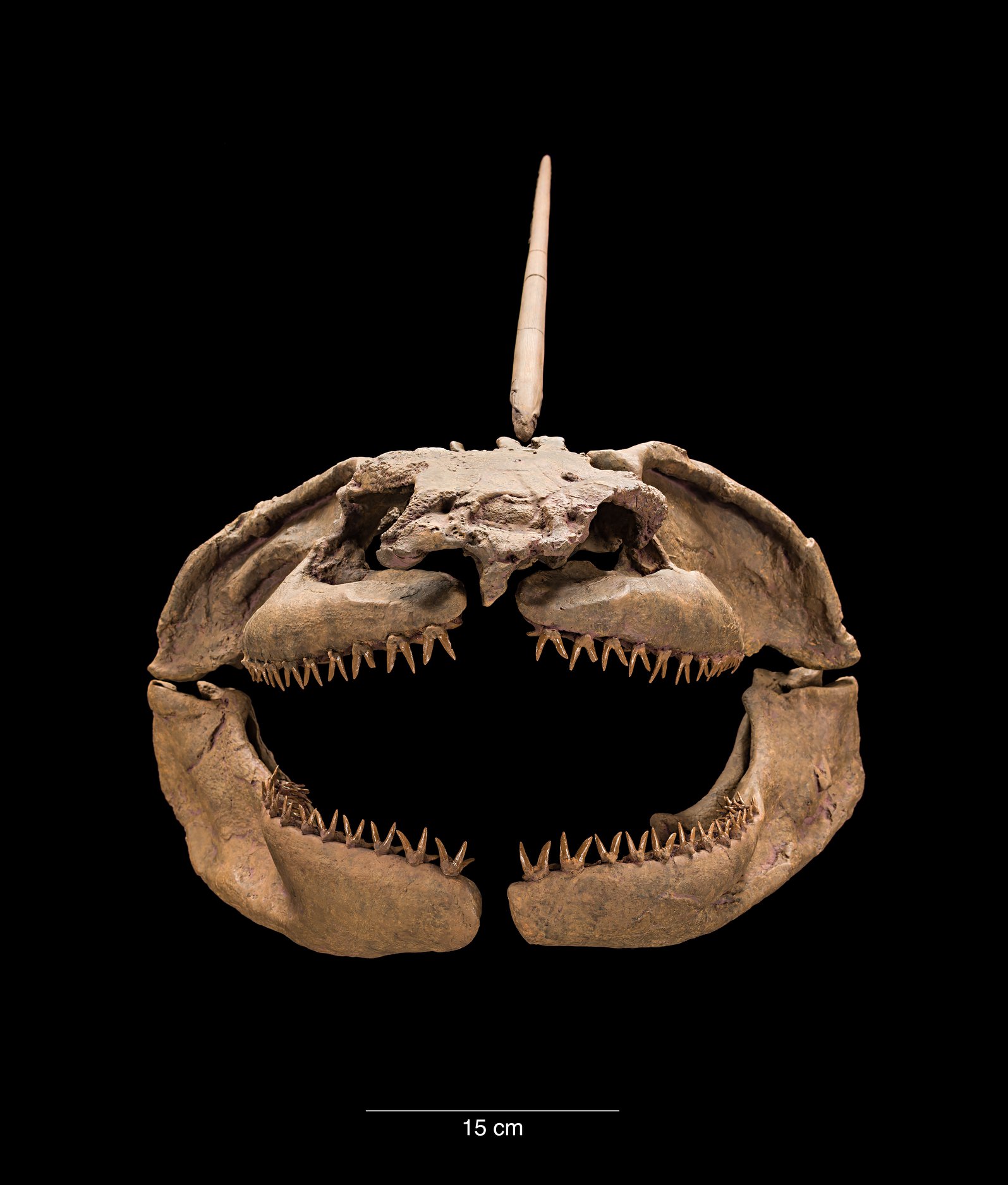
Акула Lebachacanthus процветала в доисторических болотах и заливах Европы и Северной Америки. У неё был очень длинный шип, торчащий из затылка, за которым шёл длинный ленточный спинной плавник, который придавал акуле вид угря. Акулы-самцы имели класперы (claspers), которые сохранились даже в окаменелостях, сразу перед анальными плавниками. Двойные раздвоенные зубы были ещё одной уникальной особенностью этого рода.
Семейство Диплодоселахиды † Diplodoselachidae Dick, 1981, Род Ортакантусы — † Orthacanthus Agassiz, 1843 (известен с каменноугольного, по другим данным с девонского — 407,6 млн лет назад, до мелового периода, от 314,6 до 66,0 миллиона лет назад) состоит из следующих видов: Orthacanthus buxieri Heyler & Poplin, 1989; Orthacanthus compressus Newberry, 1856 [синонимы — Didymodus compressus (Newberry, 1856), Diplodus compressus Newberry, 1856, Diplodus gracilis Newberry, 1856, Diplodus latus Newberry, 1856, Diplodus penetrans Dawson, 1868, Dittodus compressus (Newberry, 1856), Pleuracanthus compressus (Newberry, 1856), Dittodus gracilis (Newberry, 1856), Dittodus latus (Newberry, 1856), Dittodus penetrans (Dawson, 1868)]; Orthacanthus donnelljohnsi Johnson & Thayer, 2009; Orthacanthus gibbosus Agassiz, 1843 [синонимы — Diplodus gibbosus Agassiz, 1843, Dittodus gibbosus (Agassiz, 1843)]; Orthacanthus gracilis Newberry, 1875; типовой вид — Orthacanthus milleri Agassiz, 1843; Orthacanthus minor Agassiz, 1843; Orthacanthus platypternus Cope, 1884 [синонимы — Diacranodus platypternus (Cope, 1884), Didymodus platypternus (Cope, 1884)]; Orthacanthus pustulosus Agassiz, 1843; Orthacanthus texensis Cope, 1888. [L. Agassiz. 1843. Recherches Sur Les Poissons Fossiles. Tome III (livr. 15-16). Imprimerie de Petitpierre, Neuchatel 157-390]
Orthacanthus — род вымерших хрящевых рыб, похожих на акул, из семейства Diplodoselachidae отряда ксенакантообразных. Некоторые систематики выделяют род в монотипическое семейство Orthacanthidae Heyler & Poplin, 1989. По внешнему виду напоминали современных угрей. Отличительной особенностью являлся длинный спинной плавник. В длину достигали 3 метров. Мощные челюсти были покрыты двойным рядом зубов. Были пресноводными хищниками. Обитали в реках и болотах Северной Америки, Европы и Азии.
Зубы Orthacanthus имеют минимум три бугорка, два главных бугорка и промежуточный бугорок, где главные бугры имеют различные зазубрины и имеют сложную морфологию основания. Кроме того, Orthacanthus может быть диагностирован по основным поперечным осям проксимальных концов между бугорками под углом 45 градусов к губному краю основания и часто почти параллельно ему.
Ортакантус жил в морской среде и имел нектобентосную среду обитания, питаясь плотоядной пищей. Многочисленные источники также обнаружили свидетельства каннибализма в рационе ортакантуса и «сыновьего каннибализма», когда взрослый ортакантус охотился на молодых ортакантусов. Учёные, работающие в Дублине (Ирландия), сделали открытие при изучении окаменевших фекалий палеозойской пресноводной акулы Orthacanthus. Крошечные зубы, содержащиеся в их фекалиях, принадлежали совсем юным акулам этого же вида. Эта находка указывает, что 300 миллионов лет тому назад акулы съедали своих детёнышей. Древнейшие хищники нашей планеты растили своих малышей в прибрежных лагунах — тёплых и мелких водоёмах, своеобразных детских садах, которые служили защитой от непогоды и многочисленных опасных обитателей. Однако, взрослые особи практиковали уникальный, но жестокий механизм регуляции численности хищников в экосистеме — поедание своих отпрысков. Это довольно редкое явление в животном мире.
Исследователи подчёркивают, что доисторические акулы пожирали свой молодняк не из-за безжалостного характера или натуры убийцы. Такой каннибализм в природе обычно объясняется тем, что окружающие условия не позволяют выкормить всех детёнышей, которых самка произвела на свет — например, в случае дефицита пищи. Таким образом, если бы акулы оставляли в живых всё своё потомство, то оно было бы обречено на медленную голодную смерть. Ископаемые доказательства, подтверждающие акулий каннибализм, были найдены в копролитах (окаменевшие фекалии) уникальной спиральной формы в Нью-Брансуике, Канада, на участке Минто. Экскременты были идентифицированы как принадлежащие Orthacanthus благодаря своей форме. Штопорную форму прямой кишки этой древней акулы трудно спутать с чем-то другим. Вид окаменевших фекалий позволил легко идентифицировать вид акулы. Окаменевшие копролиты полны зубов маленьких акул, подтверждающие тот факт, что эти акулы на самом деле питались собственными детёнышами, что называется «сыновий каннибализм» (filial cannibalism).
Палеобиогеографическое распространение Orthacanthus platypternus предполагает онтогенетическое разделение местообитаний. Теория онтогенетической ниши предсказывает, что акулы могут изменить свою среду обитания или диету, чтобы поддерживать оптимальные темпы роста или улучшить компромисс между риском смертности и ростом. В то время как более мелкие особи, вероятно, жили в более мелких водах, таких как небольшие пруды и ручьи прибрежной равнины, более крупные особи, вероятно, жили в более глубоких водах, таких как флювио-озерные (реки и озера) и окраинные морские районы.
Самый старый из известных экземпляров Orthacanthus, Diplodus problematicus (?), был найден в Нью-Брансуике, Канада, в период от 407 до 393 миллионов лет назад. Другие образцы были найдены в таких местах, как США, Великобритания, Польша и Франция.
В отрывке из фильма «Акулы-каннибалы (Cannibal Sharks)» кинокомпании National Geographic рассказывается о проявлении акульего каннибализма ещё 300 миллионов лет назад у представителя рода Ортакантусы — † Orthacanthus Agassiz, 1843 вымершего отряда † Xenacanthiformes Berg, 1955.
Семейство Ксенакантиды † Xenacanthidae Fritsch, 1889, Род Аканторлеурусы, Аканторлеврусы — † Acanthopleurus Agassiz, 1843 (известен из раннего олигоцена) состоит из одного вида — Acanthopleurus serratus Agassiz, 1843. [L. Agassiz. 1843. Recherches Sur Les Poissons Fossiles. Tome I (livr. 18). Imprimerie de Petitpierre, Neuchatel xxxii-188]
По другим данным в род Acanthopleurus входят виды Acanthopleurus serratus Agassiz, 1843, и Acanthopleurus collettei Tyler, 1980 из олигоцена кантона Гларус (Glarus), Швейцария. Третий описанный вид Acanthopleurus (Cephalacanthus) trispinosus Ciobanu, 1977 из олигоцена Карпат Румынии, ранее принадлежавший к семейству Dactylopteridae, считался ювенильным экземпляром Acanthopleurus, хотя еще не известно, принадлежит ли он к одному из двух описанных видов или новому виду.
Семейство Ксенакантиды † Xenacanthidae Fritsch, 1889, Род Аганодусы — † Aganodus Owen, 1867 состоит из следующих видов: Aganodus apicalis Owen, 1867, синоним — Dittodus apicalis (Owen, 1867); Aganodus undatus Owen, 1867, синоним — Dittodus undatus (Owen, 1867).
Семейство Ксенакантиды † Xenacanthidae Fritsch, 1889, Род Комсакантусы — † Compsacanthus Newberry, 1856 состоит из одного вида — Compsacanthus laevis Newberry, 1856, синоним — Pleuracanthus laevis (Newberry, 1857) .
Головной шип зазубрен сзади, поперечное сечение его овальное, у верхушки круглое. Челюстные зубы по краям зазубренные. Их среднее острие короткое (1/3 наружного). Жаберные зубы с многочисленными вершинами, беспорядочно расположенными на основании. Плавники без шипов, грудные длинные, заостренные.
Семейство Ксенакантиды † Xenacanthidae Fritsch, 1889, Род Диакранодусы — † Diacranodus Garman, 1885 (известен из пермского периода) состоит из одного вида — Diacranodus texensis Cope, 1888, синоним — Pleuracanthus laevis (Newberry, 1857).
Тип рода — Didymodus texensis Соре, 1883; пермь США. Известны только череп и зубы. Носовые и слуховые капсулы и посторбитальный отросток хорошо развиты. Foramen magnum на нижней стороне черепа, в области базисфеноида. Зубы цельнокрайние, как у Triodus.
Семейство Ксенакантиды † Xenacanthidae Fritsch, 1889, Род Файолии — † Fayolia Renault & Zeiller, 1884 (известен с девонского до триасового периода) состоит из следующих видов: типовой вид — Fayolia dentata Renault & Zeiller, 1884; Fayolia sterzeliana Weiss, 1887.
Файолия преимущественно известен из пресноводных отложений со стратиграфическим диапазоном от позднего девона до триаса. Новый вид, Fayolia sharovi, был описан в 2011 году из озёрных отложений среднего триаса формации Мадыген (Madygen Formation) в Кыргызстане, Центральная Азия.
Мадыгенская формация представляет собой толщу горных пород триасового возраста. Она состоит из земных, озёрных и речных отложений. Из формации найдено много ископаемых позвоночных, в том числе хрящевых рыб и необычных триасовых рептилий, таких как Шаровиптерикс и Лонгисквама. Существует также большое разнообразие ископаемых насекомых из этой местности, причем заметным примером является Titanoptera.
Яичные капсулы Fayolia sharovi достигали общей длины 7 см и характеризуются веретенообразным телом, состоящим из двух широких спирально закрученных по часовой стрелке полос. Края полос параллельны отчетливой линией рубца, перекрытой широким фланцем, сопровождающим обе полосы. Поскольку ксенакантообразные акулы обычно считаются наиболее вероятными продуцентами Fayolia, появление капсул типа Fayolia подразумевает присутствие еще неизвестной акулы-ксенакантиды в фауне Мадыгена. Другим признаком его присутствия являются тонкие заострённые дермальные зубчики сродства ксенакантидам. Учитывая массовое появление молодых зубов и яичных коробочек в районе исследования, было высказано предположение, что ксенакантидные акулы периодически заселяли прибрежные зоны озера Мадыген для нереста. Небольшое количество взрослых особей указывает на разделение среды обитания молодых и взрослых особей, поэтому место исследования интерпретируется как питомник акул. Наличие различных видов яйцекладущих акул также указывает на некоторую степень временного разделения. Таким образом, стратегии откладки яиц (изменение среды обитания, верность местам), полученные от акул Мадыгена, удивительно похожи на стратегии современных их родственников, что позволяет предположить, что репродуктивные модели, наблюдаемые у современных акул, возникли задолго до кайнозоя.
Семейство Ксенакантиды † Xenacanthidae Fritsch, 1889, Род Лепакантусы — † Lepacanthus Arthaber, 1896 (известен из триасового периода, от 247,2 до 242,0 миллиона лет назад) состоит из одного вида — Lepacanthus gracilis Arthaber, 1896. [G. Arthaber. 1896. Die Cephalopodenfauna der Reiflinger Kalke. Beiträge zur Paläontologie und Geologie Österreich-Ungarns und des Orients 10:1-111]
Типовой образец найден в разрез Артабер (Arthaber section) Тифенграбена (Tiefengraben), который находится в карбонатных известняках пельсонского / иллирийского происхождения в формации Гутенштейн (Gutenstein Formation) в Австрии. Lepacanthus gracilis — нектобентосное плотоядное животное.
Семейство Ксенакантиды † Xenacanthidae Fritsch, 1889, Род Лофакантусы — † Lophacanthus Stock, 1880 (известен из каменноугольного периода) состоит из одного вида — Lophacanthus taylori Stock, 1880.
Вид назван в честь Джозефа Тейлора (Joseph Taylor, 1840 — 1902), который был шахтером и микроскопистом-любителем, пользовавшимся хорошей репутацией за подготовленные образцы для микроскопа. Томас Сток из отдела естественной истории Музея науки и искусства (Museum of Science and Art) в Эдинбурге писал в 1880 году: «Среди посылки с остатками рыбы, отправленной мне несколько месяцев назад мистером Джозефом Тейлором из Шир-Мур (Shire Moor), Нортумберленд (Northumberland), был шип, который, по его словам, был новым образцом. После тщательного изучения экземпляра и ознакомления со всей доступной мне литературой по этому предмету я пришёл к выводу, что шип не только специфически новый, но и должен рассматриваться как тип нового рода ... Небольшой кусочек около 7 миллиметров был удален мистером Тейлором с намерением сделать из него микроскопический срез ... Образец кажется уникальным. Пока открытие не проливает свет на его истинное сходство, он должен оставаться типом нового рода, который я предлагаю назвать Lophacanthus; и к нему я добавляю конкретное имя Тейлори, в честь первооткрывателя».
Семейство Ксенакантиды † Xenacanthidae Fritsch, 1889, Род Муреодонтусы — † Mooreodontus Hampe & Schneider, 2010 (известен из триасового периода) состоит из следующих видов: Mooreodontus indicus (Jain, 1980); Mooreodontus jaini Bhat, Ray & Datta, 2018; Mooreodontus moorei (Woodward, 1889) [синонимы — Diplodus moorei, Dittodus moorei, Triodus moorei, Xenacanthus moorei]; Mooreodontus parvidens (Woodward, 1908) [синонимы — Pleuracanthus parvidens, Xenacanthus parvidens]. [O. Hampe and J. Schneider. 2010. Mooreodontus name, in Chondrichthyes, Paleozoic Elasmobranchii: Teeth. Handbook of Paleoichthyology 3D:54]
Почти полный образец из клинкерных карьеров в Сент-Питерсе (St Peters), Сидней, Австралия, который Вудворд (1908) идентифицировал как Pleuracanthus parvidens. В последнее время Ginter et al. (2010) переименовал Pleuracanthus parvidens в Mooreodontus parvidens из-за морфологического сходства австралийских зубов с Mooreodontus, уже известным из триасовых отложений в Бразилии, Европе и США (Turner 2011). Дальнейшая работа с этими образцами указывает на то, что в Австралии присутствует несколько видов, у которых есть морфологические различия в брюшных класперах (claspers), которые могут быть диагностическими признаками (Schultze & Soler-Gij 2004, Turner 2011).
Сообщается о первом обнаружении разнообразного комплекса ксенакантов из формации Тики (Tiki Formation) из верхнего триаса в Индии, основанного на множестве хорошо сохранившихся отдельных зубов. На основании отличительной морфологии зубов описаны два вида рода Mooreodontus: Mooreodontus indicus и новый вид, Mooreodontus jaini. Новый вид диагностируется на основании трёхстворчатой коронки, содержащей два толстых, слегка расходящихся боковых выступа, указывающих в одном направлении, высокий средний бугорок, угол между основанием коронки почти 90 °, большой, округлый, апикальный узел с несколькими отверстиями и множественными отверстиями. На всех бугорках по 8—9 грубых вертикальных крист (гребней). В коллекции присутствует зубная аномалия в виде частичного четырёхкуспидального ксенакантида. Другая группа зубов ксенакантид имеет двустворчатые коронки с двумя вертикальными асимметричными бугорками, при этом мезиальный бугорок толще дистального, и постоянно отсутствуют срединный куспид. Такая отчётливая морфология двустворчатого зуба обычно присутствует у палеозойских форм и впервые обнаружена в позднем триасе. Считается, что он принадлежит к новому таксону Tikiodontus asymmetricus nov. gen., неопределённого семейства. Отличительная гистология зубов также различает два индийских рода Mooreodontus и Tikiodontus nov. gen. из других таксонов ксенакантид. Индия занимает уникальное положение с точки зрения фауны пресноводных акул позднего триаса, так как она демонстрирует явные лавразийские сходства акул. (Лавразия — древний северный континент, из двух континентов (южный — Гондвана), на которые распался протоконтинент Пангея в мезозое. Позже, от 135 до 200 миллионов лет назад, Лавразия распалась на Евразию и Северную Америку. Лавразия включала большую часть территорий, которые ныне составляют континенты Северного Полушария.) Эти пресноводные акулы редко встречались в других частях суши Гондваны.
Семейство Ксенакантиды † Xenacanthidae Fritsch, 1889, Род Охлодусы — † Ochlodus Owen, 1867 (известен из каменноугольного периода, от 306,95 до 303,4 миллиона лет назад) состоит из одного вида — Ochlodus crassus Owen, 1867 [синонимы — Diplodus equilateralis, Diplodus gibbosus, Dittodus gibbosus, Dittodus parallelus, Lophacanthus taylori, Ochlodus crassus, Orthacanthus cylindricus, Orthacanthus (Orthacanthus) cylindricus, валидное название — Orthacanthus gibbosus Agassiz, 1843, Pleuracanthus cylindricus, Pleuracanthus (Diplodus) equilateralis, Pleuracanthus (Lophacanthus) taylori, Pleuracanthus (Xenacanthus) gibbosus]. [L. Agassiz. 1843. Recherches Sur Les Poissons Fossiles. Tome III (livr. 15-16). Imprimerie de Petitpierre, Neuchatel 157-390]
Orthacanthus gibbosus Agassiz, 1843 (Ochlodus crassus Owen, 1867) — нектобентосное плотоядное животное. Типовой образец: BMNH.497, зуб. Его типовое местонахождение — Сильвердейл (Silverdale), Южный Стаффордшир (South Staffordshire), который находится на московском горизонте земли в Соединенном Королевстве.
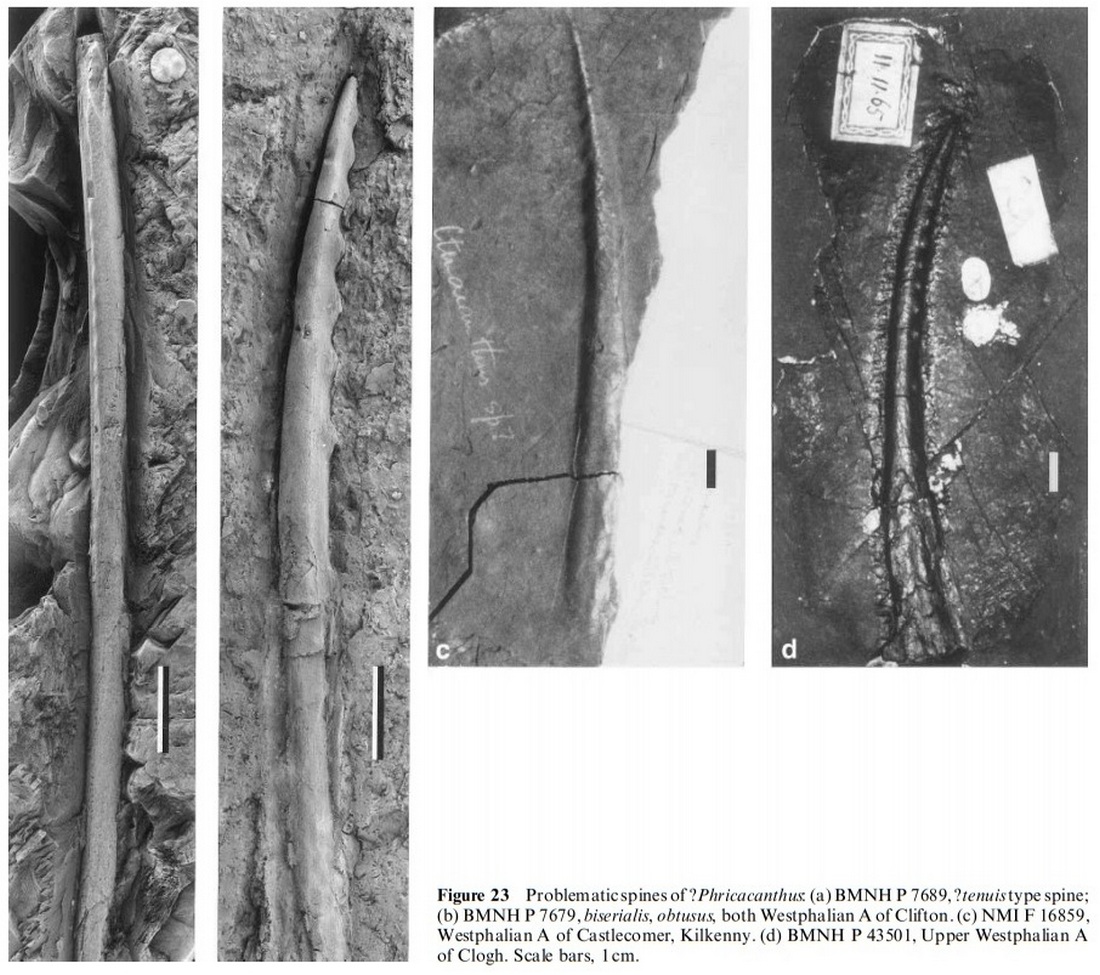
Семейство Ксенакантиды † Xenacanthidae Fritsch, 1889, Род Фрикакантусы — † Phricacanthus Davis, 1879 (известен из каменноугольного периода) состоит из одного вида — Phricacanthus biserialis Davis, 1879. [L. Agassiz. 1843. Recherches Sur Les Poissons Fossiles. Tome III (livr. 15-16). Imprimerie de Petitpierre, Neuchatel 157-390]
Шип вида Phricacanthus biserialis, описанного Дэвисом (1879, pl. X), был обнаружен в угольных месторождениях Йоркшира в северной Англии. Это плавно изогнутый элемент с регулярно и чрезвычайно широкими выступами на вентральной стороне, длиной 10,7 см (рис. 23 б). Поверхность затемнена тонким слоем угля. Широкие промежутки между выступами неизвестны ни у одного другого ксенакантида, описанного до сих пор. Дэвис обсуждал сходство фрикаканта с ортакантусом. Позже Вудворд (1889) отнес этот образец к Orthacanthus cylindricus (? Orthacanthus gibbosus). Морфология колючки Orthacanthus gibbosus сильно отличается от Phricacanthus: шипы большие, прямые, с двумя рядами выступов с многочисленными мелкими зубчиками. В замешательстве Дэвис (Davis, 1892) дал ему новое название вида, потому что biserialis была занята, и поместил Phricacanthus в Pleuracanthus, теперь как Pleuracanthus obtusus. Зидек (1993) также считал род Фрикакантусы синоним Orthacanthus с неопределенными видами. Незначительное сходство можно увидеть с шипами Orthacanthus denticulatus. Однако у Orthacanthus denticulatus шипы более тонкие и проксимальный конец не увеличивается в диаметре, как у Phricacanthus.
Современное состояние знаний и отсутствие подходящего задокументированного материала не позволяет с уверенностью сказать, является ли Фрикакантус ксенакантидом. Связь с ктенакантоидными эластожаберными так же не может быть исключена. Подобные шипы известны из вестфальских шипов в Ирландии (рис. 23 в, г). Известно лишь несколько скелетов британских ксенакантидов. Это потому, что большая часть материала была собрана
как случайное открытие при добыче продуктивного угля. Фрагментарные скелеты пластиножаберных ксенакантид действительно существуют в дополнение к более или менее полному скелету Diplodoselache woodi, обнаруженному в 1972 году, с дополнительным материалом собраны в 1974 г.
Семейство Ксенакантиды † Xenacanthidae Fritsch, 1889, Род Платиакантусы — † Platyacanthus Fritsch, 1889 (известен из каменноугольного периода) состоит из одного вида — Platyacanthus ventricosus Fritsch, 1889.
Фрич (1889) описал шип стефанианского времени из Богемии, как Platyacanthus ventricosus, который Heidtke (1998) по уважительным причинам поместил в род Lebachacanthus, из-за его внешнего вида (максимальное соотношение ширины / длины 1:7,7) и отсутствием хорошо выраженной продольной штриховки. На этом шипе нет зубчиков, что иногда встречается у Lebachacanthus senckenbergianus. Из недавних исследований Soler-GijoAn (1999) по гистологии и механизмов роста, было подтверждено, что зубчики были добавлены проксимально в ходе эволюции, как независимые дермальные элементы в ходе онтогенеза шипа. Зубчики вырастали и минерализовались в зародышевой зоне кожи. Когда шип прорывался сквозь кожу, зубчики срастались с поверхностью колючки. Тем не менее, может иметь место различная длина зубцов, промежутков в зучатом ряду или даже полное отсутствие зубцов (асинхронное расположение, циклические изменения в зубном ряду, скорость роста).
В 1978 году Зидек описал † Platyacanthus avirostratus Zidek, 1978 из формации Гарбер (нижняя пермь) на юго-западе Оклахомы, США. [Zidek, J. (1978) New Chondrichthyan Spines from the Late Paleozoic of Oklahoma. Journal of Paleontology, 52 (5): 1070–1078]
Семейство Ксенакантиды † Xenacanthidae Fritsch, 1889, Род Плевракантусы — † Pleuracanthus Agassiz, 1837 (известен из каменноугольного и пермского периодов, от 314,6 до 235,0 миллиона лет назад) состоит из следующих видов: Pleuracanthus arcuatus Newberry, 1856; Pleuracanthus biserialis Newberry, 1856; Pleuracanthus cylindricus Agassiz, 1843; Pleuracanthus decheni Goldfuss, 1847; Pleuracanthus dilatatus Newberry, 1856; Pleuracanthus levissimus Agassiz, 1837; Pleuracanthus quadriseriatus Cope, 1877; Pleuracanthus robustus Davis, 1880. [L. Agassiz. 1837. Recherches Sur Les Poissons Fossiles. Tome III (livr. 8-9). Imprimerie de Petitpierre, Neuchatel viii-72]
Род Pleuracanthus палеозойских акул карбона и нижней перми Европы и Северной Америки, имеющий субтерминальный рот, длинный спинной плавник и сильный зубчатый шип на затылке. Состав рода Pleuracanthus сильно различается у разных систематиков, некоторые из них синонимизируют его с родом Xenacanthus.
Угольные месторождения Йоркшира породили колючки нескольких виды этого рода, многие из которых находятся в прекрасной сохранности, но другие показывают все стадии эрозии, некоторые из них тупоконечные и лишены зубчики, а в последних экземплярах после удаления поверхностного гладкого слоя появляются шипы волокнистой текстуры. Compsacanthus triangularis Davis; Compsacanthus major Davis; Phricacanthus biserialis Davis; и следующие виды Pleuracanihus, а именно, ereetus Davis; planus Agassiz; wardi Davis; pulchellus Davis; по мнению Эдгара Д. Веллберна, были описаны на основании шипов Pleuracanihus на разных стадиях эрозии. Он же выделял в роде Pleuracanihus следующие действительные виды (устаревшая информация 1901 года): Pleuracanihus imvissimus Agassiz, 1837; Pleuracanihus robustus Davis, 1880; Pleuracanihus cylindricus Agassiz, 1843; Pleuracanihus alatus Davis, 1880; Pleuracanihus alternidentatus Davis, 1880; Pleuracanihus tenuis Davis, 1880; Pleuracanihus denticulatus Davis, 1880; Pleuracanihus horidus Traquair, 1881; Pleuracanihus wardi Davis, 1880 ?. [Edgar D. Wellburn Proceedings of the Yorkshire Geological and Polytechnic Society 1901, v.14; p159-174. On the Fish Fauna of the Yorkshire Coal Measures]
Семейство Ксенакантиды † Xenacanthidae Fritsch, 1889, Род Пликатодусы — † Plicatodus Hampe, 1995 (известен в Европы из каменноугольного и раннепермского периодов, от 307,0 до 295,0 миллиона лет назад) состоит из следующих видов: типовой вид — Plicatodus jordani (Hampe, 1995); Plicatodus plicatus (Fritsch, 1879) [синоним — Orthacanthus plicatus (Fritsch, 1879)];
Plicatodus sp.. [Ginter, M., Hampe, O., Duffin, C., 2010. Handbook of Paleoichthyology Volume 3D: Chondrichthyes: Paleozoic Elasmobranchii: Teeth. in Schultze, H. (ed.) Handbook of Paleoichthyology. Verlag Dr. Fredich Pfeil-Munchen.]
Типовой вид — Plicatodus jordani был описан Оливером Хэмпом в 1995 году из бассейна Саар–Наэ (Saar-Nahe) на юго-западе Германии. Этот бассейн расположен к югу от гор Хунсрюк (Hunsrück) между реками Наэ и Саар. Агрегация отложений в бассейне началась в конце карбона и ранней перми в рамках варисканской складчатости. [Plicatodus jordani n. g., n. sp., a new xenacanthid shark from the Lower Permian of Europe (Saar-Nahe Basin, Germany) by Oliver Hampe, 1995.]
Окаменелости Plicatodus обычно состоят только из отдельных зубов, за одним исключением: частичный скелет, включающий голову, спинной отдел позвоночника и грудной пояс. Типовой вид этого рода, Plicatodus jordani, был описан на основе материала пермского возраста (а именно ассельская часть цисуральского яруса), обнаруженного в верхних слоях формации Одернхейм (Odernheim Formation) в бассейне Саар-Наэ (Saar–Nahe Basin): моласса пресноводных и мелководных морских глубин. Термин «моласса» относится к песчаникам, сланцам и конгломератам, которые образуют в наземных или морских мелководных отложений в передней части растущих горных цепей.
Plicatodus plicatus первоначально был описан в качестве члена рода Orthacanthus по Карлу фон Фрич (Karl von Fritsch) 1879 год. Окаменелости найдены в формации Сланы (Slany Formation) в Раковницком бассейне (Rakovnik Basin) в Чехии из касимовского временного диапозона (карбон).
Plicatodus sp., найденный из гжельского временного диапозона (карбон) в Нижней Австрии, не определен. Хотя он может представлять неописанный третий вид данного рода, и может происходить от Plicatodus jordani или Plicatodus plicatus.
Семейство Ксенакантиды † Xenacanthidae Fritsch, 1889, Род Птернодусы — † Pternodus Owen, 1867 (известен из каменноугольного периода) состоит из одного вида — Pternodus produetus Owen, 1867.
Pternodus produetus — нектобентосное плотоядное животное.
Семейство Ксенакантиды † Xenacanthidae Fritsch, 1889, Род Триодусы — † Triodus Jordan, 1849 (известен в Европы из каменноугольного, пермского и триасового периодов, от 318,1 до 212,0 миллиона лет назад) состоит из следующих видов: Triodus elpia Johnson & Thayer, 2009; Triodus moorei Woodward, 1889; Triodus richterae Pauliv et al., 2017 и типовой вид — Triodus sessilis Jordan, 1849.
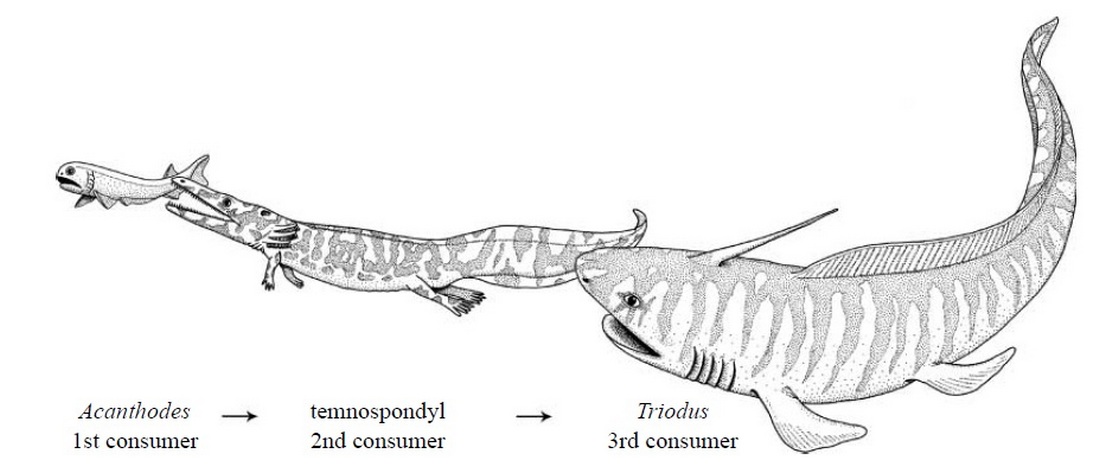
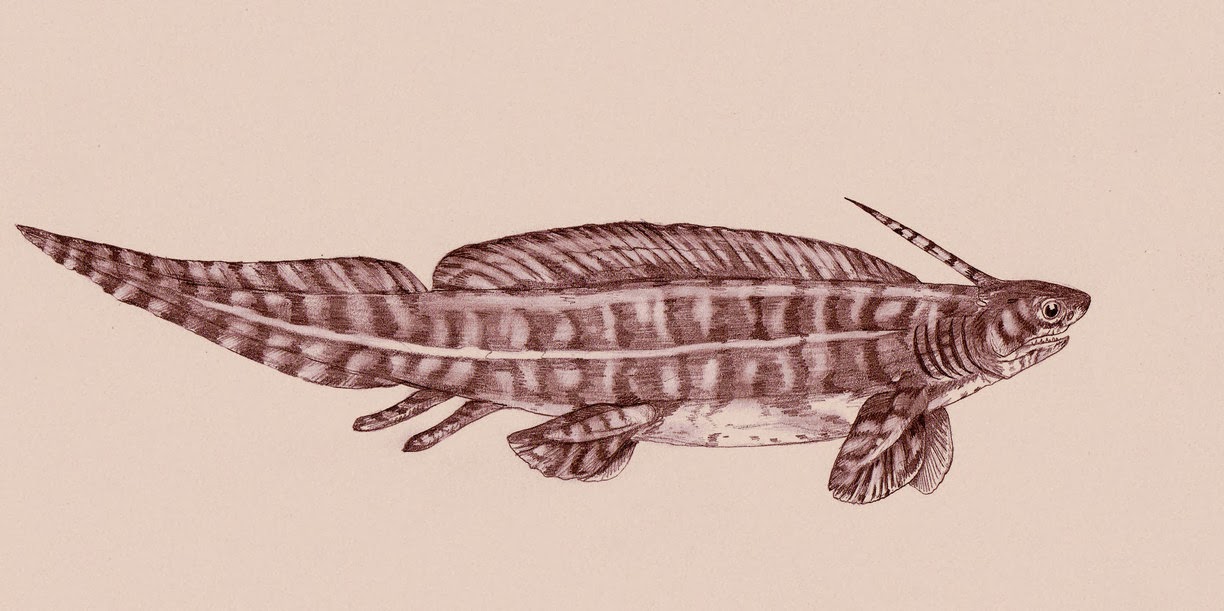
Эта рыба имела вытянутую форму, а тело отдалённо напоминало тело угря. Как и многие другие его близкие родственники, Triodus обладал длинным низким спинным плавником, который непосредственно соединялся с хвостовым плавником. Голова с короткой мордой и большими глазницами была наделена чем-то вроде заднего шипа прямо перед длинным спинным плавником. В отличие от многих подобных форм, таких как Xenacanthus, Triodus имел особенно тонкое, напоминавшее угря, тело, а размеры были небольшими: обычно его длина не превышала полуметра. Для него были характерны трёхконечные зубы (отсюда и название Triodus). Triodus, должно быть, был небольшим пресноводным хищником; в некоторых окаменелостях обнаружены остатки личинок земноводных, принадлежащих к родам Archegosaurus и Glanochthon. В свою очередь, на Triodus, скорее всего, охотились более крупные акулы, такие как Ортакантус (Orthacanthus) и Ксенакантус (Xenacanthus).
Род Triodus был впервые описан в 1849 году Джорданом; Наиболее хорошо сохранившиеся окаменелости происходят из каменноугольно-пермских бассейнов Германии, среди них есть вид Triodus sessilis, известный по множеству полных образцов. Другие окаменелости происходят из верхнего карбона Аризоны (Triodus elpia) и из верхнего триасаСоединенных Штатов (Triodus moorei, найденный в Аризоне, Нью-Мексико и Техасе). Таким образом, Triodus был очень долгоживущим родом с консервативной морфологией, который оставался почти неизменным в течение примерно 100 миллионов лет. Triodus принадлежал к группе ксенакантид, группе Elasmobranchii с удлинённым телом и характерными зубами.
Triodus Jordan, 1849 (Pleuracanthus Agassiz, 1837, nom. praeocc). Тип рода — Triodus sessilis; нижняя пермь Германии. Зубчики головного шипа в нижней части его боковых сторон, поперечное сечение шипа эллипсоидное. Челюстные зубы тонкие, по краям незазубренные, с остриями почти одинаковой длины. Жаберные зубы в виде трезубца. Плавники без шипов, грудные длинные, заострённые, как у Orthacanthus (рис. 23). Это были пресноводные рыбы, окаменелости были обнаружены в Chinle Formation и Black Prince Limestone в Аризоне, Petrified Forest Formation в Нью-Мексико и в Tecovas Formation в Техасе, США, и в западной Европе.
В 2017 году, новый вид Triodus richterae был описан из Rio do Rasto Formation в Бразилии. Triodus — хорошо известный род Xenacanthiformes, ранее отмеченный от позднего башкирского (нижний пенсильванский, каменноугольный) до среднего артинского (цисуральский, пермский) веков, в основном из лауразийских отложений (Европа и США). Впервые этот род из Западной Гондваны зарегистрирован по изолированным зубам, которые относят к Triodus richterae sp. nov. Новый вид был обнаружен с другими зубами акул (другие xenacanthiforms и, возможно, euselachian), зубами и чешуей палеонискоидов (palaeoniscoid), зубами лабиринтодонтов (labyrinthodont), костными останками четвероногих, макроскопическим углем и фрагментами листьев. Этот комплекс окаменелостей был собран в слое конгломератов на участке Барро-Алту (муниципалитет Сан-Габриэль, штат Риу-Гранди-ду-Сул, Бразилия) с предполагаемым капитанским возрастом в пачке Морро Пеладу, формация Риу-ду-Расту, бассейн Парана. У нового вида зубы с почти овальным основанием, аборальная поверхность имеет гладкую вогнутость и базальный бугорок от округлой до подковообразной формы. Венечная поверхность этих зубов имеет трёхвершинную коронку, коронковая вершина ромбовидной формы с закругленными краями с лингвально направленным стержнем. Несколько оральных отверстий расположены преимущественно на язычном крае основания и по бокам язычного стержня. Боковые бугры несут различное количество неразветвлённых вертикальных гребешков, распределённых от вершины к их проксимальной части, что придаёт поперечному сечению этих бугорков звездообразную форму. Микроструктурно эти зубы, как основание, так и бугорки, состоят из ортодентина с открытой полостью пульпы. Triodus richterae sp. nov. представляет собой самый молодой вид Triodus, учитывая, что он происходит из капитанских (поздних гваделупских) отложений, которые как минимум на 15 млн лет моложе Triodus kraetschmeri, ранее самого молодого вида этого рода. Интерпретация осадконакопления, а также совокупность окаменелостей в типовой местности нового вида и формации Риу-ду-Расто в целом указывают на еще одну находку пресноводных акул ксенакантид. [Victor E. Pauliv; Agustin G. Martinelli; Heitor Francischini; Paula Dentzien-Dias; Marina B. Soares; Cesar L. Schultz; Ana M. Ribeiro (2017). "The first Western Gondwanan species of Triodus Jordan 1849: A new Xenacanthiformes (Chondrichthyes) from the late Paleozoic of Southern Brazil". Journal of South American Earth Sciences. 80: 482–493.]
Время: пермь, карбон. Место: Европа, Германия. Описанные виды: Triodus kraetschmeri, Triodus lauterensis, Triodus obscurus, Triodus palatinus, Triodus sessilis. Новые описания видов: Triodus kraetschmeri, Triodus lauterensis, Triodus obscurus, Triodus palatinus.
Резюме: Описаны зубы пяти видов в Ротлигенде (Rotliegend) в районе Саар Нахе (Saar Nahe): Triodus sessilis Jordan 1849, Triodus lauterensis n. sp., Triodus palatinus n. sp., Triodus obscurus n. sp. и Triodus kraetschmeri n. sp. Представители рода Triodus имеют типичный набор зубов для захвата добычи, который характеризуется относительно тонкими бугорками с определённым для вида количеством вертикальных гребешков и длиной срединного бугорка. Виды также различаются расположением крист, положением базального бугорка и углом между коронкой и корнем при виде сбоку. На основе девяти признаков пять видов анализируются кладистическим способом. Эта предварительная диаграмма синапоморфов превращена в кладограмму, которая, однако, все ещё оставляет без ответа многие вопросы. [HAMPE, O. (1989) Revision der Triodus-Arten (Chondrichthyes: Xenacanthida) aus dem saarpfälzischen Rotliegenden (Oberkarbon - Perm, SW-Deutschland) aufgrund ihrer Bezahnung. Paläontologische Zeitschrift, 63 (1/2): 79–101, 7 fig.,]
Семейство Ксенакантиды † Xenacanthidae Fritsch, 1889, Род Вурдигнерии — † Wurdigneria Richter, 2005 (известен в Европы из каменноугольного, пермского и триасового периодов, от 318,1 до 212,0 миллиона лет назад) состоит из следующих видов: типовой вид — Wurdigneria obliterata Richter, 2005; Wurdigneria pricei Würdig-Maciel, 1975 и Wurdigneria santosi Würdig-Maciel, 1975. [RICHTER, M. (2005) A new xenacanthid shark (Chondrichthyes) from the Teresina Formation, Permian of the Parana Basin, southern Brazil. Revista Brasileira de Paleontologia, 8 (2): 149–158]
Казанско-татарская терезинская формация (Kazanian/Tartarian Teresina Formation) бассейна Парана до сих пор является наиболее продуктивной стратиграфической единицей в Бразилии, что касается палеозойских акул, хотя никаких сочленённых материалов пока не обнаружено. Зубы, шипы плавников и кожные чешуйки местами многочисленны, особенно в бугристых отложениях (tempestites). Биогеохимические данные указывают на пресноводное происхождение этой фауны. Новые материалы зубов хондрихтида (chondrichthyan), происходящие из одного места этого образования в муниципалитете Сан-Габриэль (São Gabriel) в штате Риу-Гранди-ду-Сул (Rio Grande do Sul), описаны и приписаны новой акуле-ксенакантиду, а именно Wurdigneria obliterata gen et sp. nov. У этого нового вида трёхвершинные зубы, у которых центральный бугорок составляет менее одной трети ширины и менее половины высоты боковых бугров. Боковые бугры ланцетовидные в поперечном сечении, округлые в проксимальном направлении, прямые вверх по отношению к основанию и лишены каких-либо зазубрин. Боковые бугры могут нести пару вертикальных неразветвленных крист. Основание округлое или треугольное, состоит из остеодентина. Верхушка коронки вздута, обычно круглая или, в редких случаях, слегка приплюснута на вершине. Имеется не менее трёх крупных базальных и язычных отверстий. Базальный бугорок больше, меньше или равен длине основания боковых бугорков. Гистологически боковые выступы Wurdigneria obliteratagen состоят из ортодентина снаружи и остеодентина, в каналах пульпы. Эмелоидная ткань отсутствует. Судя по сравнительной анатомии зубов, это была хищная акула, которая могла достигать около 100 мм в длину.
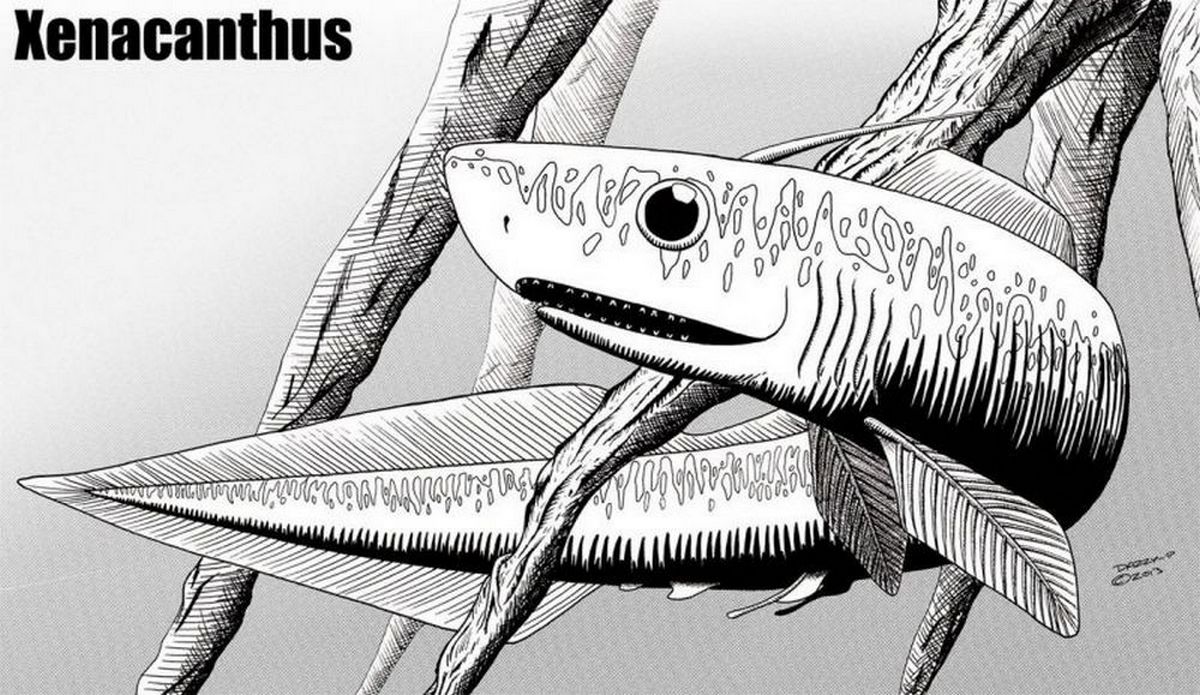
Семейство Ксенакантиды † Xenacanthidae Fritsch, 1889, Род Ксенакантусы — † Xenacanthus Beyrich, 1848 (известен пермского и триасового периодов, от 298,9 до 208,5 миллиона лет назад). Ископаемые различных видов найдены по всему миру. Как и все ископаемые акулы, Xenacanthus известен в основном благодаря окаменелым зубам и шипам.
Состав рода сильно различается у разных систематиков, некоторые из них синонимизирует с Xenacanthus род Pleuracanthus (см. выше), а часть видов относят к другим родам пластиножаберных рыб. Род может содержать до 20 вымерших видов:
• Xenacanthus atriossis
• Xenacanthus compressus = Pleuracanthus compressus (Newberry, 1856)
• Xenacanthus decheni = Pleuracanthus decheni (Goldfuss, 1847) — типовой вид
• Xenacanthus denticulatus
• Xenacanthus erectus
• Xenacanthus gibbosus ?= Diplodus gibbosus Agassiz, 1843
• Xenacanthus gracilis ?= Diplodus gracilis Newberry, 1856
• Xenacanthus howsei
• Xenacanthus levissimus = Pleuracanthus levissimus Agassiz, 1837
• Xenacanthus latus ?= Diplodus latus Newberry, 1856
• Xenacanthus luedernesis
• Xenacanthus moorei = Triodus moorei (Woodward, 1889)
• Xenacanthus ossiani
• Xenacanthus ovalis
• Xenacanthus parallelus
• Xenacanthus parvidens ?= Polyacrodus parvidens Woodward, 1916
• Xenacanthus ragonhai — Rio do Rasto Formation, Бразилия
• Xenacanthus robustus = Pleuracanthus robustus Davis, 1880
• Xenacanthus serratus
• Xenacanthus slaughteri
• Xenacanthus taylori
• Xenacanthus texensis
По данным сайта Paleobiology Database, на февраль 2018 года в род включают всего один вид и один опубликованный биномен без описания:
• Xenacanthus indicus Jain, 1980 [S. L. Jain. 1980. Freshwater xenacanthid (= pleuracanth) shark fossils from the Upper Triassic Maleri Formation, India. Journal of the Geological Society of India 21:39-47]
• Xenacanthus platypternus, nom. nud.
Ксенакант имел ряд особенностей, которые отличали его от современных акул. Строение тела больше напоминает строение современного угря. Эта пресноводная акула обычно была около одного метра в длину и никогда не превышала 2 м. Спинной плавник был лентовидным и проходил по всей длине спины и вокруг хвоста, где и соединялся с анальным плавником. Такое расположение плавников напоминает их расположение у современных морских угрей. И, наверное, ксенакант плавал похожим образом. Характерный шип выступал из задней части головы, и дало роду название. Высказывались даже предположения, что этот шип был ядовитым, возможно, подобно шипам скатов-хвостоколов. Это вполне вероятно, поскольку скаты — близкие родственники акул. Зубы имели необычную V-образную форму, и ксенакант, вероятно, питался мелкими ракообразными и крупночешуйчатыми палеонисцидными рыбами.
Семейство Ксеносинеходонтиды † Xenosynechodontidae Gluckman, 1980, Род Ксеносинеходусы — † Xenosynechodus Gluckman, 1980 (известен в Восточной Европы (Россия, Ишеево) из средней и поздней перми, от 268,0 до 252,3 миллиона лет назад) состоит из одного вида — Xenosynechodus egloni Gluckman, 1980.
Одним из наиболее богатых местонахождений раннетатарских позвоночных Восточной Европы является Ишеево (Татарстан), раскопки которого проводились под руководством И.А. Ефремова в конце 1930-х гг. Местонахождение приурочено к песчаной толще, образовавшейся в районе впадения речной системы в крупный солоноватоводный водоем, вследствие чего в захоронении хорошо представленными оказались как наземные, так и водные животные. При раскопках обнаружены скелеты высокотелых рыб-платисомусов (Platysomus biarmicus), уникальные черепа древних акул-ксеносинеходов, различные лабиринтодонты.
Род Анодонтакантусы — † Anodontacanthus Davis, 1881 (известен в России и США
из пермского периода, от 298,9 до 268,0 миллиона лет назад) состоит из следующих видов: Anodontacanthus americanus Hussakof, 1911, Anodontacanthus ruthenorum Chabakov 1928. [A. W. Chabakov. 1928. Anodontacanthus ruthenorum sp. n., a new ichthyodorulite from the Permian of European part of USSR. Annuaire de la Societe Paleontologique de Russia 7:127-132]
По другим данным род состоит из видов: Anodontacanthus alatus Davis, 1880; Anodontacanthus belemnoideus Zidek, 1978 и Anodontacanthus triangularis Davis, 1880. [James W. Davis On Anodontacanthus, a new Genus of Fossil Fishes from the Coal-measures; with Descriptions of three new Species, Quarterly Journal of the Geological Society, 37, 427-429, 1 February 1881]
Колючки были взяты из угольного месторождения Вест-Райдинга (West-Riding) в Йоркшире (Yorkshire), а также из месторождения Железного камня (Ironstone occurring), у Нижних известняков (Lower Limestone) около Эдинбурга (Edinburgh).
Три вида Anodontacanthus обладают специфическими характеристиками. В некоторых особенностях они напоминают род Pleuracanthus. Шипы состоят из прочного, мелкозернистого, волокнистого вещества. Их общая форма также сходна с формой шипов Pleuracanthus: внутренняя полость заканчивается у базальной конечности; стенки у основания становятся тоньше по сравнению с остальной частью колючки; метод имплантации в тело рыбы тоже должен был быть таким же. Отличительной чертой этих двух остатков является отсутствие какой-либо формы зубцов на шипе. У плевракантов есть два ряда зубцов либо вдоль каждой боковой поверхности, либо на некотором участке между боковой и задней линиями. Может ли это различие быть достаточным для создания второго рода, или дальнейшие открытия докажут, что этот род должен быть включен в род Pleuracanthus. Но в настоящее время его рассматривают как отдельный род.
На фотографии предположительно изображён шип Anodontacanthus triangularis (?) из Сэндифорда (Sandyford), Ирландия. Ширина шипа около 20 мм.
%201f.jpg)
%202f%20Eucritta%20melanolimnetes.jpg)
%203f%20fossil.jpg)
%201f%20fossil.jpg)
%202f%20skeleton.jpg)
%203f%20teeth.jpg)
%204f%20teeth.jpg)
%201f.jpg)
%202f%20fossil.jpg)
%203f%20fossil.jpg)
%204f%20fossil.jpg)
%205f%20fossil.jpg)
%206f%20fossil.jpg)
%207f%20fossil.jpg)
%208f%20fossil.jpg)










%201f%20fossil.jpg)
%201f%20fossil.jpg)
%201f%20fossil.jpg)
%201f%20fossil.jpg)
%202f%20egg%20case.jpg)
%201f%20fossil.jpg)
%201f%20fossil.jpg)

%201f%20skeleton.jpg)
%202f%20skeleton.jpg)









%201f%20teeth.jpg)
%202f%20teeth.jpg)
%201f%20fossil.jpg)
%202f%20fossil.jpg)
%203f%20fossil.jpg)




%201f.jpg)
%202f%20teeth.jpg)
%203f%20teeth.jpg)
%204f%20teeth.jpg)
%205f%20teeth.jpg)
%206f%20teeth.jpg)
%201f.jpg)
%202f.jpg)
%203f.jpg)
%204f%20skeleton.jpg)
%201f%20fossil.jpg)
%201f%20teeth.jpg)






















%201f.jpg)
%202f.jpg)
%203f.jpg)
%204f.jpg)
%205f.jpg)
%201f.jpg)