(Teleostei Müller, 1846) Инфракласс Телеостеи, Костистые рыбы, Настоящие костистые рыбы, Infraclass Teleostei Müller, 1846 (Teleosts, Teleost fishes) 42 отряда
Материал подготовил Фактистов И. Е., 2020 г. Сайт www.fishbiosystem.ru
Костистые рыбы или Костистые (Teleostei Müller, 1846) — клада Лучепёрых рыб (Actinopterygii Klein, 1885), которой присваивают ранг от инфракласса до подотдела.
Костистые рыбы впервые появляются в среднем триасе, в мелу становятся уже многочисленными, а с кайнозойской эры распространяются повсеместно, образуя необычайное многообразие форм (больше 90% ныне живущих видов рыб).
Рыбы Японского моря и сопредельных частей Охотского и Желтого морей. Часть 7. (Dactylopteriformes — Pegasiformes). Г.У.Линдберг, В.В.Федоров, З.В.Красюкова 1997 г.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ОТРЯДОВ МОРСКИХ РЫБ (OSTEICHTHYES, TELEOSTOMI), УЧТЕННЫХ В НАСТОЯЩЕЙ ЧАСТИ ОПРЕДЕЛИТЕЛЯ
(Линдберг, 1971; Lindberg, 1974.)
1 (12). Верхнечелюстные кости обычного строения, не соединены с предчелюстными костями и не слиты с ними. Зубы не слиты вместе, не образуют выступающего вперед «клюва».
2 (3). Тело и голова сильно приплюснуты сверху и покрыты костными пластинками, образующими панцирь. Грудные плавники большие, веерообразной формы, расположены горизонтально; в них от 10 до 18 неветвистых лучей, которые у основания заметно твердые, а на концах мягкие и членистые. Спинной плавник один, короткий; в нем, как и в анальном, 5 мягких неветвистых лучей. Брюшные плавники заметно позади основания грудных, в каждом из них 1 колючий и 1—3 членистых луча. В хвостовом плавнике 8 лучей. Рыло удлиненное, часто шилообразное, зазубренное. Рот нижний. Зубов нет. Плавательного пузыря нет. XXXVII. Pegasiformes
3 (2). Тело, голова и грудные плавники другой формы.
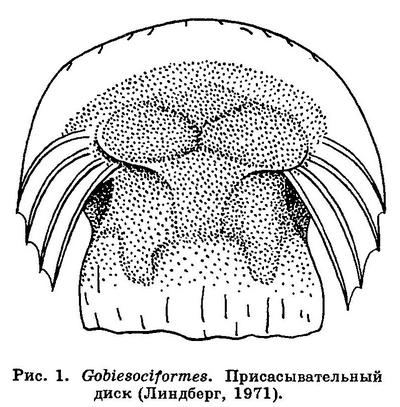
4 (5). На груди имеется присасывательный диск полуокруглой формы (рис. 1). Брюшные плавники широко отставлены друг от друга, в каждом из них один колючий (скрытый в коже) и 4 неветвистых луча. Спинной и анальный плавники расположены в задней части тела; в них нет колючих лучей. Кожа голая. Плавательного пузыря нет. XXXV. Gobiesociformes
5 (4). На груди нет присасывательного диска.
6 (7). Первый спинной плавник видоизменен в присасывательный орган, расположенный на голове и передней части тела. XXXIII. Echeneiformes
7 (6). Первый спинной плавник не изменен в присасывательный орган.
8 (9). Жаберные отверстия малы, овальной формы, расположены под или позади грудных плавников, но не впереди них, или только частично впереди них. Брюшные плавники, если имеются, расположены на горле, имеют 1 колючий и 5 ветвистых лучей. Первый спинной плавник представлен отдельными лучами, не соединенными перепонкой, первый из них удлинен и превращен в «удочку» (illucium). Голова и передняя часть тела приплюснуты. XXXVI. Lophiiformes
9 (8). Жаберные отверстия расположены впереди грудных плавников.
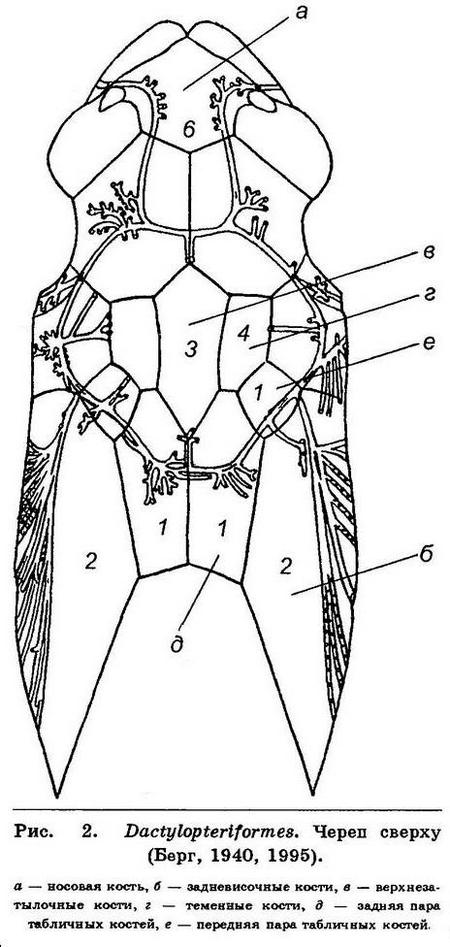
10 (11). Голова как бы заключена в костный шлем, так как носовые кости слиты в одну, задневисочные кости (posttemporale) очень большие, удлинены в каудальном направлении, а позади верхнезатылочной и теменных костей имеются 2 пары табличных косточек (tabularia), из которых задняя пара очень крупная. Грудные плавники очень большие, часто
достигают основания хвостового плавника (рис. 2). XXXII. Dactylopteriformes
11 (10). Тело, голова и плавники иного строения, не такие как на рис. 2. (Рыбы, учтенные в предыдущих шести частях определителя.)
12 (1). Верхнечелюстные кости плотно соединены с предчелюстными костями, иногда даже слиты с ними. Зубы иногда хорошо различимы, но нередко слиты вместе и образуют «клюв». Рот очень мал. Жаберные отверстия маленькие. Тело иногда покрыто костными пластинками, образующими панцирь. XXXIV. Tetraodontiformes
Атлас-определитель рыб: Книга для учащихся. Н.А.Мягков 1994 г.
Костистые рыбы (Teleostei)
Систематическая группа, объединяющая прогрессивные надотряды современных костных рыб (более 98% видов), наиболее обычных в пресных и морских водах планеты, в том числе и в водах нашей страны. Объединяет представителей восьми надотрядов рыб, из которых представители семи — обычны в наших водах.
Надотряд Клюпеоидные (Clupeomorpha)
Один спинной плавник без колючих лучей, последних нет и в других плавниках. Жировой плавник есть или его нет. Чешуя циклоидная. Сравнительно примитивные древние пресноводные морские и проходные рыбы. В наших водах обитают представители двух отрядов; наиболее обычны лососи, сиги и сельди, являющиеся ценнейшими объектами промысла.
Надотряд Араваноидные (Osteoglossomorpha)
Древняя группа рыб, примыкающая к клюпеоидным и ранее включавшаяся в предыдущий надотряд. Характерно положение брюшных плавников (лежат посредине брюшка) и внутреннее строение ряда важных органов. Объединяет два отряда тропических пресноводных рыб, представители которых отсутствуют в наших водах.
Надотряд Ангвиллоидные (Anguillomorpha)
Представители отличаются длинным змеевидным телом; обычно лишены брюшных, а некоторые и грудных плавников (редуцированы). Характерно развитие с матаморфозом, личинка (лептоцефал) имеет листовидную форму и сильно отличается от взрослых рыб (ранее их даже описывали в качестве самостоятельных таксонов). В наших водах обычны представители одного отряда.
Надотряд Циприноидные (Cyprinomorpha)
Многочисленный надотряд (включает более 6 тыс. видов) преимущественно пресноводных рыб. Брюшные плавники лежат далеко за грудными. Один спинной плавник; нет настоящих колючих лучей в плавниках, лишь у некоторых видов первые членики ветвистых лучей спинного и грудных (реже и анального) плавников срастаются в мощную колючку (первую в плавнике). В наших водах обитают многочисленные представители двух отрядов.
Надотряд Атериноидные (Atherinomorpha)
У представителей немногочисленного отряда тело в сечении округлое, рот большой. Один или два спинных плавника без мощных колючек, жировой плавник отсутствует. Морские и пресноводные рыбы теплых и умеренных широт. В наших водах обитают представители трех отрядов.
Надотряд Параперкоидные (Parapercomorpha)
Объединяет рыб, находящихся как бы между довольно примитивным и прогрессивным отрядами, между карпообразными и окунеобразными. Есть общие примитивные черты и признаки специализации (например, в строении и положении плавников). Преимущественно морские рыбы. В наших водах весьма многочисленны представители одного отряда, в большинстве своем ценные объекты морского рыболовства.
Надотряд Перкоидные (Percomorpha)
Характеризуется наличием колючих нечленистых лучей в плавниках и положением брюшных плавников, лежащих на груди или на «горле» рыбы. Чешуя ктеноидная. Объединяет большое число прогрессивных в эволюционном, молодых в историческом и разнообразных в экологическом планах рыб. В наших водах обитают представители шести отрядов.
Надотряд Батрахоидные (Batrachomorpha)
Для представителей этого сравнительно малочисленного надотряда морских рыб характерны широко расставленные брюшные плавники с малым числом лучей. В наших водах обитают представители одного отряда.
Под общим названием костистые или высшие (настоящие) рыбы объединяют представителей 32 отрядов рыб, входящих в восемь надотрядов. В наших водах эпизодически встречаются представители более чем 20 из них, но постоянно обитают рыбы 16 отрядов данной сборной группы.
Аквариумные тропические рыбы. И.Петровицкий 1984 г.
СХЕМАТИЧНОЕ ИЗОБРАЖЕНИЕ СПОСОБОВ НЕРЕСТА РЫБ, ПОКАЗАННЫХ В РАЗДЕЛЕ С ИЛЛЮСТРАЦИЯМИ
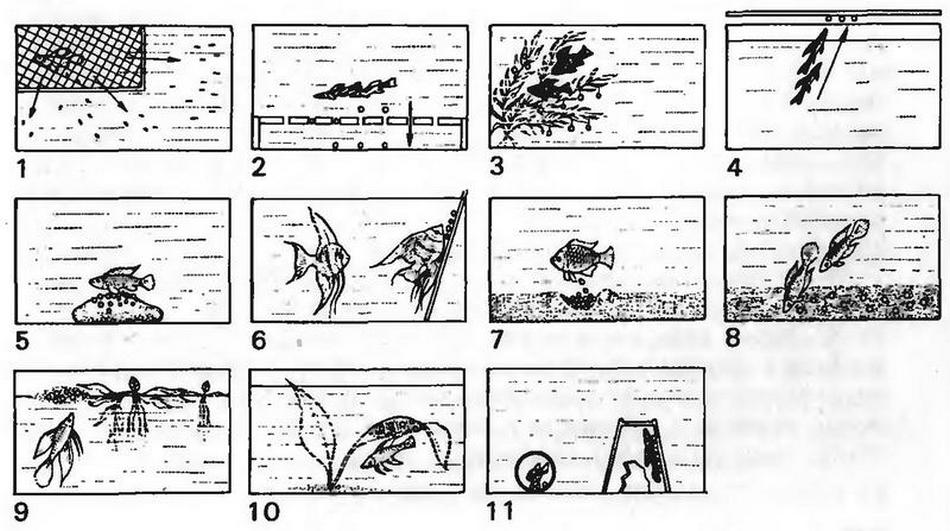
1. Нерестовые корзинки рекомендуются для живородящих рыб. Корзинки помещаются в средние и большие по емкости вырастные аквариумы. Они также служат убежищем для мальков при нападении самки.
2. Если икринки рыб нелипкие, то на дно аквариума помещают сепараторную сетку, размер ячеек которой обеспечивает беспрепятственное прохождение опускающихся на дно икринок. Сетка защищает икринки от каннибализма родителей.
3. Липкие икринки или икринки с волокнами откладываются в сплетении растений. Оно же частично предохраняет икринки от каннибализма родителей.
4. Нерест вне воды является исключительным случаем, но аквариумистам он хорошо известен благодаря виду Copella arnoldi. В неволе икринки откладываются на верхнее стекло, в природе — на листья прибрежных растений, склоняющихся к поверхности воды. При нересте рыбы совершают совместные прыжки к субстрату. Время от времени самцы появляются около нерестилища и хвостовым плавником брызгают воду на икринки. После выклева личинки падают в воду, и тогда заботы самца кончаются.
5. Икринки могут быть отложены на твердый субстрат (камни, ветви). В большинстве случаев это свойственно видам с высокоразвитым инстинктом заботы об икринках и молоди, причем проявляется он у обоих родителей или у одного из них. Некоторые виды вынашивают икринки во рту («инкубирующие»), где некоторое время продолжается развитие мальков. Если виды или производители имеют склонности к каннибализму, то субстрат с икринками следует перенести в отдельный аквариум или выловить производителей.
6. Как и в случае, описанном в пункте 5, рыбы мечут икру на твердый субстрат, но расположенный вертикально или наклонно — боковые стекла, ветви, большие листья и стебли растений (род Pterophyllum). И здесь икринки можно изолировать, перенеся их в отдельный сосуд вместе с субстратом или сначала смести их мягкой щеточкой в марлю.
7. Икринки могут откладываться в подготовленные в грунте ямки. У некоторых видов один из родителей или оба охраняют нерестилище в период развития икринок.
8. Некоторые виды рыб, особенно те, которые распространены в периодических водах, откладывают икринки в мягкий субстрат дна, который предохраняет их от высыхания в неблагоприятных условиях засухи, когда взрослые рыбы гибнут. После того, как водоемы снова наполнятся водой, развитие икринок завершается и из них выклевываются личинки. Личинки быстро превращаются в мальков, те быстро растут и вскоре становятся половозрелыми.
9. Икринки откладываются в пенные гнезда на поверхности воды, где гнезда крепятся в сплетении водных и болотных растений (лабиринтовые рыбы), или рыбы строят их под широкими листьями водяных лилий или подобных растений (сомики). В аквариумах широкие листья имитируются подвешенными, перевернутыми вверх дном пластмассовыми мисками. В обоих случаях гнездо строит самец и он же заботится об икринках, а позднее — о личинках до той поры, пока не исчезнет желточный мешок и мальки станут самостоятельными.
10. Как и в предыдущем случае, икринки откладываются в пенные гнезда. Рыбы их строят под поверхностью или в средних слоях воды, укрепляя их под листьями растений или на козырьках скалистых террас.
11. Некоторые виды рыб находят себе нерестилища в полостях предметов, в щелях и пещерках. Такие условия можно создать с помощью цветочных горшков или трубок подходящего диаметра. И в этом случае можно легко манипулировать с субстратом и икринками. Охраняют нерестилище оба партнера или один из них, чаще всего — самец.
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
Около 200 миллионов лет назад
появились первые костистые рыбы (Teleostei).
Вначале немногочисленные, они бурно
развились в течение мелового периода
(за 135—70 миллионов лет до нашего
времени), возобладав над ганоидными
рыбами, достигнув необыкновенного
разнообразия и фактически завоевав воды нашей
планеты. К настоящему времени
сохранилось всего около 45 видов ганоидных рыб,
тогда как костистых рыб насчитывается
около 20 000 видов, что примерно в 30 раз
больше, чем всех остальных рыб и
рыбообразных, вместе взятых.
НАСТОЯЩИЕ КОСТИСТЫЕ РЫБЫ (TELEOSTEI)
Под общим названием костистых рыб,
или настоящих костистых рыб (Teleostei), обычно противопоставляют более
примитивным ганоидным рыбам все
остальные группы (надотряды) и отряды
костных рыб. В их строении, как
правило, не имеется такого набора архаических
признаков, как у ганоидных, хотя
отдельные признаки обнаруживаются у
некоторых видов менее специализованных
семейств и отрядов.
У костистых рыб хвостовой плавник
внешне равнолопастный — гомоцеркальный, и только скелет его основания
асимметричен, отражая происхождение от
асимметричного гетероцеркального
плавника. Осевой скелет хорошо
окостеневающий, хотя у некоторых видов может
сохраняться между телами позвонков хорда.
Как правило, нет артериального конуса
в сердце, а в кишечнике нет спиральной
складки, сохраняющихся только у
единичных видов. Чешуя, в виде тонких костных
пластинок, гладких (циклоидная чешуя)
или с шипиками (ктеноидная чешуя), не
ромбической формы. Количество лучей в
спинном и анальном плавниках
соответствует числу поддерживающих их скелетных
элементов (радиальных лучей) в отличие от
того, что имеется у осетрообразных и многоперообразных ганоидных рыб (а также
у хрящевых рыб). Выводное отверстие
отодвинуто, как правило, от оснований
брюшных плавников.
Различаются три основные
группировки отрядов костистых рыб. Менее специализованная группировка, содержащая
отряды, появляющиеся уже начиная с
нижнего мела (т. е. свыше 100 миллионов лет
до нашей эры) и сохранившие некоторые
древние черты строения, охватывает
надотряды клюпеоидных, араваноидных, ангвиллоидных, циприноидных и отчасти
атериноидных рыб, преимущественно
сельдеобразного, лососеобразного,
щукообразного, карпообразного или угреобразного
облика.
Для большинства рыб этих групп
характерны положение брюшных плавников
в средней трети тела на брюхе
(«абдоминальное» положение), наличие только
одного спинного плавника или одного
обычного спинного плавника и
добавочного жирового плавника (не содержащего
скелетных лучей); отсутствие нечленистых
колючих лучей в плавниках (хотя у
некоторых карповых и сомовых имеются
толстые, нередко зазубренные костные
колючки, а у угревидных спиношипообразных рыб (Notacanthiformes) есть и колючие
лучи на спине); обычно наличие
открывающегося протоком в пищевод
плавательного пузыря и другие признаки.
В меловой период, т. е. 70—100
миллионов лет назад, наряду с этими группами
рыб появилась вторая группировка —
параперкоидные рыбы — переходная к позже развившимся прогрессивным
колючеперым рыбам.
Третью, наиболее прогрессивную,
группировку представляют настоящие
колючеперые рыбы, появившиеся с
верхнемелового периода и достигшие наибольшего
расцвета в нашу эпоху. Это
многочисленные отряды перкоидных и батрахоидных
рыб окунеобразного, скумбриеобразного,
камбалообразного и других типов
строения. Скелет плавников у них обычно
состоит из колючих и членистых лучей,
брюшные плавники сдвинуты вперед и
находятся на груди или на горле
(«торакальны» или «югулярны»),
плавательный пузырь не имеет соединения с
пищеводом.
НАДОТРЯД КЛЮПЕОИДНЫЕ (CLUPEOMORPHA)
Объединяемые в этом надотряде
отряды тарпонообразных, гоноринхообразных,
сельдеобразных, лососеобразных, миктофообразных и китовидкообразных рыб
содержат наименее специализованных
костистых рыб, частично сохранивших
некоторые архаические черты строения.
Плавники у рыб этой группы не имеют
нечленистых колючих лучей, почему эту
группу называли ранее мягкоперыми
(Malacopterygii). Брюшные плавники
находятся, как правило, в средней части
брюха («абдоминальны») и содержат
обычно не менее 6 лучей. Основания грудных
плавников расположены низко, вдоль
брюшного края тела или немного
наклонно к нему. Чешуя без шипиков,
циклоидная (за единичными исключениями).
Верхняя челюсть образована обычно
несколькими костями (предчелюстные,
верхнечелюстные). Позвонки однотипны, не
изменены добавочными скелетными
образованиями, поэтому рыб этой группы
называли также равнопозвонковыми (Isospondyli). Сюда относятся рыбы, имеющие
сельдеобразное или вальковатое тело, с
одйим спинным плавником, основание
которого обычно сдвинуто вперед
относительно основания анального плавника.
Остатки этой группы встречаются
начиная со среднего триаса, т. е. примерно
за 150—160 миллионов лет до нашей
эры.
В этой группе объединяются отряды
трех типов строения. Наиболее близки
к исходным предкам рыбы сельдевидного
облика, имеющие сельдеобразное (иногда
вальковатое) тело с серебристыми
боками, у многих без боковой линии, не
имеющие жирового плавника позади
сдвинутого кпереди спинного плавника.
Плавательный пузырь у этих рыб обычно имеет
выросты, подходящие впереди к слуховым
капсулам. Таковы Тарпонообразные
(Elopiformes), Гоноринхообразные (Gonorhynchiformes) и собственно
Сельдеобразные (Clupeiformes).
Ко второму типу строения относятся
рыбы, имеющие обычно позади спинного
плавника жировой плавник, нередко
сохраняющие остатки хорды в осевом
скелете и хрящ в черепной коробке, не
имеющие, в отличие от сельдеобразных,
нижних межмышечных косточек и имеющие
обычно боковую линию. Таковы лососевидные рыбы из отрядов Лососеобразные
(Salmoniformes) и Миктпофообразные
рыбы (Myctophiformes). Во многих
семействах этого типа строения встречаются
глубоководные рыбы, имеющие органы
свечения.
К третьему типу строения относятся щуковидные, галаксиевидные, стомиевидные,
гладкоголововидные рыбы из
лососеобразных и Китовидкообразные (Cetomimiformes). Спинной плавник у них
отодвинут назад и располагается над анальным,
тело у многих удлиненное. Жирового
плавника у большинства нет. Многие
семейства глубоководные.
Остатки лососеобразных рыб
встречаются в отложениях начиная с
верхнемелового периода, в основном же — в
третичном периоде. Таким образом, рыбы
этого типа строения появляются
несколько позже тарпонообразных и
сельдеобразных.
НАДОТРЯД АРАВАНОИДНЫЕ OSTEOGLOSSOMORPHA)
Надотряд включает два отряда
пресноводных рыб — отряд араванообразных и
отряд клюворылообразных. Характерны
для них абдоминальное (в средней части
брюха) положение брюшных плавников
(кроме одного лишь Pantodon),
супротивное расположение спинного и анального
плавников (кроме имеющих угревидное
тело гимнарховых рыб и длиннохвостых,
сжатых с боков нотоптеровых рыб), низко
посаженные грудные плавники,
архаические черты в строении черепа. Это
древняя группа, непосредственно
примыкающая к клюпеоидным и включавшаяся
ранее в их состав.
НАДОТРЯД АНГВИЛЛОИДНЫЕ (ANGUILLOMORPHA)
К данному надотряду относятся рыбы,
имеющие очень удлиненное тело
характерной угревидной или змеевидной формы,
у большинства с длинными спинным и
анальным плавниками, но не имеющие
обычно брюшных, а некоторые и грудных
плавников. Плечевой пояс не связан с
черепом, как у других групп рыб, а
прикрепляется к позвоночнику. Очень
характерно прохождение во время развития
особой личиночной стадии — лептоцефала. Личинка-лептоцефал имеет
маленькую голову и сжатое с боков высокое
прозрачное тело, напоминающее по форме
приостренный на обоих концах лист ивы.
К ангвиллоидным рыбам относятся угреобразные, мешкоротообразные и спиношипообразные.
НАДОТРЯД ЦИПРИНОИДНЫЕ (CYPRINOMORPHA)
Этот надотряд содержит около 6000
видов, уступая по объему только надотряду
перкоидных (Percomorpha). Циприноидные рыбы доминируют в пресных водах,
составляя не менее половины всех видов
пресноводных рыб. Почти не встречаясь
в морях, они заселяют внутренние воды
всех материков, за исключением только
Австралии и Антарктики.
Циприноидные рыбы имеют много
общего по строению с клюпеоидными,
существенно отличаясь от последних,
однако, устройством передних 4—5
позвонков, части которых видоизменены в серию
косточек, соединяющих плавательный
пузырь с внутренним ухом. Это так
называемый Веберов аппарат, представляющий
собой, по-видимому, специальное
устройство для усиления слуха. Отсюда и
старое название циприноидных рыб — костнопузырные (Ostariophysi). Характерны
для циприноидных также положение
брюшных плавников около середины
брюха («абдоминальные» плавники), один
спинной плавник. У них нет настоящих
колючих лучей в плавниках; имеющиеся
у некоторых карповых и сомовых рыб
толстые зубчатые шипы у переднего края
спинного, анального и грудных плавников
образованы путем слияния члеников
мягких лучей. Плавательный пузырь
соединяется, как правило, протоком с
кишечником. Два больших отряда относятся к
циприноидным рыбам — карпообразные
и сомообразные.
НАДОТРЯД АТЕРИНОИДНЫЕ (ATHERINOMORPHA)
Надотряд объединяет три отряда — карпозубообразных, атеринообразных и сарганообразных рыб, распространенных в
тропических, субтропических и умеренно
теплых водах. Большинство видов имеет
вальковатое тело (у некоторых
удлиненное) с одним или двумя спинными
плавниками. Спинной и анальный плавники
большей частью отодвинуты в заднюю
половину тела; основания грудных
плавников сдвинуты на бока, вверх от
брюшного края тела; брюшные плавники
расположены в средней или передней части
брюха или еще далее кпереди — на
груди, содержат 6—7 (редко 3—4)
плавниковых лучей.
В некоторых группах самцы и самки
различаются по внешнему виду, резко
выражен половой диморфизм и имеются
замечательные приспособления для
внутреннего оплодотворения.
Большинство видов откладывает икру,
некоторые группы живородящие.
Икринки крупные, сферические, почти у всех
донные, с плотной оболочкой, снабженные
прилипающими нитевидными или волосковидными выростами; у эмбрионов
сердце сдвинуто вперед от головы, на
переднюю часть желточного мешка.
Всего насчитывается 650—700 видов
атериноидных рыб. Они широко
распространены в морских, солоноватых и
пресных водах тропического и умеренных
поясов, причем большинство солоноватоводных и пресноводных видов обитает в
водах Юго-Восточной Азии, Индо-Малайского архипелага, Австралии. Именно
среди них имеются более примитивные и
близкие друг к другу виды разных групп,
что указывает на вероятное
происхождение и формирование здесь основных
направлений эволюции атериноидных рыб.
НАДОТРЯД ПАРАПЕРКОИДНЫЕ (PARAPERCOMORPHA)
Надотряд объединяет отряды рыб,
переходные по строению от более
примитивных к достигшим наивысшей для костных
рыб специализации колючеперым, перкоидным рыбам. Представители этих
отрядов имеют частично признаки,
характерные для более примитивных форм;
таковы, например, жировой плавник у
перкопсовых, выдвинутое вперед
положение обонятельных долей около носовых
капсул у перкопсовых, тресковых и ошибневидных, отсутствие колючек в
плавниках у трескообразных и ошибневидных,
наличие у многих циклоидной чешуи и
другие признаки. В то же время имеются
признаки специализации: брюшные
плавники обычно сдвинуты кпереди и
расположены под грудными или впереди них,
плавательный пузырь не соединен с
кишечником, нет костных клеток в костях, нет
межмышечных косточек и т. д.
В надотряде параперкоидных рыб
объединяются отряды перкопсообразных и
трескообразных рыб.
НАДОТРЯД ПЕРКОИДНЫЕ РЫБЫ (PERCOMORPHA)
Почти все перкоидные рыбы
характеризуются наличием колючих нечленистых
лучей в плавниках; положением
брюшных плавников на груди или на горле
(только у рыб отрядов кефалеобразных
и колюшкообразных они бывают
расположены позади грудной области и в
средней части брюха, как у более
примитивных отрядов); ктеноидной, с зубчиками,
чешуей (но у некоторых чешуя бывает
вторично циклоидная, или заменяется
костными бляшками, пластинками, или ее
нет вовсе); рот окаймлен сверху только
предчелюстными костями; плавательный
пузырь замкнутый, не соединен с
кишечником или его нет; на костях головы у
многих шипы.
Надотряд перкоидных рыб превосходит
по объему другие надотряды. Он включает
10 отрядов — Бериксообразные, Солнечникообразные, Опахообразные,
Колюшкообразные, Кефалеобразные, Слитножаберникообразные,
Окунеобразные, Скорпенообразные, Камбалообразные и Иглобрюхообразные. Всего он содержит
190—200 семейств, около 1500 родов,
около 9000 видов.
НАДОТРЯД БАТРАХОИДНЫЕ (BATRACHOMORPHA)
Надотряд батрахоидных охватывает
рыб, имеющих короткое и широкое
тело, с широколобой большой головой.
Очень характерны для них широко
раздвинутые брюшные плавники,
содержащие малое число лучей. Спинной и
анальный плавники супротивные (один над
другим), хвостовой плавник округлый
или усеченный, с малым числом лучей.
К батрахоидным рыбам относятся
отряды пегасообразных (относимый
нередко к перкоидным), батрахообразных,
присоскообразных и удильщикообразных.
Обитатели бездны. Уильям Кроми 1971 г.
НАСТОЯЩИЕ РЫБЫ
(В русской ихтиологической литературе акулы, как и другие первичноводные (то есть никогда не бывшие наземными) челюстноротые, называются рыбами, а бесчелюстные, миноги и миксины, — рыбообразными. Американский ихтиолог Г. Нельсон надкласс челюстноротых делит на два класса, один из которых (Elasmobranchiomorpha) включает в себя акул, скатов и химер, а второй (Teleostomi) — всех остальных рыб и всех наземных позвоночных. Такое деление, в сущности, не противоречит взглядам советского ихтиолога Г. В. Никольского на распределение рыб в геологической шкале времени и на их родственные связи.)
Около 450 миллионов лет назад крохотное, похожее на головастика существо взмахом своего мускулистого хвоста освободило позвоночных животных от необходимости всю жизнь ползать и пресмыкаться по дну. Постепенно к хвосту этого первого плавающего существа прибавлялись плотные клетки, наполненные водой, и в конце концов сформировалась желеобразная хорда. По мере того как хорда становилась прочнее, головастик стал плавать более энергично, хотя и несколько хаотично. Он вырос до нескольких дюймов и в какой-то момент оказался заключенным в костяную броню. Прочный череп и кольчуга из плакоидных пластинок, или чешуек, были необходимы для защиты от таких грозных хищников, как похожие на скорпионов эвриптериды длиной около 2,5 метра. Эти чудовищные «жуки» имели веслообразные ноги и могли, по-видимому, передвигаться с такой же скоростью, как и ранние рыбы, которые плавали, неуклюже изгибаясь всем телом из стороны в сторону, наподобие нынешних миног и миксин.
Со временем скорость плавающего существа увеличилась, и у одной группы животных такого рода появились челюсти, что позволяло им обороняться от хищников и хватать добычу. Обладая таким огромным преимуществом, эти плакодермы, или панцирные рыбы, сделались агрессивными и стали властелинами мира воды. Появились разновидности рыб разной величины, начиная от акулообразных рыб размером с миногу и кончая девятиметровыми дредноутами с огромной, покрытой панцирем головой. Вымершие около 250 миллионов лет назад, плакодермы явились родоначальниками как хрящевых акул, так и костистых рыб.
Такова довольно приблизительная картина происхождения рыб, основанная на исследовании ископаемых фрагментов костей древних животных и на изучении строения современных существ. Прямых свидетельств не существует, толковать данные можно по-разному. Возможно, что акулы возникли от животных, для которых море было родным домом, а костистые рыбы — от пресноводных предков. Некоторые данные говорят о том, что рыбы, по крайней мере многие из них, появились в реках, а затем мигрировали в моря.
Исследуя ископаемые останки животных, мы видим, что в мире рыб, когда их положение упрочилось, произошел «демографический взрыв». Они начали развиваться во всех направлениях, стало увеличиваться их число и количество видов. Ныне они столь же многочисленны, как звезды на небе, но разнообразие их еще более поразительно. Разновидностей рыб больше, чем всех остальных позвоночных вместе взятых. Рыб — тех, что вы кладете на сковородку, заказываете в ресторане или с которыми вы меряетесь смышленостью во время воскресной рыбалки, — существует около 20 000 различных форм (Их во много раз больше, если в понятие «форма» автор включает и внутривидовые разности. Число видов рыб очень неопределенно: непрерывно описываются новые виды, закрываются и вновь восстанавливаются старые. Надежное выделение вида требует глубоких и разносторонних знаний, а многие группы рыб и во многих районах изучены еще очень слабо.). Под ломтиком лимона, маслом и петрушкой лежит поистине благородное животное, властелин морей, венец своего направления эволюционного развития.
Существуют торпедообразные, трубообразные, круглые, плоские рыбы, а также рыбы, форма которых не поддается описанию. Самые малые — некоторые бычки — имеют в длину всего 13 миллиметров, самые крупные — китовые акулы — достигают 14 метров. Есть рыбы, дышащие воздухом, рыбы, которые летают, ходят, разговаривают и даже лазают по деревьям. Древнейшие из ныне существующих позвоночных, за 450 миллионов лет своего развития они приспособились и проникли в каждый уголок бескрайнего мира воды: холодные, мрачные глубины, сумеречное мелководье, залитые солнцем воды открытого моря и изрезанные гротами и пещерами коралловые рифы, быстрые потоки и пруды с застоявшейся водой. В сущности, эволюция рыб — это история приспособления организмов к новой среде и иному образу жизни.
Некоторые костистые рыбы, как, например, тунец, превратились в самых быстрых, наиболее развитых обитателей моря; такие рыбы, как осетр, остались во многом похожими на своих предков.
ЧЕШУЙЧАТЫЙ ПАНЦИРЬ
У древних осетров скелет состоял из прочных костей, у современных же видов они заменены хрящом. Замена почти завершена: большая часть оставшихся костей находится вне, а не внутри длинного, стройного тела рыбы. Пять рядов костных жучек, далеко поставленных друг от друга рядов плакоидных дисков, проходят от головы до хвоста. Острые пластинки на верхней части «акульего» хвоста осетра используются как оружие и могут больно порезать всякого, кто зазевается. Эти пластинки — напоминание о поре, когда все рыбы были облачены в кольчугу, состоявшую из костяных пластинок, тесно поставленных и эластично соединенных меж собой.
По мере того как для рыб все большее значение приобретали скорость и маневренность, а не прочность брони, на смену мозаике тяжелых пластинок пришла более тонкая, легкая чешуя. Уже около 50 миллионов лет назад у большинства рыб появились мягкие, гибкие чешуйки, перекрывающие друг друга подобно черепице. Укрепленные во внутренних слоях кожи, они, при всей их кажущейся непрочности, представляют достаточно надежную защиту.
У огромного большинства рыб чешуйки покрыты тонким, почти невидимым слоем кожи. У некоторых же видов, как, например, у меч-рыбы и многих сомовых, чешуя отсутствует; у угрей внешний слой кожи настолько толст, что видны лишь самые верхние кончики чешуек. В отличие от громоздкой кольчуги, обременявшей древних предков рыб и приковывавшей их ко дну, современная чешуя позволяет делать быстрые, ловкие движения и является приспособлением к очень подвижной жизни в верхних слоях воды.
Увеличению скорости перемещения в воде способствует наличие на чешуе слоя слизи, уменьшающей трение между телом рыбы и водой (Вероятно, в этом участвует и чешуя.). Выделяемая невидимыми железами, разбросанными по всей поверхности тела, эта слизь служит также антисептическим средством, препятствующим появлению бактерий, грибков и других организмов, замедляющих скорость рыбы и опасных для ее здоровья. Слизь вместе с чешуей представляет собой прочное покрытие, сохраняющее жизненные соки рыбы и мешающее проникновению внутрь нее воды (Иной раз слизь защищает и от хищников. Некоторые рыбы-попугаи (Scaridae) надевают на ночь «рубашку» из толстого слоя слизи, предохраняющую от нападения мурен. Мощный слой слизи защищает мальков Mupus maculatus от поражения щупальцами физалии, среди которых они прячутся. По запаху слизи стайные рыбы, а также самцы и самки не стайных рыб нередка находят друг друга. Рыбки Symphysodon discus и Etroplus maculatus выкармливают мальков собственной слизью. Родственники нашего угря, закапывающиеся угри, всю свою жизнь проводит в норках, которые роют в песке. Их слизь очень быстро цементирует стенки такой норки. Вот какой разнообразной может быть функция слизи у рыб. Такого же рода примечания можно было бы сделать при обсуждении функций любого их органа.).
Чешуйки растут вместе с рыбой. Новый «материал» образуется в виде колец по краям чешуек (Говоря о кольцах, автор имеет в виду микроскопические валики (склериты), которые образуются на верхнем слое чешуи; снизу чешуя растет по всей площади, и склериты там не формируются. В течение года может появиться всего лишь несколько склеритов или несколько сотен их — все зависит от строения чешуи у данного вида рыб и от скорости роста рыбы в данном году.). Эти кольца становятся более многочисленными, а расстояние между ними — более значительным, когда пища в изобилии и размеры рыбы быстро увеличиваются. С уменьшением количества пищи в холодное время года рост замедляется, а то и вовсе прекращается и кольца становятся все уже и чаще (Близко расположенные один от другого склериты, часто к тому же неполные, идущие лишь по переднему краю чешуи, образуют годовое кольцо. Но у некоторых рыб, например у сельдей, годовые кольца образуют не склериты, а темные и светлые зоны на внутреннем слое чешуи, подобные кольцам на костях.). За этой сменой времен года можно проследить по светлым (летним) и темным (зимним) полосам на отолитах (Отолиты, или «ушные камни», находятся у рыб в органе равновесия — лабиринте, составляя его часть.) и некоторых костях. Как у деревьев, у рыб имеются годовые кольца, и зачастую, подсчитав их количество, можно узнать возраст рыбы (Кроме годовых колец, на отолитах, костях, срезах лучей плавников, а особенно часто на чешуе по разным причинам нередко образуются дополнительные кольца, которые затрудняют определение возраста, но если в них разобраться, дают дополнительную информацию о важных событиях в жизни рыбы — нересте, массовом заражении паразитами и т. д.). А поскольку размер чешуйки пропорционален размеру рыбы, иногда можно определить, какой величины рыба достигала к концу каждого года.
В 1953 году канадские биологи, сосчитав концентрические кольца на срезе с колючки грудного плавника осетра весом 97 килограммов, установили, что рыба достигла преклонного возраста — 152 года. К прочим долгоживущим рыбам относятся 227-килограммовый палтус, проживший до 60 лет, и угорь, достигший 55-летнего возраста. Крупные рыбы и черепахи, вероятно, живут долее других животных, старея настолько медленно, что в большинстве своем гибнут от «несчастных случаев» и болезни, а не от дряхлости и упадка сил.
Фауна СССР. Рыбы. Том IX, вып.4. Трескообразные. А.Н.Световидов 1948 г.
КРАТКИЙ ИСТОРИЧЕСКИЙ ОЧЕРК СИСТЕМЫ ОТРЯДА
В отряд Anacanthini J. Müller (1846 : 177), впервые установивший его, объединил рыб, имеющих плавники без колючих лучей, брюшные плавники, если они имеются, на горле или груди, плавательный пузырь, если он имеется, не соединенный с кишечником. Всю обширную группу рыб, объединяемую в один отряд по этим признакам, J. Müller делит следующим образом (l. с. : 202):
Ordo Anacanthini.
Subordo I. Anacanthini subbrachii.
Fam. 1. Gadoidei.
2. Pleuronectides.
Subordo II. Anacanthini apodes.
Fam. 1. Ophidini.
В таком объеме отряд Anacanthini, как указывает сам J. Müller (l. с. : 177, 199), состоит частично (Anacanthini subbrachii) из Malacopterygii subbrachii, частично (Anacanthini apodes) из Malacopterygii apodes системы Cuvier (1817 : 212, 229). По внутреннему строению J. Müller считает Anacanthini близкими к отряду Anacanthopterygii, с которыми они, точно так же как и с Pleuronectidae, сближались долгое время и впоследствии.
Günther (1862 : 317), принимая отряд Anacanthini в объеме, установленном J. Müller'ом, считает, что колючие лучи у них имеются только в вертикальных плавниках, и делит отряд на два подотряда несколько иначе:
Anacanthini Gadoidei. Голова симметричная. Семейства: Gadopsidae, Lycodidae, Gadidae, Ophidiidae (группы: Brotulina, Ophidiina, Fierasferina, Ammodytina, Congrogadina), Macruridae, Ateleopodidae.
Anacanthini Pleuronectoidei. Обе стороны головы несимметричны. Семейство: Pleuronectidae.
Система Günther'a отличается и большим подразделением внутри подотрядов.
Соре (1872 : 341) определяет Anacanthini как рыб, у которых основание черепа простое (без миодома), posttemporale вильчатое, лопаточное отверстие между scapula и coracoideum; pharyngialia три, горизонтальные, третья более длинная; лучи спинных плавников мягкие; хвостовой плавник дифицеркальный. По этим признакам он относит к Anacanthini, считаемым им за одну из групп отряда Percomorphi, только семейства Gadidae и Macruridae, выделяя большинство сближаемых с ними семейств в особую группу того же отряда Scyphobranchii. Камбал Соре выделяет в самостоятельный отряд Heterosomata, в качестве которого они приняты почти во всех более поздних системах. Система Anacanthini Соре является значительным шагом вперед, так как объем и многие положенные им в основу ее признаки приняты в большинстве более поздних систем.
Jordan и Gilbert (1883 : 783) в подстрочном примечании указывают, что в отряде Acanthopteri семейства Gadidae, Macruridae, Lycodidae, Congrogadidae, Fierasferidae и Ophidiidae образуют особую группу или подотряд Anacanthini или Jugulares, Отличаемый от других Acanthopteri длинными спинными и анальными плавниками, состоящими только из мягких лучей. Семейство Gadidae они разделяют на подсемейства Gadinae, Merlucciinae, Chiasmodontinae, Brotulinae, хотя последнее в определительных таблицах (р. 79) выделено ими в самостоятельное семейство.
Gill (1884 : 167) дает следующие подразделения Anacanthini или Jugulares:
Gadоidеа. Орбито-ростральная часть черепа длиннее, чем задняя, черепная полость широко открыта спереди, supraoccipitale хорошо развито и с килем сзади, лопаточное отверстие между scapula и coracoideum. Семейства: Gadidae, Merlucciidae, Bregmacerotidae, Ranicipitidae, Macruridae.
Ophidioidea. Орбито-ростральная часть черепа сжата и короче, чем задняя, черепная полость частично замкнута расширением и соединением parasphenoideum и frontalia, scapula с отверстием в середине. Семейства: Brotulidae, Brotulophididae, Ophidiidae, Fierasferidae, Congrogadidae.
Lycodoidea. Орбито-ростральная часть черепа сжата и короче, чем задняя, черепная полость спереди открыта, но ограничена с боков расширением соприкасающихся parasphenoideum и frontalia, supraoccipitale наклонно и крышевидно сзади, scapula с отверстием в середине. Семейства: Lycodidae, Ammodytidae, Ateleopodidae, Xenocephalidae,
Все группы семейства Ophidiidae и ряд родов семейств Gadidae, Ophidiidae и Ateleopodidae системы Günther'a Gill считает семействами; семейство Gadopsidae и род Chiasmodus он считает семействами Acanthopterygii. Весьма существенным в системе Gill'а является то, что в основе разделения отряда на группы им указаны новые остеологические признаки, имеющие значение и в настоящее время.
Значительно ближе к современному объему, взяв в основу признаки группы Gadoidea системы Gill'a, принимают Anacanthini Jordan и Evermann (1898 : 2528), считая их точно так же одним из подотрядов Acanthopterygii. К Anacanthini они относят семейства Gadidae, Merlucciidae, Macruridae. Семейство Ranicipitidae не включено в состав подотряда Anacanthini, вероятно, только потому, что его у берегов Сев. Америки нет. Семейства Bregmacerotidae, Brotulidae, Ophidiidae, Lycodidae, Fierasferidae и некоторые другие объединены в группу Ophidioidea, сближаемую с группой Blennioidea подотряда Xenopterygii из отряда Acanthopterygii. Камбалы являются особым подотрядом (Heterosomata) отряда Acanthopterygii, сближаемым с Anacanthini. Семейство Ammodytidae отнесено к подотряду Rhegnopteri, сближаемому с подотрядом Percesoces.
Boulenger (1901 : 268) объединяет тресковых вместе с семействами Trachinidae, Callionymidae и Nototheniidae в подотряд Jugulares, относимый им к отряду Acanthopterygii по общности положения у них лопаточного отверстия между scapula и coracoideum и брюшных плавников на горле. Эта система, как увидим ниже, признания не получила, так как сближать рыб в один подотряд только по положению брюшных плавников и лопаточного отверстия невозможно.
В 1903 г. Regan (1903 : 459) определяет Anacanthini следующим образом: parietalia отделены друг от друга посредством supraoccipitale; prooticum и occipitalia lateralia разделены увеличенным opisthoticum; плечевой пойс прикреплен к черепу; mesocoracoideum нет; непарные и брюшные плавники без колючих лучей (за исключением первого луча спинного плавника у некоторых Macruridae); брюшные плавники под грудными или впереди них; тазовый пояс позади ключичного симфизиса и прикреплен к нему лишь при помощи связки; жабры гребневидные; плавательный пузырь не соединен с кишечником.
Положению лопаточного отверстия между scapula и coracoideum Regan не придает существенного значения, так как оказалось, что у Gadomus longifilis из семейства Macruridae отверстие это находится в scapula. Точно так же и у Trematomus из семейства Nototheniidae, которое Boulenger по положению лопаточного отверстия между scapula и coracoideum сближал с семействами Anacanthini, отверстие это оказалось в scapula.
К Anacanthini Regan относит семейства Gadidae, Macruridae и Muraenolepidae. Роды Merluccius и Raniceps, выделяемые некоторыми в особые семейства, включены им в семейство Gadidae. К семейству Gadidae относит Regan и род Bregmaceros, у которого, по его исследованиям, кости брюшного пояса слабо прикреплены к плечевому поясу, хвостовой плавник симметричный и лопаточное отверстие находится между scapula и coracoideum, а не в scapula, как считали Jordan и Evermann (1898 : 2526), выделявшие Bregmaceros в самостоятельное семейство и сближавшие его с семействами Brotulidae и Ophidiidae.
В настоящее время система Anacanthini Regan'a является общепринятой. С небольшими изменениями принята она и Jordan'oм (1923 : 162).
СИСТЕМА
Goodrich, принимая в Vertebrate Craniata (1909 : 478) Anacanthini, называемых им Gadiformes, в объеме и определении Regan'a, обращает внимание на давно известную (Stannius, 1854 : 63), но забытую особенность строения их межглазничной области и передней части мозга, считая эту особенность наиболее характерным для них признаком.
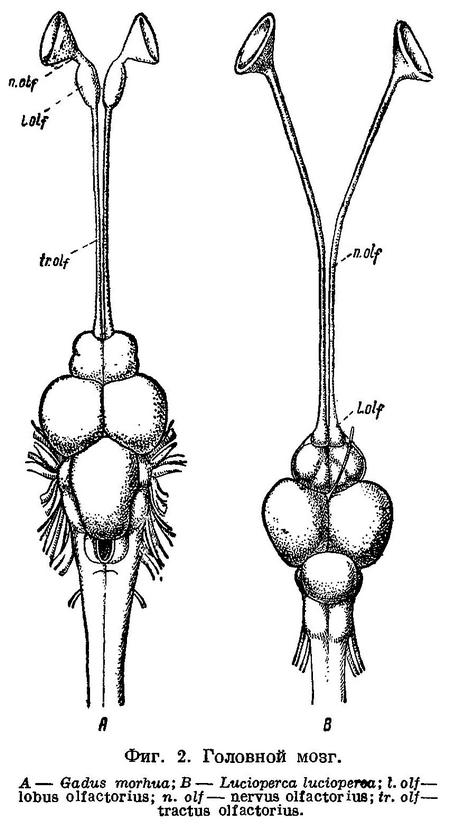
Черепная полость у Gadiformes продолжена вперед над septum interorbitale в виде узкого канала, где расположены обонятельные нервы и через посредство которого черепная полость оказывается сближенной с носовыми капсулами. Благодаря этому, обонятельные нервы у Gadiformes не проходят через орбиту и на всем протяжении расположены друг около друга (фиг. 1, А) в отличие от громадного большинства остальных Teleostei, у которых обонятельный нерв каждой стороны проходит через орбиту и отделен межглазничной перегородкой от нерва другой стороны (фиг. 1, С). Исключением среди Teleostei, подобно Gadiformes, являются далеко стоящие от них семейства Galaxiidae (Swinnerton, 1903 : 61), Siluridae, Cyprinidae (Sagemehl, 1891 : 489), некоторые Characinidae (Sagemehl, 1885 : 1) и Plectognathi (Diodon) (Owen, 1846 : 184, fig. 47), у которых межглазничная область устроена сходным образом. Кроме того, lobi olfactorii у Gadiformes далеко отстоят от переднего мозга, располагаясь в канале над septum interorbitale около носовых капсул, вследствие чего n.n. olfactorii у них очень короткие, а с передним мозгом lobi olfactorii соединяются при помощи длинных tractus olfactorii (фиг. 2, А), что среди Teleostei известно лишь у Galaxiidae (Swinnerton, 1903 : 63), некоторых Cypriniformes (Sagemehl, 1885 : 74; 1891 : 489) и Plectognathi (Owen, 1846 : 184).
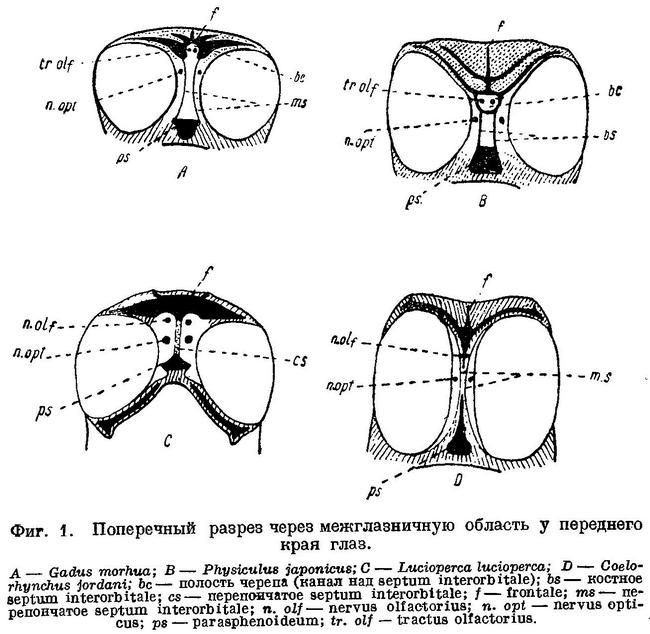
Помимо упомянутых семейств и отрядов рыб, обонятельные нервы, по указанию Dietz'a (1913 : 86), не проходят через орбиту еще у Liparis и Cyclopterus. В межглазничную же область полость черепа продолжена только у Liparis, у которого она здесь такой же ширины, как и сзади, и доходит до самых носовых капсул; что касается Cyclopterus, то у него полость черепа оканчивается спереди на уровне заднего края глаз, далее же обонятельные нервы разделены хрящом, заполняющим всю межглазничную область. Обонятельные нервы через орбиту не проходят у ряда других исследованных нами представителей подотряда Scleroparei (Муохоcephalus, Cottocomephorus, Comephorus, Sebastes, Trigla, Occa), куда относятся обе упомянутые выше рыбы, и Blennioidei (Blennius, Lumpenus, Ophidion), но полость черепа у всех них в межглазничную область не продолжена, за исключением Myoxocephalus, у которого она в виде канала тянется до самых носовых капсул. Однако, что является самым существенным, lobi olfactorii у всех перечисленных рыб расположены непосредственно около переднего мозга и с носовыми капсулами соединены при помощи длинных n.n. olfactorii, которые расположены между перепонкой septum interorbitale и костями или хрящом межглазничной области (см. фиг. 1, D) и через орбиту, таким образом, не проходят. (Указание Dietz'a (1921 : 436) на положение lobi olfactorii у Cyprinidae (фиг. 2) около переднего мозга и у Liparis (фиг. 5) около носовых капсул неверно: как это общеизвестно, у Cyprinidae lobi olfactorii находятся около носовых капсул, у Liparis, по нашим исследованиям, — около переднего мозга. Что касается скелета хвостового плавника, то у Gadiformes он совершенно иного устройства, чем у Cyclopterus, Liparis и Cottus, сближаемых Dietz'eм по этому признаку с Gadiformes, хотя у этих рыб он несколько менее асимметричен, чем у других Scleroparei. Наиболее существенный признак Gadiformes — большое opisthoticum с отверстием для n. glossopharyngeus Dietz не рассматривает.) Канал над septum interorbitale с расположенными в нем обонятельными нервами имеется, как обнаружено нами, и у сем. Brotulidae (Hoplobrotula armata), у представителей которого он, как и у сем. Cyprinidae, костный, но lobi olfactorii тесно примыкают к переднему мозгу, а к носовым капсулам тянутся по каналу длинные n.n. olfactorii.
Таким образом, межглазничная область иногда даже у одной и той же группы рыб устроена довольно разнообразно и в некоторых группах существенного значения как систематический признак иметь не может. Что касается положения lobi olfactorii, то около носовых капсул они расположены, насколько это известно в настоящее время (см. Lissner, 1918 : 127), только у семейств Galaxiidae, Cyprinidae, Siluridae, у некоторых Characinidae, у отряда Plectagnathi и у Gadiformes. На некотором расстоянии от носовых капсул, приблизительно посередине между ними и передним мозгом, расположены lobi olfactorii из Cyprinidae лишь у Carassius и из Gadidae — у Raniceps, вследствие того, что в передней части канала около носовых капсул у этих рыб разрастается хрящ и тем самым lobi olfactorii как бы отодвигаются от носовых капсул. Весьма характерно, что у Brotulidae lobi olfactorii расположены около переднего мозга, несмотря на наличие у них в межглазничной области канала, доходящего до самых носовых капсул. Возможно, что и у тех Characinidae, у которых lobi olfactorii расположены не около носовых капсул, это обусловлено разрастанием здесь хряща.
Lobi olfactorii расположены около носовых капсул, однако, не у всех Gadiformes. У Macrurus cavernosus (Pfüller, 1914 : 48, pi. 1—2), Coelorhynchus jordani, Coelorhynchus coelorhynchus, Coelorhynchus japonicus, Trachirinchus trachirinchus, Coryphaenoides rupestris, Coryphaenoides altipinnis и Abyssicola macrochir (наши исследования) из семейства Macruridae lobi olfactorii тесно примыкают к переднему мозгу, полость черепа в виде канала над septum interorbitale далеко не продолжена, вследствие чего tractus olfactorii отсутствует, а n.n. olfactorii длинные. Кроме того, у всех них оба обонятельных нерва, расположенные вначале друг около друга между перепонками septum interorbitale правой и левой сторон черепа, затем разделяются при помощи frontale и mesethmoideum и проходят к носовым капсулам, располагаясь между этими костями и перепонкой septum interorbitale и минуя, таким образом, орбиту (фиг. 1, В). У семейства Gadidae вдоль черепа с нижней стороны frontalia тянутся два костных гребня, ограничивающие с боков канал над septum interorbitale. У семейства Macruridae посередине черепа с нижней стороны frontalia тянется один непарный гребень, вдоль которого с боков расположены n.n. olfactorii. Нужно полагать, что подобным же образом расположены lobi olfactorii и устроена межглазничная область и у остальных представителей этого семейства. Следовательно, по одним из наиболее важных признаков — положению lobi olfactorii и строению межглазничной области — семейство Macruridae существенно отличается от других семейств Gadiformes. Из признаков, свойственных семейству Macruridae и другим семействам Gadiformes, следует отметить увеличенное opisthoticum с отверстием для n. glossopharyngeus (В этом отношении из Macruridae нами исследован только один вид — Coelorhynchus anatirostris Jord. et Gilb.); остальные признаки свойственны многим другим группам Teleostei.
Таким образом, по некоторым признакам семейство Macruridae должно быть отнесено к Gadiformes, по другим признакам оно должно быть из этого отряда выделено. И то и другое будет справедливо почти в одинаковой степени. Однако, после исключения из Gadiformes семейства Macruridae многие признаки (положение лопаточного отверстия между scapula и coracoideum, только мягкие лучи в непарных плавниках, положение брюшных плавников впереди грудных), не считаемые Regan'ом характерными, становятся общими для всех семейств отряда, и Gadiformes (Anacanthini) можно считать действительно мягкоперыми, объединенными в отряд большим количеством признаков. Поэтому обширное семейство Macruridae, повидимому распадающееся в свою очередь на несколько семейств, следует выделить в самостоятельный отряд, близкий, с одной стороны, к Gadiformes, с другой — к некоторым Acanthopterygii.
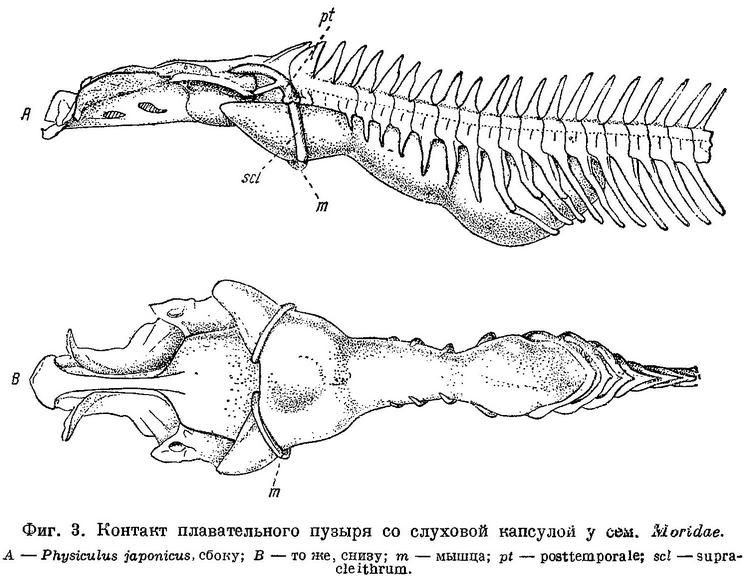
По исследованиям Stannius'a (1844 : 63) и по Goodrich'у (1909 : 478), septum interorbitale и нижняя часть канала n.n. olfactorii у Gadiformes перепончатые (фиг. 1, А). По нашим исследованиям, у значительной части представителей семейства Gadidae канал n.n. olfactorii целиком, a septum interorbitale на значительном протяжении костные (фиг. 1, В). Кроме того, у рыб, с костной septum interorbitale и каналом n.n. olfactorii, neurocranium спереди открывается лишь узким отверстием, будучи почти замкнут расширением и соединением parasphenoideum и frontalia, которые образуют и septum interorbitale. В neurocranium у рыб с костной septum interorbitale сзади с каждой стороны от foramen magnum имеется по большой, затянутой перепонкой фонтанелли, через которые при помощи двух длинных рожкообразных выростов плавательный пузырь находится в контакте с ушными капсулами (фиг. 3, А и В). Соединение плавательного пузыря с ушной капсулой у одного из представителей семейства Gadidae — Lotella bacchus было обнаружено давно (Parker, 1883 : 234), но оставалось мало известным. О том, что подобное соединение плавательного пузыря с ушной капсулой имеется и у некоторых других тресковых рыб, не было известно, и поэтому значения для системы эта важная анатомическая особенность не получила. Целиком костный канал n.n. olfactorii, костное на значительном протяжении septum interorbilale, фонтанелли сзали neurocranium и соединение через них плавательного пузыря с ушными капсулами, — все эти признаки имеют весьма существенное значение, и роды, которые обладают ими, должны быть выделены в особое семейство Моridae.
К семейству Moridae относятся роды: Mora, Lepidion, Uraleptus, Lotella, Physiculus, Laemonema, Antimora и ряд других.
Таким образом, отряд Gadiformes (Anacanthini) состоит из семейств Gadidae, Moridae, Bregmacerotidae. Последнее близко к семейству Gadidae, но по ряду признаков должно быть выделено в самостоятельное семейство. Что касается рода Мuraenolepis, который Regan (1903 : 466) выделяет в особое семейство Muraenolepidae и причисляет к отряду Gadiformes (Anacanthini), то по строению переднего мозга и черепа он не отличается от Gadiformes, но благодаря увеличенному числу грудных radialia (не менее 10) и некоторым другим особенностям (жаберные отверстия узкие, простирающиеся вверх не выше уровня основания грудных плавников, чешуи удлиненные и расположены под углом друг к другу; детальнее см. Световидов, 1939 : 583) он занимает несколько обособленное положение. Увеличенное число грудных radialia имеет важное значение и дает основание для выделения сем. Muraenolepidae в особый подотряд отряда Gadiformes. Таким образом, отряд Gadiformes разделяется на два подотряда: 1) Muraenolepidoidei с одним семейством Muraenolepidae и 2) Gadoidei с семействами Moridae, Bregmacerotidae и Gadidae. Последнее наиболее естественно, по строению взрослых, личинок и икринок, разделяется на подсемейства Lotinae, Мerlucciinae и Gadinae.
Происхождение и родство Gadiformes со всеми сближаемыми, с ними семействами и группами рыб весьма отдаленно. На отдаленность Gadiformes от Pleuronectiformes (Heterosomata) наиболее решительно и достаточно убедительно указывают Соре (1872 : 341) и Cunningham (1897 : 498), выводы которых Boulenger (1902 : 296) и Regan (1910а : 484) подкрепили новыми анатомическими данными. Regan (1903 : 460), указавший ряд новых признаков, хорошо отделяющих Gadiformes от сближаемых с ними семейств Blenniidae, Zoarcidae, Brotulidae, Ophidiidae и других, и Boulenger (1904 : 646, 702, 703) считают, что сходство между этими двумя группами рыб должно скорее быть приписано конвергенции, чем близкому родству. Позднее (1910 : 11) Regan высказывает предположение о происхождении Gadiformes от примитивных Iniomi, родственных ныне живущим Aulopidae. На отдаленность от Gadiformes семейства Aulopidae, имеющих соприкасающиеся parietalia, не разделенные посредством supraoccipitalia, и иное устройство жаберной крышки, указал Gregory (1933 : 207, 380, fig. 85). Кроме того, следует добавить, что канала над septum interorbitale у Aulopidae нет и n.n. olfactorii проходят через орбиту. Наиболее же существенным отличием семейства Aulopidae, равно как и других сближаемых с Gadiformes семейств и групп рыб, является положение lobi olfactorii около носовых капсул. Эта важная анатомическая особенность Gadiformes, характерная лишь для далеко стоящих от них Cyprinidae и Galaxiidae, указывает на раннее отщепление их от ныне живущих групп рыб общего с ними происхождения. При этом следует отметить, что некоторые характерные морфологические особенности, в частности дифицеркальный хвост (Whitehouse (1935 : 70) считает его гомоцеркальным и у взрослых. Barrington (1935 : 270) предлагает для хвостов такого типа название псевдокавдадьный.) и положение lobi olfactorii около носовых капсул, а не около переднего мозга, представляют собой явления вторичные: как известно, у молодых тресковых хвост гомоцеркальный, a lobi olfactorii, по указанию Т. С. Расса (Световидов, 1937 : 1288), у них располагаются чрезвычайно близко от переднего мозга. Л. С. Берг (1940 : 284), помещая Gadiformes в системе почти сразу за Cypriniformes, ниже Perciformes, считает их как низкоорганизованный отряд, происшедший от форм, родственных Pachycormidae. Для Gadidae он дает несколько иное, чем принятое здесь, разделение на подсемейства, основанное на строении только взрослых; в особые подсемейства им выделяются Eleginus (парапофизы расширены и содержат выросты плавательного пузыря) и Raniceps (lobi olfactorii от носовых капсул несколько удалены; о вторичном характере этого у Raniceps и Carassius см. выше).
Строение каналов боковой линии на голове имеет важное филогенетическое значение. У Holostei и низших Teleostei (Cypriniformes, Clupeiformes) эти каналы расположены в костях и сообщаются с наружной средой посредством узких отверстий, пронизывающих местами канал (Goodrich, 1930 : 294—295). У более высокоорганизованных Teleostei кожные кости погружаются глубже под кожные покровы, каналы менее замкнуты в костях и, как у Gadiformes, представляют бороздку, прикрытую гребнями и лишь частично замкнутую в костный канал. Слизевая ямка у мальков менее 60 мм длины, повидимому, спереди не замкнута, так как у мальков длиной 60—65 мм средние и боковые гребни frontalia соединены при помощи перепонки, которая у рыб размерами 85 мм и более окостеневает. Как происходит развитие остальных признаков — неизвестно.