(Alosa Linck, 1790) Род Алозы, Каспийско-черноморские сельди, Genus Alosa Linck, 1790 (River herrings) 22 вида
Словарь названий пресноводных рыб СССР. Г.У.Линдберг и А.С.Герд 1972 г.
3.1.0. A1оsa Cuvier, 1829 — Каспийско-черноморские сельди.
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
Род Алозы (Alosa) имеет в этой группе
особенно важное значение. Для видов
этого рода характерны сильно сжатое с боков
тело с приостренным зубчатым брюшным
килем; две удлиненные чешуйки
— «крылышки» — при основании верхней и
нижней лопастей хвостового плавника;
радиальные бороздки на крышечной кости;
заметная медиальная вырезка в верхней
челюсти; сильно развитые жировые веки
на глазах. С каждой стороны тела обычно
имеется по темному пятну за верхним
краем жаберной крышки, за которым у
некоторых видов часто следует ряд из
нескольких пятен; иногда, кроме того, под
этим рядом бывает второй и изредка
третий из меньшего числа пятен. Очень
характерны для разных видов и форм алоз
различия в форме и числе жаберных
тычинок, которые соответствуют различиям
в характере пищи. Малочисленные короткие и толстые жаберные тычинки
свойственны хищным сельдям,
многочисленные тонкие и длинные — планктоноядным
сельдям. Количество жаберных тычинок
на первой дуге у алоз бывает от 18 до 180.
Количество позвонков 43—59.
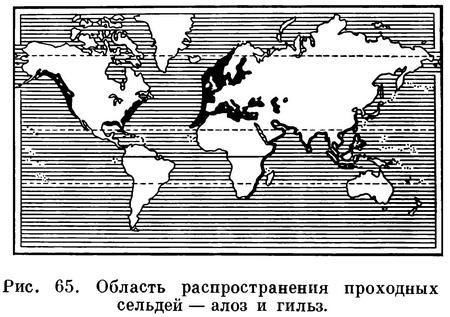
Алозы распространены в прибрежных
умеренно теплых водах бассейна
Атлантического океана в северном полушарии,
а также в Средиземном, Черном и
Каспийском морях. В этом роде насчитывается
14 видов, группируемых в два подрода:
10 видов основной формы рода настоящих
алоз (Alosa) и 4 вида помолобов (Pomolobus). У настоящих алоз высота щеки
больше ее длины, у помолобов равна длине
или меньше ее.
Два вида настоящих алоз живут в водах
восточного побережья Северной Америки
(Alosa sapidissima, Alosa ohioensis), два —
у западных берегов Европы, Северной
Африки и в Средиземном море (Alosa alosa,
Alosa fallax), два вида — в бассейнах
Черного и Каспийского морей (Alosa caspia,
Alosa kessleri), четыре вида — только в
Каспийском море (Alosa brashnikovi, Alosa saposhnikovi, Alosa sphaerocephala, Alosa curensis). Все
четыре вида помолобов (Alosa (Pomolobus)
aestivalis, Alosa (Pomolobus) pseudoharengus, Alosa (Pomolobus) mediocris, Alosa (Pomolobus) chrysochloris) живут в водах
Америки. Многие виды алоз распадаются
на большее или меньшее число форм —
подвидов, рас и т.д. По биологии
размножения различимы четыре группы видов
и форм рода алоза: проходные,
полупроходные, солоноватоводные и
пресноводные. Проходные живут в море, а для
нереста поднимаются в верхнее и среднее
течения рек (анадромные проходные);
полупроходные откладывают икру в
низовьях рек и в прилегающих предустьевых
слабосоленых участках моря;
солоноватоводные живут и нерестуют в солоноватой
морской воде. Некоторые атлантическо-средиземноморские проходные виды
образуют и местные озерные формы (подвиды),
постоянно живущие в пресной воде. В
водах Америки, Западной Европы,
Средиземноморского и Черноморско-Азовского
бассейнов живут проходные и
полупроходные виды, а также их пресноводные
формы; в Каспийском бассейне —
проходные, полупроходные и солоноватоводные
виды. В отличие от атлантическо-средиземноморских алоз черноморско-азовские
и каспийские не образуют озерные
пресноводные формы; при этом среди алоз Черноморско-Азовского бассейна
представлены три проходных и один полупроходной
вид, а в Каспийском море — один
проходной (2 формы), один полупроходной
(4 формы) и четыре солоноватоводных
вида.
У черноморских и каспийских алоз икра
созревает и выметывается в три порции,
с промежутками между выметами в 1 —1,5
недели. Количество икринок в каждой
порции составляет обычно от 30 до 80 тыс.
Икринки у видов рода Алоза
полупелагические, всплывающие на течении,
или донные, частью слабо прилипающие
(у американских помолобов и у
каспийского ильменного пузанка). Оболочка
полупелагических икринок тонкая, у
донных более плотная и импрегнированная
прилипшими частицами ила. Подобно
икринкам сардин икринки алоз имеют
большое или среднее кругожелтковое
пространство, но в отличие от сардин не
содержат, как правило, жировой капли в
желтке. Величина икринок у разных видов
различна: от 1,06 у большеглазого
пузанка до 4,15 мм у волжской сельди.
Черноморско-каспийские алозы
представлены тремя видами — пузанками (Alosa caspia), кесслеровскими сельдями (Alosa
kessleri) и бражниковскими сельдями (Alosa brashnikovi), распадающимися на ряд
подвидов и форм.
Обитатели бездны. Уильям Кроми 1971 г.
Знания, полученные путем исследований, позволяют человеку наиболее эффективно использовать богатства моря, сочетая свои усилия с усилиями природы. Одним из способов достичь этой цели является перемещение рыбы из одного района в другой. Хотя подобные мероприятия не раз оканчивались неудачей, несколько опытов оказались чрезвычайно успешными. В начале нынешнего столетия из реки Гудзон в район между рекой Сакраменто и заливом Сан-Франциско было доставлено около полутора миллионов мальков сельди-шэда. За 60 лет улов этой рыбы увеличился с 45 тонн до 2 тысяч тонн, и в настоящее время она водится вдоль всего побережья Соединенных Штатов от южной части Калифорнии до Аляски.
Фауна СССР. Рыбы. Том II, вып.1. Сельдевые (Clupeidae). А.Н.Световидов 1952 г.
9. Род ALOSA CUVIER
— Alosa Cuvier, Regne animal, ed. II, II, 1829 : 319 (тип: Alosa alosa).
— Alausa Valenciennes, in: Cuvier et Valenciennes, Hist. nat. poiss., XX, 1847 : 389 (тип: Alausa vulgaris = Alosa alosa).
— Clupeonella (non Kessler) Берг, Мат. позн. русск. рыбол., II, 3, 1913 : 10 (тип: Clupeonella caspia = Alosa caspia caspia).
— Caspialosa Берг, там же, IV, 6, 1915 : 4 (тип: Caspialosa caspia = Alosa caspia caspia).
— Alosa Regan, Ann. Mag. Nat. Hist. (8), XVIII, 1916 : 6 (обзор видов и близких родов).
— Paralosa Rоulе, Poiss. eau douces France, 1925 : 73 (тип: Alosa fallax; non Paralosa. Regan, 1916).
Верхняя челюсть с заметной медиальной вырезкой в межчелюстных костях. На глазах сильно развитые жировые веки, прикрывающие сзади и спереди большую часть их. При основании хвостового плавника на обеих лопастях по удлиненной чешуе (ala). Удлиненных последних двух лучей анального плавника и кожистой лопасти на вертикальной части ключицы нет. На крышечной кости ясные радиальные бороздки. Брюхо сжатое с боков, на всем протяжении от горла до начала анального плавника покрыто хорошо развитыми брюшными килевыми чешуями, образующими острый киль как позади, так и впереди брюшных плавников. Рот конечный, иногда нижняя или верхняя челюсть слегка выдаются вперед, большой: сочленение нижней челюсти с черепом позади вертикали заднего края глаза, задний конец верхнечелюстной кости простирается до вертикали середины глаза или почти достигает вертикали заднего края его. Зубы на межчелюстных, верхнечелюстных и нижнечелюстной костях и на нёбных и на языке есть (у Alosa kessleri kessleri, Alosa brashnikovi, Alosa saposhnikovi, Alosa sphaerocephala) или отсутствуют (у Alosa fallax и др., у Alosa fallax и Alosa caspia иногда отсутствуют на нижнечелюстных, у Alosa kessleri volgensis на нижнечелюстных слабые или отсутствуют), зубы на сошнике, расположенные на продольном гребне головки его, есть или отсутствуют (у Alosa fallax и близких видов, иногда у Alosa kessleri volgensis). Жаберные тычинки короткие и толстые или тонкие и длинные, числом от 18 до 180, у видов с большим количеством их тычинки верхней части жаберной дуги налегают на тычинки нижней части дуги. Отростки плавательного пузыря помещаются в prooticum и pteroticum. Начало спинного плавника над началом брюшных или последние расположены под передней третью спинного. Основание спинного плавника лежит в бороздке, образованной расширенными чешуями, более крупными у начала его. Позвонков 43—59. С каждой стороны тела по темному пятну за жаберной крышкой сверху, иногда за ними ряд из нескольких пятен, у некоторых, кроме того, под этим рядом есть второй и изредка третий из меньшего числа пятен. Икринки полупелагические, держатся во взвешенном состоянии в природных слоях воды, с большим околожелтковым пространством. Размеры в половозрелом состоянии 160—520 мм.
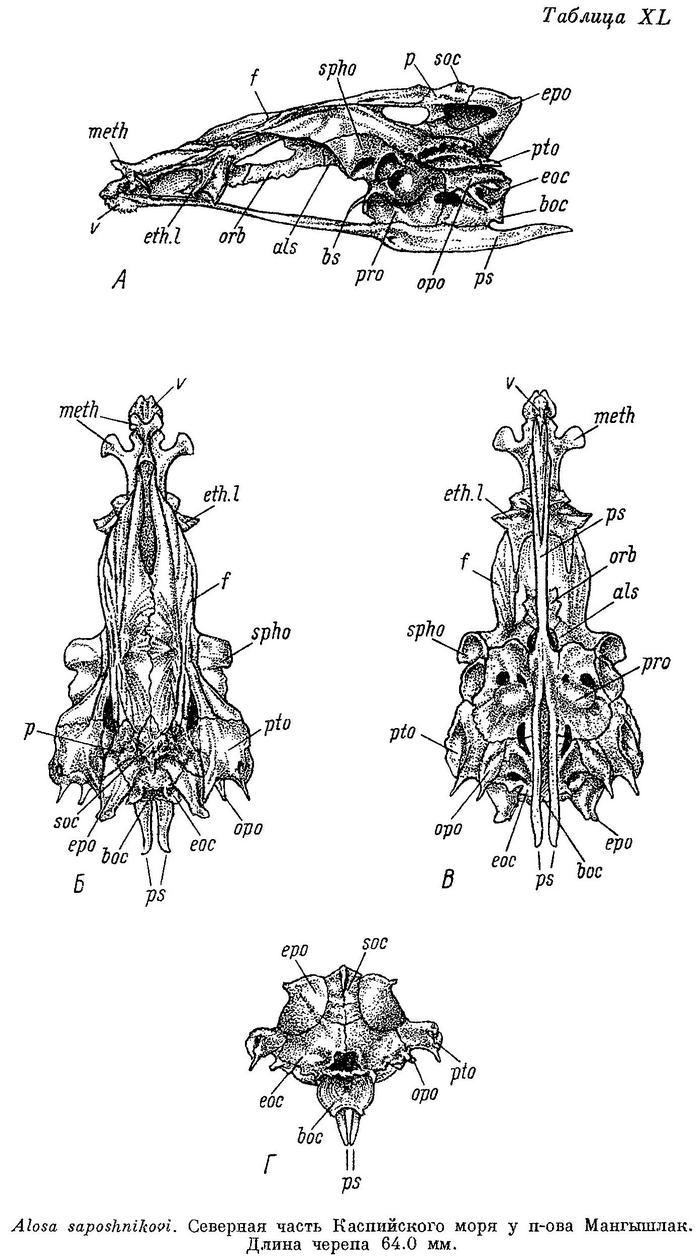
Сколет. Описание и рисунки черепа Alosa fallax см. у Мэтьюза (Matthews, 1887 : 274—292), Райдвуда (Ridewood, 1904а : 453—459, f. 122—124), Alosa sapidissima у Трэси (Tracy, 1920 : 439—474, pl. 1—3). Bulla prootica крупнее по размерам, чем bulla pterotica. Bulla prootica на черепе сбоку (табл. XL—LII, А) образует большое и выпуклое вздутие; вздутие bulla pterotica снаружи ни сбоку, ни сверху не выражено. Каждый отросток плавательного пузыря расположен в углублении вдоль переднего края occipitale laterale, сзади и отчасти сверху ограниченном гребнем; отверстие, через которое отросток входит в череп, расположено сверху, под opisthoticum. Fenestra auditiva небольшая, обычно овальной формы, далеко отстоит от bulla prootica. Спереди и от гребня вдоль отростка плавательного пузыря сзади. Fossa praeepiotica большая, несколько большая по размерам, чем foramen temporale, имеет форму треугольника, обращенного вершиной вниз, высота ее меньше длины. Foramen temporale сравнительно небольшое, длина его немного меньше длины fossa praeepiotica, наиболее высоко посередине. Сочленовная поверхность hyomandibulare разделена на две обособленные части на sphenoticum и pteroticum. Сошник и передняя часть парасфеноида почти прямые, передняя часть парасфеноида посередине сверху закругленная, крылья парасфеноида выдаются вниз под миодомом, нижний край их прямой или слегка изогнутый, заостренные на концах. Нисходящий отросток базисфеноида длинный, часто почти достигает парасфеноида. Задний край supraoccipitale и каждого epioticum на черепе сверху (Б) расходится под прямым углом. Pteroticum сзади оканчивается небольшим шипом. Передний конец каждого sphenoticum отходит в стороны, иногда немного направлен вперед или назад, на конце закругленный или с небольшим крючкообразным отростком сзади, короткий или более длинный, варьируя во всех этих отношениях в зависимости от формы и индивидуально. Поперечные отростки ethmoidalia lateralia выдаются из-под переднего конца лобных костей, значительно варьируют по форме и направлению. Передние отростки мезэтмоида направлены в стороны и немного вперед, прямые, немного закругленные на концах. Ширина черепа варьирует у разных видов, наибольшая ширина его содержится от 2.25 до 1.5 раз в длине, ширина лобных костей от 4.5 до 3.2 раз, наиболее широк череп у Alosa fallax. Сошник снизу черепа (В) закругленный, головка его с широким и высоким гребнем, несущим зубы, у некоторых (Alosa fallax и близкие виды, иногда у Alosa kessleri volgensis), у которых зубы на сошнике отсутствуют, гребень развит очень слабо, узкий и закругленный; передняя часть парасфеноида плоская, обычно более узкая сзади, крылья парасфеноида разделяются и сразу широко расходятся под серединой prooticum. Сзади на черепе (Г) fossae temporales posteriores отсутствуют, с каждой стороны между отростком opisthoticum и шипом pteroticum расположена fossa subtemporalis. На hyomandibulare сверху две отчетливо обособленные головки для сочленения с черепом. Quadratum с глубокой выемкой в верхнем крае, передний угол ее вытянут. Нижнечелюстная и сочленовная кости удлиненные и низкие, первая длиннее последней. Предкрышечная кость значительно уже широкой крышечной. Описание дано по многим скелетам почти всех видов и форм, встречающихся в наших водах, и по одному скелету Alosa fallax.
Солоноватоводные или проходные рыбы, последние образуют ряд пресноводных форм, 5 или, возможно, 6 видов с рядом подвидов в Каспийском, Черном и Азовском морях и 2—4 вида с несколькими подвидами у европейских и американских берегов Атлантического океана, в Балтийском, Средиземном и отчасти в Черном морях и впадающих в них реках (рис. 27).
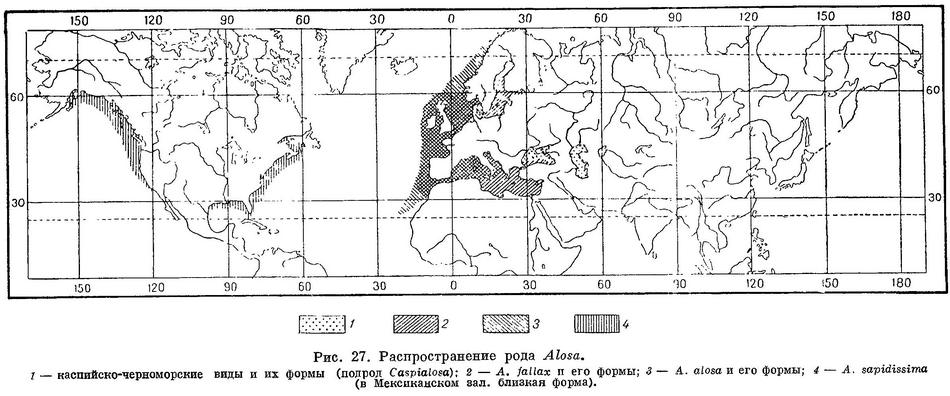
Относящиеся к этому роду виды разделялись на два рода — Alosa и Caspialosa по отсутствию зубов на сошнике и нёбных костях у первого и наличию их у второго. Однако у Alosa caspia и Alosa kessleri volgensis эти зубы развиты слабо, иногда почти не заметны, а у Alosa kessleri volgensis зубы на сошнике иногда могут отсутствовать совершенно, причем гребень вдоль головки сошника у таких рыб бывает даже менее развит, чем у Alosa fallax Такой же переходный характер имеют и другие признаки отличия (зубы на челюстных костях) между Alosa и Caspialosa. Вообще степень развития зубов на одних и тех же костях у разных видов, как указывалось выше, варьирует в широкой степени — от сильно развитых до слабых, едва заметных, или совершенно отсутствующих, как и у видов Sprattus. Различия в окраске тем более не могут быть признаны достаточными для разделения обоих родов, так как ряд темных пятен на теле, постоянно имеющийся у видов Alosa, имеют и некоторые формы Caspialosa. Самым существенным является то, что по строению черепа оба рода не имеют таких отличий, по каким различаются другие роды семейства (например Sardina, Sardinops и др. из подсем. Clupeinae). По всем признакам виды обоих родов представляют близко родственную группу, в которой виды, относимые к Alosa, по комплексу признаков, положенных в основу системы, являются более прогрессивными по сравнению с видами, относимыми к Caspialosa (см. «Система семейства и принципы классификации»). Череп видов, относимых к Alosa, лишь несколько более широк, чем у видов, относимых к Caspialosa. Это наблюдается и у ряда других родов, в частности Clupeonella, у более прогрессивного вида которого Clupeonella delicatula череп значительно шире, чем у менее прогрессивного Clupeonella grimmi. Таким образом, имеющиеся морфологические различия не дают оснований для разделения обоих родов. Особенности распространения, история развития (см. «Географическое распространение») и имеющиеся небольшие различия дают возможность считать их самое большее за подроды.
Все виды рода по характерному внешнему виду делятся на две группы: 1) пузанков с более высоким, сжатым с боков и укороченным в хвостовой части телом, большой и высокой, клиновидно сжатой с боков головой и длинными грудными плавниками и 2) сельдей с низким, не сжатым с боков и не укороченным в хвостовой части телом, более короткой, низкой и не сжатой клиновидно с боков головой и короткими грудными плавниками. По ряду морфологических особенностей пузанки являются более примитивными видами, чем сельди; экологически эти группы различаются тем, что виды и формы сельдей являются более реофильными, пузанков — стагнофильными. Среди последних нет настоящих проходных видов и форм, те же формы, которые заходят для размножения в реки, нерестуют в отличие от проходных сельдей не в основном русле, а в ильменях, на полоях и т. п.
Кесслер был первым, если не считать неопределенных высказываний Палласа (Zoogr. rosso-asiatica, III, 1811 : 205), Бэра (Bull. CI. phys.-math., XVI, 1858 : 327, 332) и некоторых других, кто сопоставил каспийских и черноморско-азовских сельдей и указал на близость их. Впервые им была отмечена (Тр. СПб. общ. ест., I, 1870 : 76) близость к каспийским проходным черноморско-азовской проходной сельди (Clupea pontica auct.), причем затруднительно сказать, какую из каспийских проходных сельдей он имеет в виду под названиями Clupea caspio-pontica и Clupea rossica (nomina nuda). Под именем Clupea pontica и Alosa caspia им, несомненно, смешаны (цит. раб. : 281, 282) Alosa kessleri kessleri и Alosa kessleri volgensis, а с последним, кроме того, и Alosa caspia. Один из экземпляров, пойманных в сентябре 1809 г. у Ярославля относится к Alosa caspia caspia. Несколько позднее Кесслер (там же, V, 1, 1874 : 301) еще в более определенной форме указывает на близость каспийских и черноморско-азовских форм Alosa caspia, причем совершенно правильно отмечает и признаки отличия черноморской разности, как он ее называет (более сильно развитые зубы, меньшее количество жаберных тычинок и др.). Что касается Clupea pontica, то под этим названием он, повидимому смешивает, судя по числу жаберных тычинок (35—55), из черноморско-азовских сельдей Alosa kessleri pontica и Alosa brashnikovi maeotica, а из каспийских, для которых он приводит еще меньшее число жаберных тычинок (25—28), по мнению Гримма (Астраханская селедка, СПб., 1887 : 5), исследовавшего экземпляры, определенные Кесслером, «3 различных формы — 2 самостоятельных вида и продолговатую разность предыдущего вида» (Clupea caspia), т. е. современные Alosa brashnikovi brashnikovi, Alosa kessleri kessleri и Alosa kessleri volgensis. После Кесслера сельди обоих бассейнов были сопоставлены Бородиным (Ежегодн. Зоолог. муз. Акад. Наук, 1, 1896 : 82—84), которым было указано (Вестн. рыбопр., XIX, 3, 1904 : 176) на близость каспийских морских сельдей к черноморско-азовской и установлен общий для них вид Clupea caspio-pontica, в качестве разновидностей которого он рассматривает Alosa saposhnikovi, Alosa brashnikovi brashnikovi, Alosa brashnikovi grimmi и Alosa brashnikovi maeotica. Самое название caspio-pontica Borodin является, как это отмечено Бергом (Мат. позн. русск. рыбол., II, 3, 1913 : 19), синонимом saposhnikovi Grimm 1887. Кроме того, это название есть nomen praeoccupatum (см. выше caspio-pontica Kessler). Несколькими годами ранее последней из упомянутых работ Бородина, Гримм (Вестн. рыбопр., XVI, 2, 1901 : 57), описывая новые формы сельдей из Азовского моря, сделал также некоторое сопоставление их с каспийскими. Позднее сельди обоих бассейнов изучались друг от друга отдельно [каспийские Бергом (Мат. позн. русск: рыбол., II, 3, 1913), Киселевичем (Каспийско-волжские сельди, I, 1923), Мейснером и Михайловской и некоторыми другими; черноморско-азовских изучали Antipa (Denkschr. Akad. Wiss. Wien, math.-naturw. Kl., LXXVIII, 1905, 1906), Браунер (Тр. Бессарабск. общ. ест., II, 2, 1912), Исаченко (Тр. Всеукр. Гос. черном.-азовск. научно-промысл. оп. ст., I, 1925), Александров (Тр. Керченск. ихт. лаб., I, 1, 1926), Borcea (Ann. Sci. Univ. Jassy, XIX, 1934; XXII, 1936; XXV, 1939), Майорова (Тр. Научн. рыбохоз. биолог. ст. Грузии, II, 1939)] и лишь некоторые сравнительные замечания о каспийских и черноморско-азовских сельдях были сделаны Киселевичем (Изв. Отд. рыбол. и научно-промысл. исслед., I, 1, 1918 : 21), отметившим, что последние отличаются от каспийских меньшими размерами. При этом из каспийских сельдей изучались преимущественно встречающиеся в северной части этого моря (Киселевич, цит. раб.; Аверинцев и Сыч, Тр. Астрах. научн. рыбохоз. ст., VII, 3, 1930), а встречающиеся в западной части Черного моря (Antipa, l. с.; Исаченко, цит. раб.; Borcea, l. с.) — независимо от встречающихся в восточной части его и в Азовском море. Изучение каспийских сельдей по всему бассейну после работы Берга (1913) было предпринято в сравнительно недавнее время Мейснером и Михайловской (Михайловская, Докл. АН СССР, XXX, 6, 1941), исследовавшими обширные материалы и описавшими ряд новых форм, главным образом из южной части моря, причем система и систематические обозначения каспийских сельдей как Мейснером и Михайловской, так и большинством других авторов (Морозов, Тр. Научн. инст. рыбн. хоз., II, 4, 1928), в том числе и Киселевичем (Промысловые рыбы Волго-каспийского района, Астрах., 1926; Сельди северного Каспия, Сталингр., 1937), предложившим ранее (цит. раб., 1923) считать всех каспийских сельдей формами одного вида (Caspialosa caspia), были приняты, согласно Бергу (цит. раб., 1913; Рыбы пресных вод, 1923; там же, I, 1932). Несколькими годами позднее Майоровой (цит. раб.) были изучены сельди из восточной части Черного моря, Керченского прол. и низовьев Дона и произведено сравнение их с сельдями западной части моря по литературным данным (Исаченко, 1925), что дало, однако, возможность сделать правильные выводы относительно некоторых форм и систематических отношений черноморско-азовских сельдей. Вследствие отсутствия сравнительного изучения каспийских и черноморско-азовских сельдей после упомянутых выше работ Кесслера и др. система и систематические обозначения сельдевых, общих обоим бассейнам (роды Clupeonella и Caspialosa), оставались до сего времени крайне противоречивыми, находясь в противоречии с систематическими отношениями многих и других рыб, общих обоим бассейнам. Черноморско-азовских пузанков (Caspialosa tanaica auct. и считаемых до настоящего времени формами этого вида Caspialosa tanaica nordmanni и Caspialosa tanaica palaeostomi) и морскую (Caspialosa maeotica auct.) и проходную (Caspialosa pontica auct.) сельдей принято считать за отдельные виды от соответствующих им морфологически и биологически каспийских пузанков (формы Caspialosa caspia auct.) и морских (формы Caspialosa brashnikovi auct.) и проходных (Caspialosa kessleri auct. и близкая к ней Caspialosa volgensis auct.) сельдей, в то время как черноморско-азовскую и каспийскую тюльку принято считать идентичными формами (Clupeonella delicatula). Между тем ряд других рыб, общих Черному, Азовскому и Каспийскому морям, представлен в каждом из бассейнов подвидами. Таковы: Gobius melanostomus Pall., Gobius fluviatilis Pall., Proterorhinus marmoratus (Kessl.), Benthophilus macrocephalus magistri Iljin, Syngnathus nigrolineatus Eichw., Atherina mochon pontica Eichw. в Черном и Азовском морях и Gobius melanostomus affinis Eichw., Gobius fluviatilis pallasi Berg, Proterorhinus marmoratus nasalis (Filippi), Behtophilus macrocephalus (Pall.), Syngnathus nigrolineatus caspius Eichw., Atherina mochon pontica n. caspia Eichw. в Каспийском море. Самостоятельными подвидами представлена, в каждом из бассейнов и тюлька (Clupeonella delicatula delicatula и Clupeonella delicatula caspia).
Сравнительное изучение каспийских и соответствующих близких им морфологически и биологически черноморско-азовских сельдей дало возможность установить, что последние не являются самостоятельными видами, а лишь подвидами общих обоим бассейнам видов. Черноморско-азовские пузанки (Caspialosa tanaica tanaica, Caspialosa tanaica nordmanni и Caspialosa tanaica palaeostomi) отличаются от каспийских (форм Alosa caspia) не более, чем некоторые из последних между собой. Так, например, Alosa caspia knipowitschi отличается внешне и количеством жаберных тычинок, которых у него не менее 120, от некоторых других форм вида (у Alosa caspia salina жаберных тычинок не более 117, у Alosa caspia persica не более 102—104) значительно больше, чем черноморско-азовские пузанки от большинства каспийских. Еще меньше отличается от каспийских проходных сельдей (Alosa kessleri kessleri и Alosa kessleri volgensis) черноморско-азовская проходная сельдь (Caspialosa pontica auct.), имеющая, подобно черноморско-азовским пузанкам, наименьшее количество жаберных тычинок, a hiatus в числе их лишь с Alosa kessleri volgensis. Отличие черноморско-азовской проходной сельди от остальных каспийских проходных сельдей значительно меньше различий между Alosa kessleri kessleri и Alosa kessleri volgensis. Следует отметить, что упомянутая выше наиболее обособленная форма каспийских пузанков (Alosa caspia knipowitschi) имеет близкие формы, промежуточные по своим признакам (например Alosa caspia caspia) как между другими каспийскими формами, так и между наиболее резко отличающимися от них из черноморско-азовских пузанков (например Alosa caspia palaeostomi и др.). Черноморско-азовская морская сельдь (Caspialosa maeotica auct.) отличается от большинства каспийских морских сельдей (форм Alosa brashnikovi), в частности от типичной формы вида (Alosa brashnikovi brashnikovi) значительно меньше, чем некоторые из последних между собой. Так, например, Alosa brashnikovi grimmi имеет не более 27 жаберных тычинок, в то время как у Alosa brashnikovi kisselevitshi их не менее 29. Обе формы, что самое существенное, резко различаются строением жаберных тычинок и внешне — формой тела, окраской. Кроме того, Alosa brashnikovi grimmi резко отличается от большинства форм вида наличием мозолистого утолщения на нижней челюсти, имеющегося в менее развитой степени еще лишь у Alosa brashnikovi nirchi. Однако нет никаких оснований считать одну из этих двух форм, в частности наиболее уклоняющуюся — Alosa brashnikovi grimmi, за самостоятельный вид, так как обе эти формы вместе с остальными формами вида, промежуточными по своим признакам между ними, представляют одну группу, связанную сходством в основных морфологических и биологических особенностях и принимаемую в настоящее время всеми с полным основанием, подобно формам Alosa caspia, за единый вид. Различия между формами Alosa brashnikovi не более различий между многими локальными формами Zoarces viviparus L., некоторые из которых резко отличаются количеством позвонков. Так, например, форма из Роскильде-фиорд имеет не более 113 позвонков, а форма из восточной части Балтийского моря (Клайпеда) имеет не менее 116 позвонков (Johs. Schmidt, C.-R. Trav. Lab. Carlsberg, XIII, 3, 1917 : 363, 366). Резко различаются отдельные формы Zoarces viviparus и внешне, в особенности в строении выемки между спинным и хвостовым плавниками, у некоторых форм совершенно отсутствующей (Johs. Schmidt, l. с., fig. 16, 17, 18), так что, не зная промежуточных форм, наиболее уклоняющиеся по этой особенности формы следовало бы считать разными видами. Однако несмотря на резкие различия между некоторыми из этих локальных форм, все они вследствие наличия между ними переходных по своим признакам форм рассматриваются совершенно справедливо как составляющие единый вид. Формальная точка зрения в вопросах систематики, как видно из этих примеров, не всегда приемлема, так как, следуя ей, некоторые формы каспийских сельдей следовало бы считать отдельными видами, а морских и проходных сельдей черноморско-азовских и каспийских соединить в один вид, потому что нет достаточно резких различий между черноморско-азовкими морской и проходной формами сельдей. Несомненно, что при разграничении видов сельдей этого рода необходимо принимать во внимание помимо признаков морфологических и эколого-биологические особенности их и условия формообразования. Следовательно, считая вполне основательно формы Alosa brashnikovi единым видом, несмотря на резкие различия между некоторыми из них, черноморско-азовскую морскую сельдь (Caspialosa maeotica auct.) следует тем более признать за форму этого вида (Alosa brashnikovi maeotica). Принимая за единый вид формы Alosa caspia, различающиеся между собой менее, чем формы Alosa brashnikovi, мы должны считать и черноморско-азовских пузанков формами этого вида (Alosa caspia tanaica, Alosa caspia nordmanni, Alosa caspia palaeostomi). Волжская сельдь (Caspialosa volgensis auct.) и черноспинка (Caspialosa kessleri auct.), принимаемые всеми в настоящее время за отдельные виды, отличаются друг от друга не более, чем Alosa caspia knipovitschi от Alosa caspia salina и Alosa caspia persica, и значительно менее, чем Alosa brashnikovi grimmi от Alosa brashnikovi kisselevitshi. Рассматривая как единые виды каспийские и черноморско-азовские формы пузанка (Alosa caspia s. l.) и каспийские и черноморско-азовскую формы морской сельди (Alosa brashnikovi s. l.), мы должны признать за единый вид всех и проходных сельдей как каспийских (Alosa kessleri kessleri и Alosa kessleri volgensis (О Caspialosa volgensis kisselevitshi auct. (= Caspialosa volgensis imitans)), так и черноморско-азовскую (Alosa kessleri pontica). Таким образом, среди сельдей подрода Caspialosa следует различать комплекс форм морских, непроходных, составляющих вид Alosa brashnikovi s. l., комплекс форм проходных, составляющих вид Alosa kessleri s. l., и комплекс широко евригалинных форм, составляющих вид Alosa caspia s. l., одни из форм которого являются морскими, непроходными, размножающимися в море, другие — полупроходными, эстуарными. По правилам номенклатуры, формам морских сельдей следовало бы присвоить видовое название Alosa maeotica, а не Alosa brashnikovi, формам же проходных сельдей — Alosa pontica, а не Alosa kessleri, так как черноморско-азовские формы их описаны раньше каспийских. Однако поступить таким образом в угоду этим правилам было бы крайне неблагоразумно, так как за более изученными и более многочисленными каспийскими формами этих видов, в частности за морскими сельдями, уже прочно укоренились принятые нами названия и перемена их на другие причинит много неудобств. Необходимо сказать, как совершенно справедливо отмечено Л. С. Бергом (цит. раб., 1932 : 17), что «никакого непреложного «закона» приоритета нет, а есть правило, которым надлежит в разумных пределах пользоваться», не доводя его до абсурда. Кроме того, самостоятельными видами следует считать резко отличающиеся друг от друга и от других видов Alosa saposhnikovi и Alosa sphaerocephala, следуя в отношении этих двух видов Л. С. Бергу и принявшим его систему сельдей этого рода ряду других авторов. За самостоятельный вид, однако, провизорно, до более детального изучения этой малоизвестной сельди, следует признать и Alosa curensis.
Большинство форм основных каспийско-черноморских видов рода, рассматриваемых в нижеследующем как подвиды, не имеют резко обособленных ареалов, так как в зимнее время встречаются в одних и тех же местах. Так, например, Alosa caspia caspia и Alosa caspia salina, размножающиеся в северной части Каспийского моря, зимой держатся в южной части моря, где постоянно живут и размножаются Alosa caspia knipowitschi и Alosa caspia persica. Alosa caspia knipowitschi, нерестующий у западных берегов южной части Каспийского моря, осенью и зимой в небольшом количестве заходит на восток до Астрабадского зал., где весной нерестует Alosa caspia persica, повидимому, заходящий, в свою очередь, на восток до Пехлевийского зал. Равным образом и Alosa brashnikovi brashnikovi и Alosa brashnikovi agrachanica, размножающиеся в северной части Каспийского моря, зимуют в южной части моря, где живут и размножаются остальные 6 подвидов этого вида. Это же можно сказать и о Alosa kessleri kessleri и Alosa kessleri volgensis, встречаемых в море совместно, но для икрометания идущих раздельно и размножающихся в разных местах по течению Волги. Таким образом, все эти подвиды, не имея резко обособленных ареалов в течение главным образом зимнего времени, в период размножения достаточно обособлены, так как места икрометания их довольно строго локализованы. Каспийские формы Alosa kessleri Бергом в последнем издании «Рыб пресных вод» (1948) обозначены как infraspecies (например Caspialosa kessleri isp. volgensis), черноморская — как subspecies (Caspialosa kessleri pontica). Как каспийские, так и черноморские формы двух других упомянутых выше видов приняты им за подвиды. Учитывая отсутствие резкой обособленности ареалов не только у каспийских форм Alosa kessleri, но в такой же или у некоторых форм даже в большей мере и других упомянутых видов, за infraspecies следовало бы считать формы всех трех видов. Однако каспийские формы Alosa caspia и Alosa brashnikovi всеми авторами приняты за подвиды, обозначение их как subspecies или как infraspecies имеет лишь то различие, что в последнем случае между видовым и подвидовым названием ставится isp. Поэтому в нижеследующем формы всех упомянутых видов приняты за подвиды, хотя это обозначение в известной мере условно. Это тем более допустимо, что подобные же ареалы свойственны ряду подвидов и птиц. Так, в Закавказье (Азербайджанская ССР) на ареале местного подвида скворца Sturnus vulgaris caucasicus Lorenz зимует совместно с ним Sturnus vulgaris vulgaris L., населяющий большую часть восточной Европы. В тех же местах зимуют совместно подвиды белой трясогузки Motacilla alba alba (L.), населяющей Европейскую часть СССР и Зап. Европу, и Motacilla alba dukhunensis Sykes, гнездящихся в Сибири на восток до басс. Лены и к югу до прикаспийских степей и Кавказа. В Закавказье же и в Таджикистане зимуют совместно подвиды полевого жаворонка Alauda arvensis cantarella Bp., гнездящегося в южной части СССР, и Alauda arvensis dulcivox Brooks, гнездящегося в западной Сибири. Из примеров, которых можно было бы привести и больше, видно, что все эти подвиды не являются, строго говоря, подвидами, однако считаются таковыми.
Некоторые авторы (Smitt 1895; Otterstrøm и др.) рассматривают Alosa fallax, Alosa alosa и Alosa sapidissima как формы одного вида. Однако Alosa fallax резко отличается от всех видов как внешне, так и количеством жаберных тычинок. Крайне сходные внешне Alosa alosa и Alosa sapidissima, с одинаково клиновидно сжатой с боков и высокой головой, напоминающей по форме голову Alosa caspia, не различаются резко и по количеству жаберных тычинок (Alosa alosa от 90 до 130, у Alosa sapidissima от 93 до 120), являясь, повидимому, формами одного вида.
Видовая принадлежность сельдей рода Alosa определяется обычно без особых затруднений, хотя иногда и встречаются экземпляры как промежуточные по своим признакам, так, повидимому, и гибридного, происхождения, отнесение которых к тому или иному виду бывает затруднительно. Что же касается подвидов, в особенности Alosa brashnikovi, то здесь такие особи, относительно которых бывает затруднительно сказать, к какой форме данного вида их отнести, довольно нередки.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (10). Зубы на нёбных костях есть, у некоторых очень слабые, почти незаметные. За жаберной крышкой сверху обычно темное пятно, у некоторых сзади него на боках тела по ряду темных пятен. Каспийское, Черное и Азовское моря и впадающие в них реки (подрод Caspialosa).
2 (7). Тело пузанковое, высокое и сжатое с боков, с укороченным хвостовым стеблем и длинными грудными плавниками. Голова большая и высокая, клиновидно сжатая в нижней части с боков.
3 (6). Жаберных тычинок на первой дуге не более 45. Тычинки толстые и короткие, равны или длиннее жаберных лепестков. Зубы хорошо развитые.
4 (5). Верхний и нижний профили головы прямые. Нижняя челюсть выдается вперед, верхний край нижней челюсти прямой. *1. Alosa saposhnikovi (Grimm)
5 (4). Верхний и нижний профили головы закругленные, в особенности впереди глаз. Челюсти одинаковой длины, верхний край нижней челюсти характерной полулунной формы. *2. Alosa sphaerocephala (Berg)
6 (3). Жаберных тычинок на первой дуге не менее 50. Тычинки тонкие и длинные, значительно длиннее жаберных лепестков, с необломанными концами, образующими на нижней половине дуги выпуклую линию. Зубы развиты очень слабо, на нижней челюсти иногда совершенно незаметные на ощупь. *3. Alosa caspia (Eichw.) s. l.
7 (2). Тело сельдеобразное, низкое и не укороченное в хвостовой области, не сжатое с боков, с короткими грудными плавниками. Голова низкая, не клиновидная, не особенно большая.
8 (9). Жаберных тычинок не более 47. Тычинки толстые и грубые, короче, равны или немного длиннее жаберных лепестков. Зубы хорошо развитые. Морские сельди. *4. Alosa brashnikovi (Borodin) s. l. (Слабо изученная Alosa curensis, которую автор не имел в достаточном количестве, в определительную таблицу не включена. Эта сельдь близка к Alosa brashnikovi, отличаясь более ранним половым созреванием (при 11—14 см).)
9 (8). Жаберных тычинок не менее 47, у каспийских от 59. Тычинки тонкие и длинные, длиннее жаберных лепестков или более толстые и короткие, с обломанными или целыми концами. Зубы более или менее развитые, иногда на нижней челюсти слабо заметные. Проходные сельди. *6. Alosa kessleri (Grimm) s. l.
10 (1). Зубы на нёбных костях отсутствуют. Кроме темного пятна за жаберной крышкой сверху есть всегда ряд таких же пятен на боках тела, ниже которого иногда имеется еще 1 или реже 2 ряда из меньшего числа пятен. По обе стороны Северной части Атлантического океана, Средиземное, Балтийское и отчасти Черное моря и впадающие в них реки (подрод Alosa s. str.).
11 (14). Жаберных тычинок не более 80, тычинки короткие, обычно равные лепесткам. Голова невысокая и не сжатая в нижней части клиновидно с боков, рыло заостренное, узкое. *7. Alosa fallax (Lacep.)
12 (13). Жаберных тычинок 37—43, в среднем около 40. Спинной плавник расположен ближе к голове, антедорсальное расстояние в среднем около 43.5% длины тела. Атлантические берега Европы, Балтийское море. *7b. Alosa fallax fallax (Lacep.)
13 (12). Жаберных тычинок 34—37, в среднем около 35. Спинной плавник отнесен несколько дальше назад, антедорсальное расстояние в среднем около 45% длины тела. Средиземное и Черное моря. *7а. Alosa fallax nilotica (Geoffr.)
14 (11). Жаберных тычинок не менее 85, тычинки тонкие и длинные, значительно длиннее жаберных лепестков. Голова высокая, клиновидно сжатая в нижней части с боков, рыло широкое. *6. Alosa alosa (L.).
15 (16). Жаберных тычинок 100—110. Черное море. *8а. Alosa alosa bulgarica Drensky.
16 (15). Жаберных тычинок 90—120. Атлантические и тихоокеанские берега Сев. Америки, у западных берегов Камчатки. *9. Alosa sapidissima (Wilson)
По положению спинного и прочих плавников, по окраске и некоторым другим особенностям виды рода Alosa располагаются следующим образом (табл. 5, рис. 5). Как видно из этой таблицы и рисунка, по положению спинного и других плавников, по ширине и длине головы, а также по степени развития темных пигментных пятен на боках тела виды рода Alosa располагаются в ряд, который, повидимому, является частью общего ряда подсем. Alosinae. Изменение всех рассматриваемых признаков происходит в общем закономерно, за исключением некоторых из них у Alosa alosa и Alosa sapidissima, что обусловлено в основном малым количеством исследованных рыб (4 экз. первого и 2 экз. второго). При этом следует отметить, что изменение положения всех плавников относительно переднего конца тела происходит в одном направлении, как у форм Clupea harengus, в отличие от подсем. Clupeinae и других видов этого подсемейства, у которых анальный плавник претерпевает перемещение в обратном направлении, чем спинной и брюшные. Однако в отличие от форм Clupea harengus расстояния между анальным и спинным и брюшными и анальным плавниками, наоборот, увеличиваются. С увеличением размеров головы, как у подсем. Clupeinae, ширина ее увеличивается.
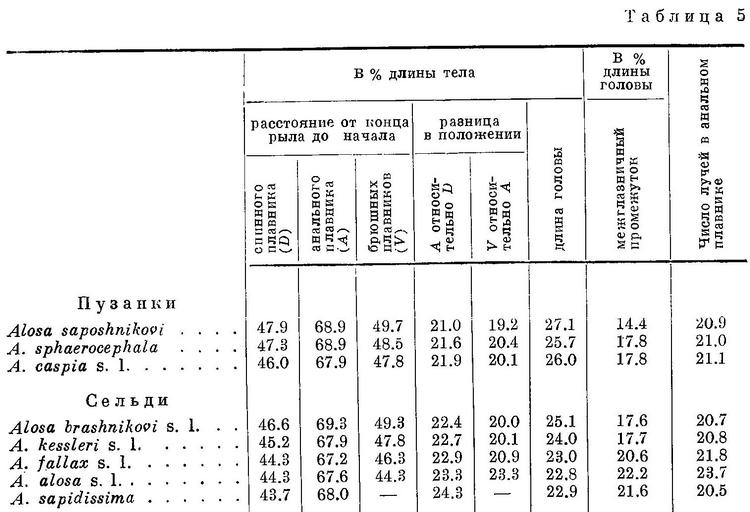
Виды рода Alosa делятся на две группы: на пузанков и сельдей. Виды из группы пузанков (Alosa saposhnikovi, Alosa sphaerocephala и Alosa caspia) по всем рассматриваемым особенностям являются более примитивными, чем виды из группы сельдей, из последних средиземноморско-атлантические виды (Alosa fallax, Alosa alosa, Alosa sapidissima) — наиболее высокоорганизованными. В связи с различием в форме тела пузанков и сельдей в положении спинного и анального плавников относительно переднего конца тела между обеими группами наблюдается некоторое захождение и параллельный характер изменения положения плавников в каждой группе, проявляющийся, как будет указано ниже, и у каспийских и у черноморско-азовских форм Alosa caspia. Однако разность в положении плавников относительно друг друга имеет непрерывный возрастающий характер за исключением разности в положении брюшного плавника относительно анального у Alosa caspia, что, невидимому, обусловлено наличием у него немигрирующих форм, локализованных в южной части моря.
У всех видов рода Alosa за жаберной крышкой сверху с каждой стороны тела имеется по темному пятну. Позади этого пятна у некоторых видов на боках тела есть еще по нескольку таких же пятен, расположенных в ряд. Из группы пузанков у Alosa saposhnikovi имеется только одно пятно за жаберной крышкой; у Alosa sphaerocephala, кроме того, бывает изредка несколько пятен и на боках тела, которые у Alosa caspia встречаются значительно чаще и в большем количестве. Далее, из сельдей у большинства форм Alosa brashnikovi есть только пятно за жаберной крышкой, но у некоторых наиболее специализированных форм этого вида иногда (Alosa brashnikovi nirchi) или обычно (Alosa brashnikovi grimmi) имеется, кроме того, по ряду расплывчатых темных пятен на боках тела. Из форм Alosa kessleri у одних (Alosa kessleri volgensis, мелкая форма Alosa kessleri pontica) имеется только по пятну за жаберной крышкой, у других (Alosa kessleri kessleri, крупная форма Alosa kessleri pontica) изредка есть по ряду из нескольких пятен и на боках тела. Наконец, у Alosa fallax, Alosa alosa и Alosa sapidissima на боках тела есть постоянно по ряду пятен, причем под этим рядом иногда имеется еще 1—2 ряда из меньшего числа пятен. Наличие пятен на боках тела у каспийско-черноморских сельдей ставилось в связь с сезоном, полом, зрелостью половых продуктов. Однако позднее (Дехтярева, 1940 : 61) установлено, что пятна на боках тела есть и у молоди тех видов, у которых они имеются во взрослом состоянии, и, следовательно, наличие их зависит не от указанных причин.
Как и у некоторых видов и подвидов подсем. Clupeinae, существенное значение, наряду с рассматриваемыми особенностями, имеет степень развития зубов. Берг (1913 : 12) на основании того, что у молоди видов Alosa зубы развиты сильнее, чем у взрослых, считает виды с хорошо развитыми зубами примитивными, виды с менее развитыми зубами — более высокоорганизованными. Мнение это стоит в полном согласии с предлагаемой системой рода за немногими лишь исключениями (Alosa kessleri kessleri, черноморско-азовские формы Alosa caspia). Из группы пузанков зубы наиболее сильно развиты у Alosa saposhnikovi, наименее — у всех форм Alosa caspia; Alosa sphaerocephala занимает промежуточное положение между этими видами. Далее, из группы сельдей зубы наиболее развиты у всех форм Alosa brashnikovi. У Alosa kessleri зубы развиты менее, особенно у Alosa kessleri volgensis, иногда почти незаметные; наиболее развиты они у Alosa kessleri kessleri. Наконец, у средиземноморско-атлантических видов Alosa fallax, Alosa alosa и Alosa sapidissima зубы наименее развитые из всех видов рода. У молоди этих видов, как и у упомянутых выше, зубы более развитые (ясно заметные на ощупь), чем у взрослых. Следует отметить, что у молоди зубы наиболее развиты у тех видов (Alosa saposhnikovi, Alosa brashnikovi и др.), у которых они сильнее всего выражены во взрослом состоянии, у молоди же видов со слабыми зубами они менее заметны (Дехтярева, 1940 : 58).
Изменение признаков, положенных в основу системы видов рода Alosa, в обоих группах, пузанков и сельдей, находятся в такой же связи с особенностями их образа жизни, как и у подсем. Clupeinae. Alosa saposhnikovi и Alosa sphaerocephala, у которых спинной и другие плавники наиболее отнесены назад и наименьшая разность в положении плавников относительно друг друга, всю жизнь проводят в море, не заходя в реки. Оба эти вида, судя по всему, совершают миграции в основном из средней части Каспийского моря в северную. Alosa caspia совершает более отдаленные миграции, в особенности его северокаспийские формы, идущие на нерест из южной части моря в северную, причем все черноморско-азовские формы и отчасти Alosa caspia caspia заходят в реки. Из сельдей наименее протяженные миграции совершают формы Alosa brashnikovi, большинство которых всю жизнь проводит в южной части Каспийского моря, и лишь две формы (Alosa brashnikovi brashnikovi, Alosa brashnikovi agrachanica) идут для размножения в северную часть его, не заходя в реки. Черноморско-азовская форма (Alosa brashnikovi maeotica), подобно двум последним, совершает миграции значительной протяженности из Черного моря и северо-западную часть его и в Азовское море. Проходные виды, у которых спинной и другие плавники расположены ближе к переднему концу тела и разность в положении плавников относительно друг друга наибольшая, совершают наиболее отдаленные миграции. Каспийские формы Alosa kessleri (Alosa kessleri volgensis, Alosa kessleri kessleri) из южной части моря идут для икрометания в реки северной части, черноморско-азовская (Alosa kessleri pontica) — из Черного моря в реки северо-западной части его и Азовского моря. Средиземноморско-атлантические виды (Alosa fallax, Alosa alosa, Alosa sapidissima) являются также проходными. Протяженность миграций их в море не известна, но в отношении Alosa sapidissima можно считать установленным, что эти миграции превосходят по протяженности таковые у Alosa kessleri.
Наиболее прогрессивные виды как из пузанков, так и из сельдей имеют и более широкий ареал, отличаясь вместе с тем и большей численностью.
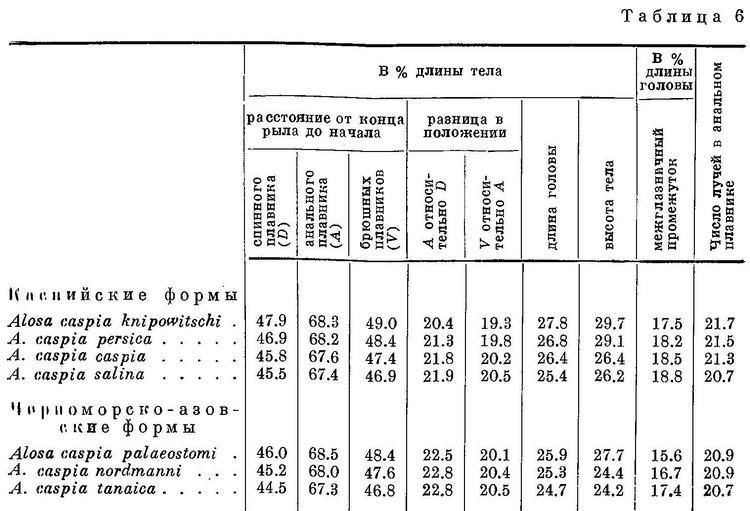
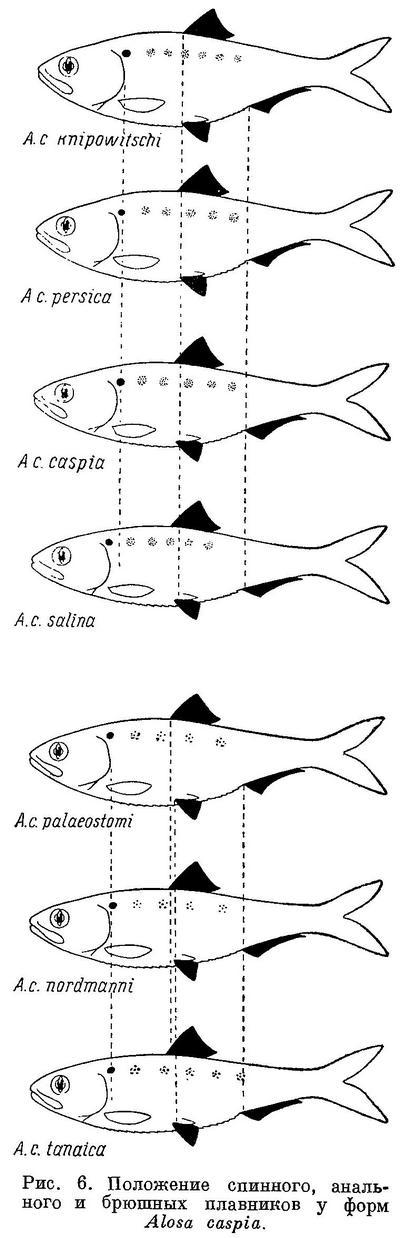
Изменение признаков, положенных в основу классификации, рассматривалось у видов рода Alosa. У подсем. Clupeinae, как указывалось выше, оно наблюдается и у подвидов в пределах видов. То же самое можно, установить у некоторых видов рода Alosa. Так, например, у Alosa caspia такая же зависимость, как и у видов рода, наблюдается в отношении почти всех рассматриваемых признаков, причем, как и у видов из группы пузанков и сельдей Alosa, изменение их, в частности изменение положении плавников, у каспийских и черноморско-азовских форм имеет некоторое захождение и параллельный характер. Однако в каждой из групп изменение положения плавников имеет характер закономерного перемещения их к передней части тела (табл. 6, рис. 6). При этом все признаки изменяются сходным образом, как и у видов рода. В сходной взаимосвязи находятся рассматриваемые признаки и с образом жизни форм Alosa caspia. Среди каспийских форм спинной и анальный плавники расположены далеко назад, разность в положении их относительно друг друга меньше и лучей в анальном плавнике больше у форм, живущих о размножающихся в южной части моря (Alosa caspia knipowitschi, Alosa caspia persica). У форм, зимующих в южной части моря, а для размножения идущих в северную часть моря (Alosa caspia caspia, Alosa caspia salina), спинной и анальный плавники расположены ближе к переднему концу тела, разность в положении их относительно друг друга больше и лучей в анальном плавнике меньше. Черноморско-азовские формы Alosa caspia соответственно в общем более близкому расположению плавников к переднему концу тела и пр., чем у каспийских форм, ведут в общем и более подвижный образ жизни, причем все они для размножения заходят в низовья рек, в озера и лиманы, и ни одна из них не мечет икру в море, между тем как большинство каспийских форм размножается в море, в заливах и ильменях или в устьях рек. Наименьшие по протяженности миграции из всех черноморско-азовских форм Alosa caspia совершает Alosa caspia palaeostomi, локализованный в южной части моря, наибольшие — Alosa caspia nordmanni и Alosa caspia tanaica; первые с мест зимовок от южных берегов Крыма и западных Черного моря идет в реки северо-западной части его, второй от южного кавказского побережья — в р. Дон и лиманы дельты Кубани. Помимо указанных признаков как у каспийских, так и у черноморско-азовских форм Alosa caspia сходным образом изменяется, высота тела, находящаяся также в зависимости от миграционных особенностей. Как видно из рис. 6 и табл. 6, у форм, совершающих миграции большей протяженности в северные части моря, тело значительно ниже, в особенности у каспийских, чем у форм, локализованных в южных частях моря. Что касается темных пятой на боках тела, то у черноморско-азовских форм они встречаются реже и в менее выраженной форме, чем у каспийских. Следует отметить, что у черноморско-азовских форм и других видов рода Alosa (Alosa kessleri pontica) пятна на боках тела развиты менее.
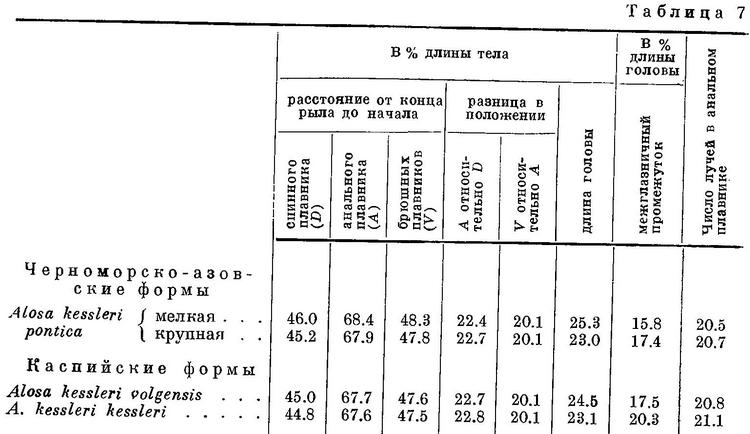
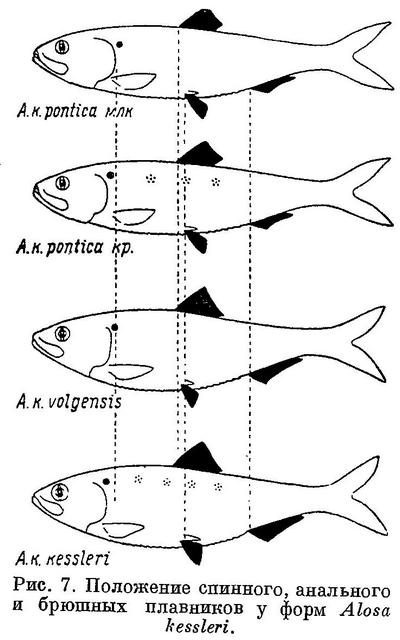
У Alosa kessleri рассматриваемые признаки изменяются непрерывно на протяжении всех форм вида (табл. 7, рис. 7), за исключением длины головы, изменение которой, как и у форм Alosa caspia, имеет параллельный характер соответственно делению форм вида на черноморско-азовских и каспийских (табл. 7, рис. 7). Что касается степени развития зубов, то они менее всего развиты у Alosa kessleri volgensis, более — у Alosa kessleri pontica и Alosa kessleri kessleri. Таким образом, степень развития зубов в данном случае не стоит в связи с изменением других признаков, что вполне понятно, так как эта особенность связана не с движением, а с питанием.
Судя по всем рассматриваемым признакам, за исключением степени развития зубов, наиболее высокоорганизованной формой вида следует считать Alosa kessleri kessleri. Это стоит в полном согласии и с тем, что пятна на боках тела из обеих каспийских форм имеет только эта. По более отнесенным назад плавникам, большим размерам и большей ширине головы и пр. наиболее примитивной формой Alosa kessleri следует считать Alosa kessleri pontica. При этом крупная форма Alosa kessleri pontica, имеющая изредка на боках тела темные пятна, соответствует биологически и отчасти морфологически Alosa kessleri kessleri, у которой пятна на теле бывают чаще, а мелкая форма — Alosa kessleri volgensis. Более высокой степени развития рассматриваемых признаков у Alosa kessleri kessleri по сравнению с Alosa kessleri volgensis и крупной формы Alosa kessleri pontica по сравнению с мелкой соответствуют особенности в распространении и биологии, подобные отмеченным у более высокоорганизованных видов рода Alosa и форм Alosa caspia. Alosa kessleri kessleri и крупная форма Alosa kessleri pontica заходят для нереста высоко вверх по течению рек, в то время как мелкая форма Alosa kessleri pontica и Alosa kessleri volgensis высоко в реки не поднимаются и нерестуют в нижнем течении, последняя иногда частично, даже в предустьевом пространстве Волги.
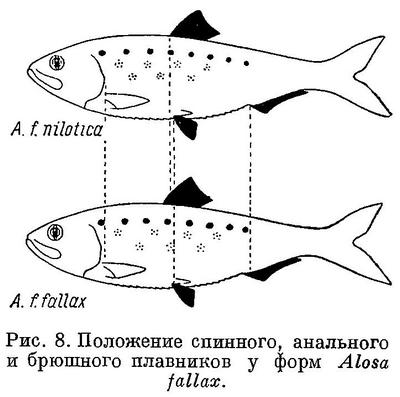
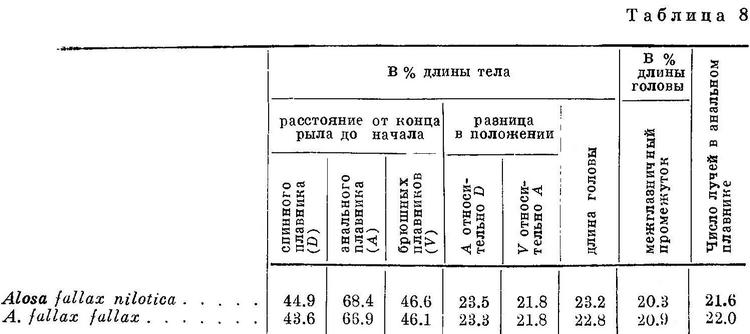
Из всех форм Alosa fallax и Alosa alosa представилась возможность исследовать лишь некоторые и притом на немногочисленном материале. Исследуемые признаки и этих форм обоих видов имеют в общем такие же закономерные изменения, как и у других видов рода (табл. 8, рис. 8).
Alosa brashnikovi, наиболее богатый формами из всех видов рода, обладающий, кроме того, некоторыми своеобразными биологическими адаптациями, крайне изменчив в основных, связанных с этими адаптациями морфологических особенностях. Формы этого вида, расположенные соответственно положению спинного плавника, почти не обнаруживают какой-либо закономерности в изменении других рассматриваемых признаков, почему фактический материал здесь не приводится. Однако следует отметить, что и здесь у форм, идущих для нереста в северную часть Каспийского моря (Alosa brashnikovi brashnikovi и Alosa brashnikovi agrachanica), спинной и анальный плавники расположены ближе к переднему концу тела, межглазничный промежуток шире, чем у других форм. Вместе с тем, у Alosa brashnikovi kisselevitshi, ведущей наиболее пелагический образ жизни, но живущей и размножающейся в южной части Каспийского моря, спинной плавник расположен еще ближе к переднему концу тела, а межглазничный промежуток так же широк, как у обеих упомянутых форм. Наиболее отнесены плавники назад и уже голова у южнокаспийских форм (Alosa brashnikovi orientalis, Alosa brashnikovi grimmi, Alosa brashnikovi nirchi), ведущих придонный образ жизни.
Морфологические особенности форм Alosa brashnikovi обусловлены приспособлениями их к различным условиям существования. Основным фактором образования форм этого вида является, повидимому, приспособление их главным образом к различному характеру питания, крайне разнообразному у форм этого вида, вследствие чего у них и наблюдаются большое разнообразие и различия в строении жаберных тычинок, рта и частей черепа, связанных с питанием. Наиболее резко обособленная форма вида, Alosa brashnikovi grimmi, является вместе с тем по образу жизни и характеру питания наиболее придонной из всех форм вида. Питается эта форма преимущественно бычками, иногда даже моллюсками (Cardium), реже тюлькой, в связи с чем жаберных тычинок у ней наименьшее количество, а на конце нижней челюсти у взрослых рыб имеется мозолистое утолщение, развивающееся, повидимому, вследствие соприкосновения нижней челюсти с грунтом при захватывании пищи. Весьма характерно, что у молодых рыб, длиной до 20 см, мозолистого утолщения на нижней челюсти нет. Повидимому, по этой же причине отсутствует у Alosa brashnikovi grimmi и бугорок на конце нижней челюсти, имеющийся почти постоянно у всех других форм вида. Вследствие механического воздействия грубой пищи жаберные тычинки искривляются, на конце утолщаются, расплющиваются, разветвляются и обламываются, а тычинки на нижнем конце жаберной дуги редуцируются и отсутствуют даже у молодых, благодаря чему первая тычинка у этой формы отстоит далеко от основания языка. Окраска у Alosa brashnikovi grimmi белесая, а на боках тела с каждой стороны, кроме пятна за жаберной крышкой, обычно имеется по ряду расплывчатых серых пятен, почти сливающихся в продольную полосу. Близкая к Alosa brashnikovi grimmi и ведущая подобно ей придонный образ жизни Alosa brashnikovi nirchi имеет иногда на нижней челюсти точно такое же, но небольшое мозолистое утолщение; окраска у этой формы бледная; на боках тела, помимо пятна за жаберной крышкой, иногда имеется по ряду расплывчатых серых пятен. Наиболее резко уклоняющаяся от этих форм, а в особенности от Alosa brashnikovi grimmi, Alosa brashnikovi kisselevitshi имеет самое большое количество жаберных тычинок из всех форм вида, причем тычинки у этой формы обычно заостренные, не утолщенные и не разветвленные на концах, первая тычинка расположена близко от основания языка. Держится эта форма в поверхностных слоях воды, питается тюлькой, креветками, бычками. Вследствие различий в характере питания наблюдаются изменения и в строении частей скелета головы Alosa brashnikovi grimmi. Часть черепа у этой формы в области прикрепления hyomandibulare, а посредством него и нижней челюсти (именно передний конец каждого sphenoticum) укорочена и сзади закруглена и не изогнута крючкообразно, как у других форм вида (например у Alosa brashnikovi brashnikovi, Alosa brashnikovi orientalis, Alosa brashnikovi kisselevitshi). Сходное строение имеет передний конец каждого sphenoticum и у Alosa brashnikovi nirchi. У этих форм наблюдаются существенные различия и в строении жаберных тычинок, подробно описанные в диагнозах.
Таким образом, из этих примеров видно, что наиболее специализированными формами Alosa brashnikovi следует считать Alosa brashnikovi grimmi и затем Alosa brashnikovi nirchi, приспособившихся к совершенно несвойственному сельдевым придонному образу жизни и преимущественно бентоядному питанию. Эта биологическая специализация сопровождается рядом глубоких морфологических изменений и появлением пятен на боках тела, свойственных, как указывалось выше, наиболее высокоорганизованным видам и формам. Спинные же и другие плавники расположены ближе к переднему концу тела, и разница в положении их относительно друг друга больше у форм, идущих для размножения в северную часть Каспийского моря (Alosa brashnikovi brashnikovi и Alosa brashnikovi agrachanica), чем у форм, постоянно живущих и размножающихся в южной части моря (Alosa brashnikovi sarensis, Alosa brashnikovi autumnalis), а среди последних менее всего у ведущих придонный образ жизни (Alosa brashnikovi grimmi, Alosa brashnikovi nirchi). Среди южнокаспийских форм Alosa brashnikovi спинной и анальный плавники расположены ближе всего к переднему концу тела и разность в положении этих плавников относительно, друг друга больше у наиболее пелагической формы (Alosa brashnikovi kisselevitshi).
Естественно, что наиболее прогрессивные по рассматриваемым морфологическим особенностям формы Alosa caspia и Alosa brashnikovi, идущие для икрометания в северную часть Каспийского моря, занимают и более широкий ареал, чем формы, локализованные в южной части моря. Вместе с тем эти формы характерны и большей численностью.
При сопоставлении состава фауны сельдевых Каспийского, Черного и Азовского морей обращают на себя внимание несколько особенностей. Одна из этих особенностей заключается в том, что фауна рыб Черного моря, как известно, значительно более богатая и разнообразная по своему составу, чем Каспийского, более богата и разнообразна и в отношении сельдевых, но не видами и более низкими формами (подвидами), а родами. Здесь, помимо общих с каспийскими родом Clupeonella и подродом Caspialosa, имеются еще 3 рода из этого семейства — Sprattus, Sardina, Sardinella и подрод Alosa s. str., широко распространенных, кроме Черного моря, в Средиземном море и в Атлантическом океане, а некоторые из них, кроме того, в Индийском и Тихом океанах. Эта особенность черноморской фауны сельдевых, свойственная, как отмечалось выше, вообще фауне Черного моря, обусловлена, как известно, инвазией в Черное море средиземноморской фауны после прорыва Босфора.
Еще больший интерес представляет другая особенность черноморских и каспийских сельдевых, не отмеченная до сего времени и заключающаяся в том, что общие обоим морям род Clupeonella и подрод Caspialosa в Каспийском море как в видовом отношении, так и по количеству более низких форм (подвидов) представлены значительно более богато, чем в Черном и Азовском морях. Эта особенность была отмечена автором ранее в предварительном сообщении (Световидов, 1941 : 807, предст. 13 VI 1940). Эта статья должна была быть напечатана одновременно со статьей А. А. Михайловской (1941 : 559, предст. 24 VI 1940), которая была знакома с выводами автора и, говоря о богатстве каспийских сельдей, сделала ссылку на нее. Вследствие того, что статья автора была опубликована позднее: статьи А. А. Михайловской, эта ссылка в ее статье помещена не была.
Состав фауны сельдевых этих родов в Каспийском и Черном и Азовском морях таков (В скобках указано количество подвидов.):

1 Четвертый вид этого рода Clupeonella abrau встречается в оз. Абрау и Абулионд; о нем сказано особо ниже.
Род Clupeonella в Каспийском море представлен 2 эндемичными видами и лишь одним — формой того же вида, что и в Каспийском, — в Черном и Азовском морях. В Каспийском море из рода Alosa имеются 6 видов, из которых 3 свойственны только этому морю; в Черном же и Азовском морях из этого рода встречается только 3 вида, представленные здесь формами видов, общих с каспийскими. Таким образом, в Черном и Азовском морях нет ни одного вида, который не был бы представлен в Каспийском и притом значительно большим количеством форм, чем в Черном и Азовском морях, в то время как в Каспийском море имеются 2 вида из рода Clupeonella и 3 вида из рода Alosa, отсутствующие в Черном и Азовском морях.
Богатство сельдевых обоих этих родов в Каспийском море обусловлено преобладанием главным образом морских, непроходных видов и подвидов. Таковыми из числа видов являются Clupeonella grimmi и Clupeonella engrauliformis и Alosa saposhnikovi, Alosa sphaerocephala и Alosa curensis, в то время как проходная (Alosa kessleri s. l.) и полупроходная (Alosa caspia s. l.) сельди и тюлька (Clupeonella delicatula s. l.) представлены в обоих морях. Из числа морских, непроходных сельдей общими для обоих морей являются лишь Alosa brashnikovi s. l. Подвидами в Каспийском море особенно богат морской, непроходной вид Alosa brashnikovi s. l., представленный здесь 8 подвидами, против одного черноморско-азовского (Alosa brashnikovi maeotica). Полупроходные и проходные виды имеют в Каспийском море значительно меньшее количество подвидов. Так, например, Alosa caspia, одни подвиды которого являются морскими, непроходными (Alosa caspia salina), другие полупроходными или отчасти даже проходными (Alosa caspia caspia и др.), представлен в Каспийском море 4 подвидами и, повидимому, еще
2 более низкими формами (nationes), а в Черном и Азовском морях — 3 подвидами. Типично проходной вид Alosa kessleri s. l. имеет в Каспийском море 3 подвида, а в Черном и Азовском лишь один, распадающийся на 2 более низких формы (nationes).
Необходимо также отметить некоторые биологические особенности каспийских и черноморско-азовских форм Alosa brashnikovi s. l. и Alosa caspia s. l., а именно то, что каспийские формы этих видов являются более морскими рыбами, чем черноморско-азовские. Все каспийские формы Alosa brashnikovi s. l. избегают пресной воды и размножаются только в море, между тем как черноморско-азовская форма этого вида (Alosa brashnikovi maeotica) заходит в совершенно опресненные участки Таганрогского зал., и в нижнюю часть дельты Дона. Имеются указания и на нерест Alosa brashnikovi maeotica почти в пресной воде. Среди каспийских форм Alosa caspia s. l. есть такие, которые мечут икру в солоноватой воде в море (Alosa caspia salina) или в предустьевых пространствах (Alosa caspia caspia), в то время как все черноморско-азовские формы этого вида мечут икру только в пресной воде, заходя в реки, озера и лиманы.
Заслуживает внимания и еще одна особенность каспийских форм обоих рассматриваемых родов, отмеченная ранее Киселевичем (1918 : 21), — их значительно большие размеры, по сравнению с черноморско-азовскими. Clupeonella delicatula caspia достигает в длину до 14 см, средние размеры ее около 8—10 см, между тем как размеры Clupeonella delicatula delicatula в Азовском море не превышает 9 см, имея в среднем около 7 см. Каспийские формы Alosa caspia s. l. точно так же значительно крупнее черноморско-азовских. Alosa caspia caspia иногда достигает в длину до 28 см, средние размеры его около 18—21 см; таковы же размеры и остальных каспийских форм этого вида, в то время как черноморско-азовские формы не превышают в длину 20 см, средние размеры их около 15 см; каспийские формы Alosa brashnikovi s. l. достигают в длину почти до 50 см, средние размеры их около 30 см; наибольшие размеры черноморско-азовскои формы этого вида (Alosa brashnikovi maeotica) не превышают 31 см, средние размеры крупной расы этой формы — от 16 до 20 см, мелкой расы — около 12—13 см. Наиболее крупная из каспийских форм Alosa kessleri s. l., Alosa kessleri kessleri, достигает в длину до 52 см, в половозрелом состоянии во время хода в реку основная масса ее имеет длину от 36 до 44 см. Более мелкая форма этого вида, Alosa kessleri volgensis, достигает в длину до 40 см, средние размеры ее во время хода в реку около 28 см, в то время как наибольшие размеры черноморско-азовской формы (Alosa kessleri pontica) в Дону не превышают 30, а в Днепре 39 см.
Наиболее важная особенность сельдевых обоих рассматриваемых родов заключается, однако, в том, что каспийские формы их различаются значительно больше, чем черноморско-азовские формы тех же видов. Различия между каспийскими формами Alosa caspia s. l. значительны как в количестве жаберных тычинок, так в некоторых случаях и внешне (например Alosa caspia knipowitschi). Черноморско-азовские формы Alosa caspia s. l., не различающиеся между собой внешне, значительно меньше, чем каспийские формы этого вида, отличаются друг от друга и количеством жаберных тычинок. Разница в средних значениях количества жаберных тычинок у наиболее различающихся по этому признаку черноморско-азовских форм Alosa caspia s. l., Alosa caspia tanaica и Alosa caspia nordmanni, равна только 4, а у менее различающихся Alosa caspia tanaica и Alosa caspia palaeostomi — всего лишь 0.1, между тем как у наиболее различающихся в этом отношении каспийских форм этого вида Alosa caspia salina и Alosa caspia knipowitschi разница в средних значениях количеств жаберных тычинок равна 55.5. Даже у наименее отличающихся по количеству жаберных тычинок каспийских форм этого вида Alosa caspia salina и Alosa caspia persica разница в средних значениях равна 7.8. Такова же степень различия между каспийскими формами Alosa caspia s. l. с одной стороны, и черноморско-азовскими, с другой, и в остальных признаках. Черноморско-азовская проходная сельдь (Alosa kessleri pontica), как показали исследования последних лет, распадается на 2 формы, существенно различающиеся биологически, но морфологически ни количеством и строением жаберных тычинок, ни какими-либо другими особенностями, кроме размеров, не отличающиеся и являющиеся более низкими таксономически формами (nationes). Обе каспийские проходные сельди (Alosa kessleri kessleri и Alosa kessleri volgensis) существенно отличаются друг от друга количеством и строением жаберных тычинок, некоторыми другими особенностями и отчасти даже внешне. Еще более различаются между собой каспийские формы Alosa brashnikovi s. l., представленного в Черном и Азовском морях одной формой, которая, подобно Alosa kessleri pontica, повидимому, распадается на 2 более низкие, чем подвиды, формы (nationes), различающиеся лишь размерами и биологическими особенностями.
Менее различаются между собой в Черном море и виды Alosa brashnikovi s. l. и Alosa kessleri s. l. Наиболее близкая к Alosa brashnikovi s. l. по количеству жаберных тычинок форма Alosa kessleri s. l., а именно Alosa kessleri kessleri, имеет не менее 59 тычинок, в то время как у Alosa brashnikovi kisselevitshi, имеющей наибольшее количество жаберных тычинок из всех каспийских форм этого вида, количество их не превышает 49. Что касается черноморско-азовских форм Alosa brashnikovi s. l. и Alosa kessleri s. l., то у Alosa brashnikovi maeotica наибольшее количество жаберных тычинок 46, а наименьшее количество их у Alosa kessleri pontica 47.
Богатство каспийских сельдевых родов Clupeonella и Alosa как в видовом отношении, так и количеством форм общих видов, их гигантизм по сравнению с черноморско-азовскими формами, отмеченные выше биологические особенности и большая степень отличий обусловлены различиями в условиях развития и формообразования сельдевых как в Каспийском и Черном и Азовском морях, так и в тех морях, которые предшествовали им. Поэтому для рассмотрения условий развития и формообразования каспийских и черноморско-азовских сельдевых обоих этих родов и выяснения причин такого резкого различия в составе их, размерах, биологических особенностях и степени отличий необходимо обратиться к истории развития этих морей, так как история развития фауны, по вполне справедливому замечанию Колесникова (1940 : 6), не должна быть оторвана от истории развития бассейна, в котором она обитает.
Понто-каспийский морской бассейн, как известно, претерпел сложную геологическую историю. К концу миоцена, при сокращении Тетиса, одновременно с похолоданием климата, северо-восточная часть его отделилась, образовался сарматский и ряд других пришедших ему на смену бассейнов, фауна которых в дальнейшем развивалась в значительной степени изолированно, находясь, в отличие от фауны образовавшегося к этому времени Атлантического океана и стоявшего в связи с ним Средиземного моря, под сильным воздействием периодических опреснений и осолонений. В меньшей степени опреснения и бассейны с затрудненным сообщением с нормально соленым морским бассейном или даже замкнутые бассейны в северо-восточной части Тетиса были и раньше, в нижнем и среднем миоцене (Жижченко, 1940 : 42, 148).
Биологические взаимоотношения видов Clupeonella и Alosa с другими рыбами как в настоящее время в Каспийском море и Черном и Азовском морях, так несомненно и в морях, предшествовавших им в предыдущие эпохи, различны. Пелагическая фауна открытых частей Каспийского моря состоит исключительно из сельдевых, которые (все виды Clupeonella и формы Alosa caspia s. l.) являются здесь основными потребителями планктона (см. об этом Бенинг, 1938 : 33). Других пелагических рыб, кроме сельдевых, в галистатической области Каспийского моря в сущности нет. Население пелагиали более открытых частей Черного моря составляют в основном средиземноморские иммигранты как из сельдевых, но других родов. Что касается сельдевых из родов Clupeonella и Alosa, то они вместе с остальной автохтонной реликтовой фауной понтического озера-моря встречаются в опресненных частях Черного моря и в Азовском море. Тюлька (Clupeonella delicatula delicatula) встречается здесь только в Азовском море, в лиманах и в опресненных участках у берегов северо-западной части Черного моря, Румынии, Болгарии, в Феодосийском зал. и в оз. Палиастоми. Сельди в зимнее время держатся вдоль берегов Кавказа, у берегов Крыма, летом заходят в опресненные части Черного моря и в Азовское море и в реки.
В галистатической части Черного моря ни один из представителей этих двух родов не найден.
Таким образом, виды рода Clupeonella, живущие только в море и притом в открытых его частях (Clupeonella grimmi и Clupeonella engrauliformis), и морские, непроходные виды рода Alosa (Alosa saposhnikovi, Alosa sphaerocephala, Alosa brashnikovi s. l.), мечущие икру в море, в Черном море не могли образоваться или получить развитие потому, что пелагиаль открытых частей его после прорыва Босфора была заселена более сильными конкурентами, иммигрантами из Средиземного моря. Наоборот, можно полагать, что сельдевые из этих двух родов в настоящую фазу существования Черного моря испытывают в нем сильную депрессию, будучи оттеснены в наиболее опресненные части его и в Азовское море более жизненными средиземноморскими иммигрантами.
Совершенно иные условия оказались в Каспийском море, которое в средней и, в особенности, в южной части в настоящую фазу его существования представляет солоноватоводный, в южной части почти такой же морской бассейн, как Черное море. В этом бассейне пелагических рыб кроме сельдевых из родов Clupeonella и Alosa не оказалось, поэтому фауна пелагиали его, в особенности в южной и средней частях, могла формироваться только из этих рыб, и поэтому же они, в особенности морские непроходные, при отсутствии конкуренции с другими видами и получили здесь пышный расцвет; расцвет этот сопровождается и гигантизмом; аналогичное явление наблюдалось и в понтической реликтовой фауне, например в киммерийском бассейне (Колесников, 1940 : 930). Расцвет в Каспийском море одного из наиболее богатого здесь формами видов, Alosa brashnikovi s. l., в особенности в южной части моря, где локализовано большинство форм его, обусловлен не только отсутствием здесь других пелагических рыб, кроме сельдевых, но также приспособлением некоторых форм его и к придонному образу жизни и питанию. При этом некоторые формы Alosa brashnikovi s. l., помимо существенных различий в образе жизни, характере питания и в связанных с этим некоторых морфологических особенностях, о чем подробно указывалось выше, отличаются большим разнообразием и во времени нереста, имея в этом отношении некоторое сходство с атлантической сельдью (Clupea harengus harengus). Среди форм этого вида имеются как наиболее холодолюбивые (Alosa brashnikovi brashnikovi), так и самые теплолюбивые (Alosa brashnikovi kisselevitshi) из всех каспийских сельдей, причем по окончании нереста одной формы (например Alosa brashnikovi grimmi, в июне и даже в июле) на тех же местах позднее начинают метать икру другие (Alosa brashnikovi kisselevitshi).
Пелагиаль открытых частей Средиземного моря и Атлантического океана, благодаря постоянству состава их ихтиофауны с конца миоцена, была всегда заселена, помимо упомянутых для Черного моря, рядом других пелагических рыб. В частности, в пелагиали бореальной области Атлантического океана наиболее существенную роль играет атлантическая сельдь (Clupea harengus harengus). Поэтому средиземноморско-атлантические сельди из Alosa, не имея подходящих условий для расцвета в прошлом, не имеют и, вероятно, не имели и ранее ни одного вида, который размножался бы в море или в солоноватой воде; все они являются проходными видами. Более того, многие из них образовали чисто пресноводные, речные или озерные формы, которых каспийско-черноморские сельди не имеют. Таковыми являются: Alosa fallax lacustris (озера Маджоре, Комо, Лугано), Alosa fallax gracilis (оз. Гарда), Alosa fallax killarnensis (оз. Килларней в юго-западной части Ирландии), Alosa alosa macedonica (оз. Бешик-гёль, бассейн Эгейского моря), Alosa alabamae ohiensis (р. Огайо, бассейн Миссисипи).
Средиземноморско-атлантические сельди Alosa alosa и Alosa fallax морфологически и биологически соответствуют каспийским проходным сельдям Alosa kessleri kessleri и Alosa kessleri volgensis. Alosa alosa alosa и Alosa kessleri kessleri — крупные и жирные сельди, заходящие в реки с еще не вполне зрелыми половыми продуктами и подымающиеся для нереста высоко вверх по течению. Alosa fallax fallax и Alosa kessleri volgensis — менее крупные и жирные сельди, в реки заходят с вполне зрелыми половыми продуктами и высоко вверх не подымаются (Alosa fallax fallax в Рейне мечет икру в пределах Голландии, Alosa kessleri volgensis — в Волге в главной массе ниже Сталинграда, в то время как обе первые сельди нерестятся значительно выше). Черноморско-азовская проходная сельдь Alosa kessleri pontica, как указывалось выше, распадается на 2 формы (nationes) — крупную и мелкую. Крупная форма в реки входит раньше и с менее зрелыми половыми продуктами, чем мелкая, она более жирна. В настоящее время вследствие возведения плотин на Днепре и на Дону трудно сказать, подымается ли крупная форма Alosa kessleri pontica вверх по течению выше, чем мелкая, но в прежние годы в Днепре на порогах ловилась только крупная сельдь.
Как нетрудно видеть, крупная форма Alosa kessleri pontica биологически соответствует Alosa kessleri kessleri и Alosa alosa alosa, а мелкая — Alosa kessleri volgensis и Alosa fallax fallax, и все эти проходные сельди в трех рассматриваемых морских бассейнах представляют 3 формы разной степени обособления. Благодаря тому что в Атлантическом океане и в Средиземном море с конца миоцена не было таких изменений солености и сопутствующих им изменений в составе ихтиофауны, как в бассейнах, давших начало Черному и Каспийскому морям, в конечном итоге, в силу изменчивости в одном направлении, при которой не появлялись формы, имевшие признаки, отчасти свойственные предшествовавшим формам, отличия между Alosa alosa и Alosa fallax оказались более значительными и они обособились здесь до степени видов, имеющих и качественные различия. Каспийско-черноморские проходные сельди, замкнутые в сарматском и других сменивших его бассейнах, вследствие ряда последовательных опреснений и осолонений в этих морях, вызывавших, как указывалось выше, появление форм, имевших некоторые признаки, отчасти свойственные предыдущим формам, обособились менее. Каспийские сельди, являющиеся в настоящую фазу существования Каспийского моря единственными пелагическими рыбами, обособились до степени подвидов, имеющих лишь количественные отличия, в то время как черноморско-азовские проходные сельди, оттесненные средиземноморскими иммигрантами в опресненные части Черного моря и в Азовское море и находящиеся в менее благоприятных условиях, обособились менее, до степени менее обособленных форм (nationes). Большей степенью различий, чем черноморско-азовские, отличаются, как указывалось выше, и другие каспийские формы общих с черноморско-азовскими формами видов (Alosa caspia s. l., Alosa brashnikovi s. l.). Разная степень различия между видами подрода Alosa s. str. и формами каспийских и черноморско-азовских проходных сельдей подрода Caspialosa наблюдаются, между прочим, и в количестве лучей в анальном плавнике. Количество лучей в анальном плавнике у всех, даже у видов Alosa, трансгрессивно, но разница в средних у видов Alosa значительно больше, чем у подвидов и более низких форм (nationes) Alosa, что видно из нижеследующего: Alosa alosa alosa — 23.1, Alosa fallax fallax — 22.0, Alosa kessleri kessleri — 21.1, Alosa kessleri volgensis — 20.8, Alosa kessleri pontica крупная форма — 20.7, Alosa kessleri pontica мелкая форма — 20.5.
Подводя итоги всему сказанному об условиях развития и формообразования сельдевых из рода Clupeonella и Alosa, следует подчеркнуть, что богатство их в Каспийском море обусловлено расцветом их в этом море главным образом в настоящую фазу его существования, со времени последнего соединения его с морями, предшествовавшими Черному и Азовскому, которое могло быть в послехвалынское и в бакинское, а ранее в апшеронское и акчагыльское время (см. об этом: Колесников, 1940 : 928), а не в одном из морей, предшествовавших Каспийскому, как полагал Киселевич (1918 : 24). При этом расцвет их обусловили не «значительное протяжение и разнообразие климатических условий в различных частях бассейна» (Киселевич, 1918 : 24), а наличие в Каспийском море незаселенной другими рыбами пелагиали, которую могли заполнить только сельдевые рассматриваемых двух родов. Что касается приводимого К. А. Киселевичем мнения Сарса, что каспийские сельди «находятся в стадии незакончивщегося видообразования» (1918 : 24), то в стадии видообразования находится весь органический мир, даже наиболее древние его представители.
Рыбы пресных вод СССР и сопредельных стран. Л.С.Берг 1948 г.
Род ALOSA CUVIER
Alosa Cuvier, Regne animal, ed. 2, II, 1829, p. 319 (тип: Alosa alosa).
Alausa Valenciennes in Сuv. - Va1., XX, 1847, p. 389 (тип: Alosa vulgaris = Alosa alosa).
Alosa C. Tate Regan, Ann. Mag. Nat. Hist. (8), XVIII, 1916, p. 6 (обзор видов).
Paralosa Roule, Les poissons des eaux douces de la France, 1925, p. 73 (тип: Alosa fallax Lac. = Alosa finta), non Paralosa Regan 1916.
Как Caspialosa, но зубов на сошнике никогда не бывает; очень мелкие зубы бывают только на межчелюстной и верхнечелюстной костях. Поперечных рядов чешуй 55—86. Позвонков 52—59. Пилорических придатков 100.
5 видов в Европе, в Ниле, Алжире и в атлантических частях Сев. Америки, Мексиканский залив, Миссисипи. Входят из моря в реки.
1(2). Жаберных тычинок на первой дуге менее 50. 1. Alosa fallax (Lac.)
2(1). Жаберных тычинок на первой дуге не менее 90. 2. Alosa alosa (L.)
Род CASPIALOSA BERG. — КАСПИЙСКО-ЧЕРНОМОРСКИЕ СЕЛЬДИ
Clupeonella Берг (non Kessler), Мат. к позн. русск. рыбол., II, вып. 3, 1913, стр. 10.
Caspialosa Берг, там же, IV, вып. 6, 1915, стр. 4 (тип: Caspialosa caspia).
Брюхо сжатое с боков, снабженное на всем протяжении от горла до начала анального плавника шипообразными чешуйками; эти брюшные шипы сильные, ясно заметные, свободные (не лежат в бороздках), числом 29—36. Начало спинного плавника ближе к концу рыла, чем к основанию хвостового плавника, в нем II—V 11—16 лучей. Основание спинного плавника лежит в бороздке, образованной расширенными чешуями. Последний луч D не вытянут. Брюшные плавники с I 8—9 лучами, начало их под началом D или чаще несколько позади: под передней третью D. Анальный плавник с II—IV 15—20 лучами. Последние два луча анального плавника не вытянуты. При основании хвостового плавника с каждой стороны по удлиненной чешуе (ala). Зубы на сошнике всегда есть (у взрослых Caspialosa caspia слабо развитые), обыкновенно есть и на челюстях, нёбных костях и на языке. На межчелюстной кости заметная вырезка. Рот конечный, большой: сочленение нижней челюсти с черепом заходит за вертикаль заднего края глаза. Верхняя челюсть доходит или почти доходит до вертикали заднего края глаза. На глазах хорошо развитые веки. На крышечной кости ясные радиальные полоски. Жаберных тычинок 18—168. Ширина свободной части щек меньше их высоты. Чешуя легко опадающая, расположенная в 49—60 поперечных рядов. Пилорических придатков 18—60. Позвонков 43—55. Яичники желтого цвета. Икра донная, полупелагическая; икринки без жировой капли, с большим околожелтковым пространством (Относительно икринок см:. Т. С. Расc, Доклады Акад. Наук 1936, II, стр. 301, фиг. 3.). Длина 160—520 мм.
Около 12 видов. Солоноватоводные и проходные рыбы Каспийского и Черного морей. Moreau (Hist. nat. des poissons de la France. Supplement. Paris, 1891, p. 75—78) упоминает об одном экземпляре «Clupea pontica» длиною 103 мм, добытом в 1885 г. в южной Франции, у Сетта (Cette, etang de Thau). Описание не оставляет сомнения в том, что оно относится к представителю рода Caspialosa, но я думаю, что оно составлено не по экземпляру в 103 мм из Сетта, а по экземпляру из Константинополя длиной в 233 мм, который автор имел для сравнения. Сомнительно, чтобы молодой экземпляр Caspialosa мог дойти до берегов Франции. Предполагать же, чтобы Caspialosa размножалась у берегов Франции, нет оснований. Не имел ли в руках Moreau молодой экземпляр Alosa fallax (finta)?
Метод определения каспийских сельдей по комбинации признаков был разработан А.В.Морозовым. Из признаков, имеющих значение для характеристики видов, рода Caspialosa, нужно указать на число позвонков. (См.: Г. П. Булгаков, Бюлл. Ср.-аз. унив., № 16, 1927, стр. 123—133. См. также Аверинцев и Сыч, 1930.) Замечу еще, что характер ветвления чувствительных каналов на infraorbitalia, praeoperculum, operculum, на верху головы, на затылочных и плечевых чешуях, обнаруживая, в основном строении, сходство у разных видов этого рода, в деталях показывает известные отличия.
Д. Ф. Замахаев обнаружил на чешуе как каспийских, так и черноморских проходных видов рода Caspialosa нерестовые знаки. (Д. Ф. Замахаев, Нерестовые марки на чешуе каспийских сельдей. Труды Инст. мор. рыбн. хоз., XIV, 1940, стр. 3—19.)
Об икре и личинках каспийских сельдей см. в статьях Т. А. Перцевой, Рыбн;. хоз., 1938, № 7, стр. 28—32; Тр. Инст. мор. рыбн. хоз., XIV, 1940, стр. 109 (пузанок); И. И. Казановой и Н. А. Халдиновой, там же, стр. 77; С. Г, Сомовой, там же, стр. 149 (черноспинка); о сеголетках см. А. И. Дехтерева, там же, стр. 47 (определитель).
Выяснено, что из северного Каспия все сельди — как проходные, так и непроходные — на холодное время года, как правило, уходят в среднюю и южную части моря. Отсюда они весной возвращаются на север для нереста, после чего главная масса сельдей еще летом уходит на юг; лишь немногие остаются на севере до осени или до заморозков, а ильменный пузанок, по данным М. А. Идельсона, может даже перезимовывать в ильменях, подо льдом. Но это исключение. Кроме взрослых рыб, на юг уходит и молодь. Словом, эти сельди ведут себя как перелетные птицы. Точно так же и в Черном море сельди на зиму передвигаются из северо-западной части моря на юг, в более теплые места. Из Азовского моря они уходят осенью через Керченский пролив в Черное море и направляются вдоль кавказских берегов к югу (Малятский, 1931). Ниже приводятся данные, заимствованные у Киселевича («Сельди северного Каспия», 1937, стр. 70) и из журнала «Рыбное хозяйство» о средних годовых уловах каспийских сельдей по пятилетиям, в тысячах центнеров:
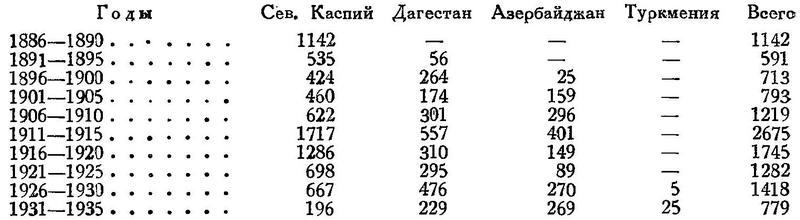
Начиная с 1937 г., обнаружилось повышение общих уловов сельдевых.
Бассейн Каспийского моря (В таблицу включены также сельди, не заходящие в пресную воду, если они имеют промысловое значение, кроме плохо известной Caspialosa curensis (Suworow).)
1 (13). Жаберных тычинок на 1-й дуге у взрослых обычно менее 90.
2 (10). На челюстях заметные зубы.
3 (8,9). Жаберных тычинок 45 и менее. Жаберные дуги толстые, крепкие.
4 (7). Верхний и нижний профили головы прямые, не закруглены. Голова низкая, клиновидная. Жаберные тычинки длиной не превосходят длины жаберных лепестков.
5 (6). Глаз большой, диаметр его не менее 6% длины тела (до конца средних лучей С) и менее полутора раз в длине рыла. Позвонков
наичаще 50. Небольшие рыбы, длиной до 200—250 мм. 3. Caspialosa saposhnikovi (Grimm)
6 (5). Глаз небольшой, диаметр его обычно менее 6% длины тела (но бывает до 6.4%) и обычно более полутора раз в длине рыла. Позвонков наичаще от 50 до 52 (у разных форм). Крупные рыбы, длиной свыше 250 мм. В пресную воду не входят. 1. Caspialosa brashnikovi (Borodin)
7 (4). Верхний и нижний профиль головы закруглены. Голова высокая, не клиновидная. Жаберные тычинки длиннее жаберных лепестков. Небольшие рыбы. В пресную воду не входят 4. Caspialosa sphaerocephala (Berg)
8 (3, 9). Жаберных тычинок у взрослых обычно 50—55. Жаберные дуги тонкие, слабые. Длина жаберных тычинок в полтора раза больше длины жаберных лепестков. Небольшие рыбы. 5. Caspialosa suworowi (Berg)
9 (3,8). Жаберных тычинок 60—90, наичаще 70—80. Жаберные дуги толстые, крепкие. Длина жаберных тычинок не больше длины жаберных лепестков. Крупные рыбы, длиной до 500 мм. 6. Caspialosa kessleri (Grimm)
10 (2). На челюстях слабые зубы.
11 (12). Жаберных тычинок 43—95, чаще 60—85, в среднем 72. Крупные рыбы, половозрелые в среднем 31 см. Caspialosa kessleri isp. volgensis imitans Berg
12 (11). Жаберных тычинок 63—95, наичаще 70—85, в среднем 78; тычинки на концах иногда булавовидно расширены. Мелкие рыбы, половозрелые длиной 20—22 см. Caspialosa kessleri isp. volgensis bergi Tanassijtschuk
13 (1). Жаберных тычинок на 1-й дуге у взрослых обычно более 90. На челюстях едва заметные зубы или их совсем нет.
14 (15). Тело низкое, удлиненное, высота его обычно менее 25% длины тела, в среднем 23%. Позвонков наичаще 52. Крупные рыбы длиной до 400 мм. Caspialosa kessleri isp. volgensis (Berg)
15 (14). Тело высокое, высота его обычно более 25% длины тела. Позвонков наичаще 50. Небольшие рыбы, длиной обычно меньше 250 мм. 7. Caspialosa caspia (Eichwald)
16 (23). Жаберных тычинок в среднем меньше 130.
17 (20). Жаберных тычинок в среднем больше 100.
18 (19). Жаберных тычинок в среднем 122. Высота тела в среднем 25—29% длины тела. Диаметр оплодотворенной икринки в среднем более 2 мм, оболочка ее тонкая, не покрытая илом. Caspialosa caspia typ.
19 (18). Жаберных тычинок в среднем 117. Тело ниже, высота его в среднем 25—26%. Диаметр икринки в среднем менее 2 мм, оболочка ее плотная, обычно покрытая тонким слоем ила. 7d. Caspialosa caspia aestuarina Berg
20 (17). Жаберных тычинок в среднем меньше 100.
21 (22). Жаберных тычинок в среднем 86. Высота тела в среднем 26.5%. — Сев.-вост. часть Каспия.7е. Caspialosa caspia salina Svetovidov
22 (21). Жаберных тычинок в среднем 80. Высота тела в среднем 28.5%. — Южн. Каспий.11. Caspialosa caspia persica Iljin
23 (16). Жаберных тычинок в среднем 140 (у 2- и 3-годовалых). Высота тела 28%. — Южн. Каспий. 7g. Caspialosa caspia knipowitschi Iljin
Бассейн Черного и Азовского морей. (А. Н. Световидов (Зоол. журн., 1943, стр. 231) правильно считает черноморских сельдей из рода Caspialosa относящимися к тем же видам, что и каспийские: Caspialosa maeotica = Caspialosa brashnikovi, Caspialosa pontica = Caspialosa kessleri, Caspialosa tanaica и Caspialosa nordmanni = Caspialosa caspia.)
1 (5, б). На челюстях заметные зубы.
2 (3, 4). Жаберных тычинок на 1-й дуге 27—39. 1а. Caspialosa brashnikovi maeotica (Grimm)
3 (2, 4). Жаберных тычинок 40—73. 6а. Caspialosa kessleri pontica (Eichwald)
4 (2, 3). Жаберных тычинок 76—95. 7а. Caspialosa caspia nordmanni (Antipa)
5 (1, 6). На нижней челюсти едва заметные зубы. Жаберных тычинок
62—93. 7с. Caspialosa caspia palaeostomi Sadowsky
6 (1, 5). На нижней челюсти нет зубов. Жаберных тычинок 65—78. 7b. Caspialosa caspia tanaica (Grimm)