(Clupeonella Kessler, 1877) Род Клупеонеллы, Тюльки, Сардельки, Каспийские кильки, Genus Clupeonella Kessler, 1877 (Sprat, Kilka, Tyulka) 6 видов
Словарь названий пресноводных рыб СССР. Г.У.Линдберг и А.С.Герд 1972 г.
3.3.0. Clupeonella Kessler, 1877 — Тюльки.
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
Род Тюльки, или Каспийские кильки
(Glupeonella), содержит 4 вида мелких
сельдевых рыб, обитающих в Черном,
Азовском и Каспийском морях и в их
бассейнах. Брюхо у тюлек сжатое с боков,
снабженное на всем протяжении от горла
до анального отверстия 24—31 сильными
шиповатыми чешуйками. Брюшные
плавники примерно под передней третью
спинного. В анальном плавнике два последних
луча удлинены, как у сардин и сардинелл.
Рот верхний, беззубый, маленький,
верхнечелюстная кость не заходит назад
далее переднего края глаза. Икринки
плавучие, с очень крупной фиолетовой
жировой каплей, с большим кругожелтковым
пространством. Позвонков 39—49.
Тюльки — евригалинные и евритермные рыбы,
живущие как в солоноватой, до 13‰,
так и в пресной воде при температуре от
0 до 24°С.
Каспийские кильки — обыкновенная,
анчоусовидная и большеглазая — служат
основным кормом хищных рыб Каспия.
Ими кормятся хищные сельди, белуга,
тюлени.
Промысел каспийских килек начался
в 20-х годах и производился вначале
вблизи берегов. С начала 50-х годов стал
интенсивно развиваться другой вид
промысла, основанный на приманивании
рыбы светом спущенной в воду сильной
электрической лампы. Облов
собирающейся к лампе кильки производился
сначала подъемными коническими сетями,
а затем и через раструб спускаемого
вблизи лампы шланга, засасывающего рыбу
насосом. Промысел кильки настолько
развился, что ее улов составил к середине
60-х годов более трех четвертей всего
улова рыбы на Каспии.
Фауна СССР. Рыбы. Том II, вып.1. Сельдевые (Clupeidae). А.Н.Световидов 1952 г.
8. Род CLUPEONELLA KESSLER — ТЮЛЬКИ И САРДЕЛЬКИ
— Clupeonella Кесслер, Рыбы Арало-касп.-понт. обл., 1877 : 187, табл. VI, фиг. 24 (тип: личинки Clupeonella grimmi Kessler).
— Harengula (non Valenciennes, 1847) Берг, Мат. к позн. русск. рыбол., II, 3, 1913 : 42; там же, IV, 6, 1915 : 4; Рыбы пресных вод, 1923 : 28.
— Clupeonella Regan, Ann. Mag. Nat. Hist. (8), XIX, 1917 : 377; Берг, Рыбы пресных вод, I, 1932 : 101.
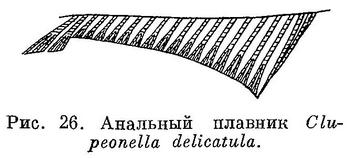
Верхняя челюсть без медиальной вырезки. Последние два луча анального плавника, как у Sardinella, Sardinops и Sardina, удлиненные и образуют лопасть (рис. 26), но удлиненные чешуи (ala) при основании лопастей хвостового плавника отсутствуют. Кожистого выроста на вертикальной части ключицы, как у Sardinops и Sardina, нет, но радиальные бороздки на крышечной кости в отличие от них отсутствуют. Жировые веки зачаточные или почти отсутствуют. Задние концы мочеточников с пучком многочисленных (до 10) выростов, свободно свисающих у личинок, у взрослых скрыты в стенке тела между половым сосочком и анальным плавником (См.: Владимиров, Докл. АН СССР, LVIII, 6, 1948 : 761—763.). Брюхо сжатое с боков или слегка закругленное, брюшные килевые чешуи хорошо развитые, на всем протяжении от горла до начала анального плавника образуют явственный киль. Рот небольшой: сочленение нижней челюсти с черепом позади вертикали переднего края глаза, не достигая вертикали середины его, задний конец верхнечелюстной кости едва заходит на вертикаль переднего края глаза. Нижняя челюсть выдается вперед. Зубов на челюстных и нёбных костях и на сошнике нет. Владимиров (1949 : 20) указывает немногочисленные зубы на нижней челюсти, встречаемые не у всех экз. Clupeonella delicatula delicatula. На костях таких зубов я не обнаружил. Повидимому, это не зубы, а зазубренный край нижней челюсти. Жаберные тычинки верхней части жаберной дуги не налегают на тычинки нижней части дуги, тычинки удлиненные, тонкие, 39—67. Отростки плавательного пузыря помещаются только в prooticum, в pteroticum их нет. Пятен на теле нет. Икринки пелагические, с очень большой жировой каплей и большим околожелтковым пространством. Небольшие рыбы длиной менее 200 мм.
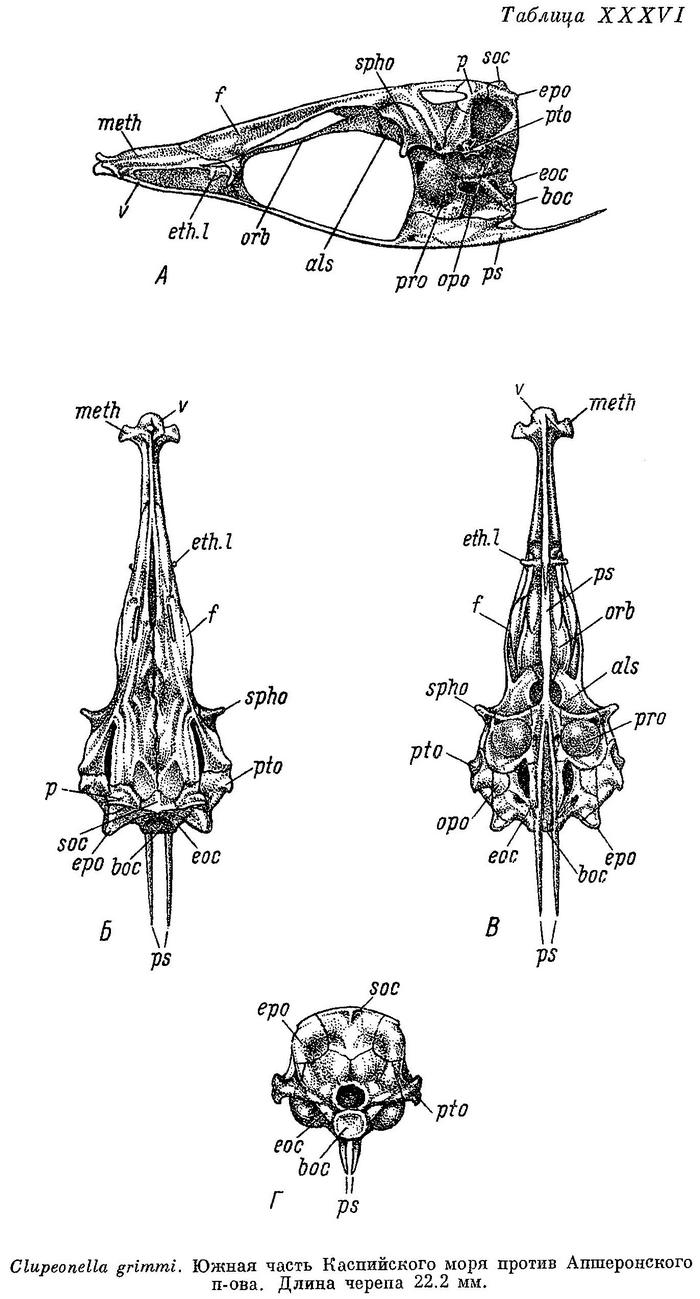
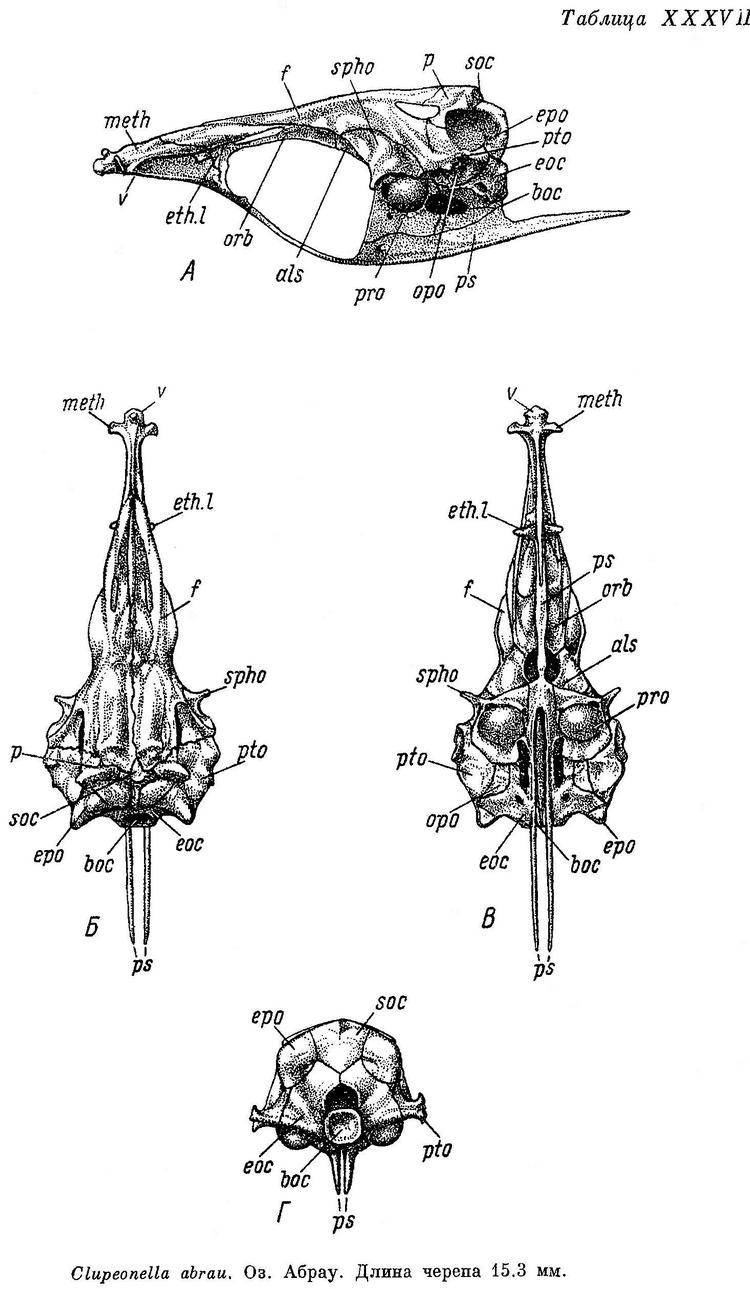
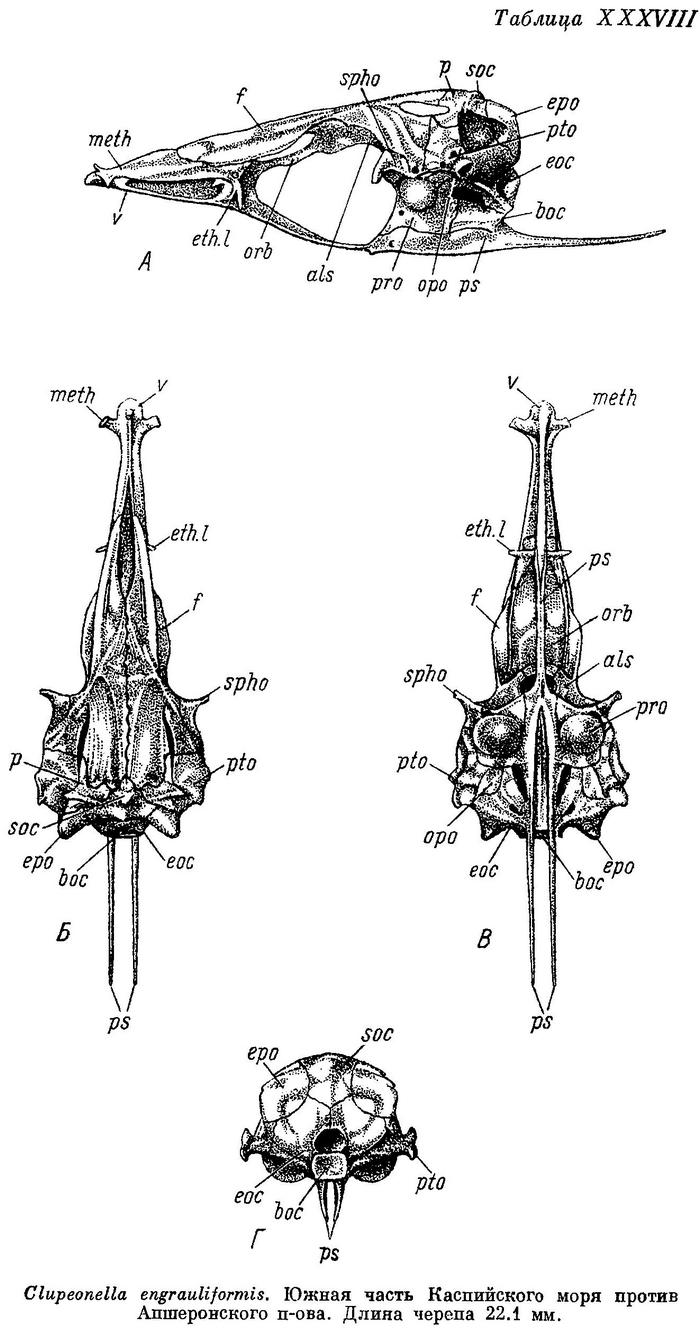
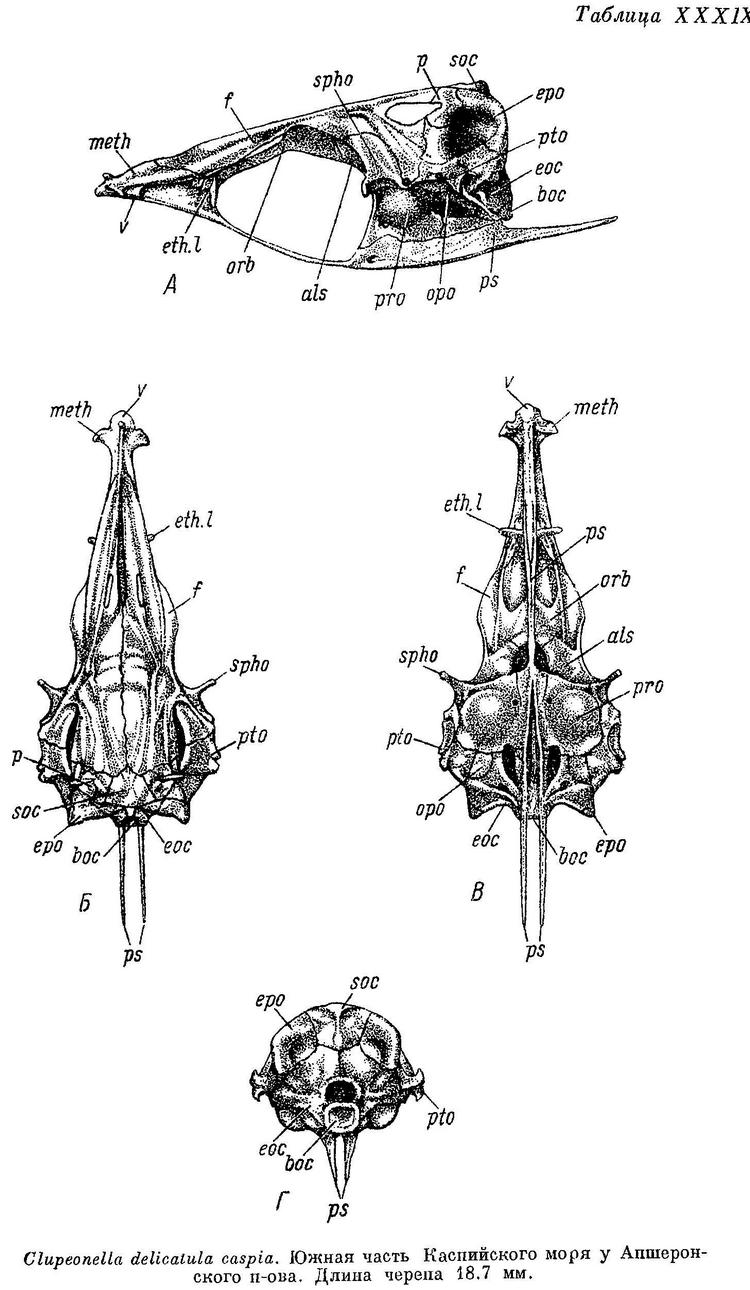
Скелет. Bulla prootica образует сбоку черепа (табл. XXXVI—XXXIX, А) большое и выпуклое вздутие, занимающее большую часть поверхности этой кости. Каждый отросток плавательного пузыря на occipitale laterale расположен в бороздке у переднего края этой кости, вдоль заднего края ее короткий гребень; отверстие, ведущее в череп, расположено в верхней части, под opisthoticum. Fenestra auditiva значительных размеров, близко расположена к bulla prootica, не соприкасается с ней, широко отделена от отверстия n. vagus. Fossa praeepiotica большая и высокая, широким краем обращена кверху, по длине равная или слегка большая или меньшая, чем foramen temporale, последнее наиболее высокое посредине. Поверхность для причленения hyomandibulare состоит из двух разобщенных частей на sphenoticum и pteroticum. Сошник почти прямой, передняя часть парасфеноида изогнутая, тонкая, с закругленным верхним краем, крылья парасфеноида тонкие и длинные, заостренные на конце. Нисходящего отростка базисфеноида нет. Задний край supraoccipitale и каждого epioticum на черепе сверху (Б) расходится в обе стороны под тупым углом. Задний конец pteroticum без шипа на конце, передний конец sphenoticum в виде удлиненного и тонкого отростка, направленного в сторону и слегка вперед. Из-под переднего конца лобных костей слегка выдаются в стороны задние крылья ethmoidalia lateralia. Поперечные отростки мезэтмоида направлены в стороны и немного вперед, одинаковой ширины на всем протяжении, тупые и закругленные на концах. Череп узкий, более широкий у Clupeonella delicatula, наиболее узкий у Clupeonella grimmi, наибольшая ширина его около 2.5 раз у первого и 2.75 у второго в длине его, наибольшая ширина лобных костей около 3.5 у первого и около 5 раз у второго. Сошник на черепе снизу (В) слегка закругленный, головка его с небольшим продольным закругленным гребнем, передняя часть парасфеноида слегка вдоль вогнутая, более широкая спереди, крылья парасфеноида разделяются под передним концом bulla prootica или немного позади. Задний конец opisthoticum без отростка. С каждой стороны на черепе сзади (Г) неглубокие fossae temporales posteriores. Верхняя часть hyomandibulare разделена на две совершенно обособленные головки для сочленения с черепом, с большой выемкой между ними. Quadratum без выемки в верхнем крае. Нижнечелюстная и сочленовная кости высокие, сочленовная короче нижнечелюстной. Предкрышечная довольно широкая, немного уже крышечной. Описание дано по многим скелетам Clupeonella delicatula delicatula и Clupeonella delicatula caspia, нескольким скелетам Clupeonella engrauliformis и Clupeonella grimmi и одному Clupeonella abrau.
Солоноватоводные или пресноводные рыбы, некоторые из солоноватоводных заходят в лиманы и реки.
4 вида с 1, возможно, 2 подвидами в Каспийском, Азовском и опресненных частях Черного морей и их бассейне, в оз. Абрау и Абулионд (басс. Мраморного моря).
Судя по размерам (до 45 мм), а также времени (зимой) и глубине нахождения — 146—457 м (80—250 саж.), личинки, описанные Кесслером под названием Clupeonella grimmi, могли относиться только к этому виду. Систематическое положение и распространение Clupeonella abrau весьма своеобразно. Краниологически и по ряду других особенностей этот вид, живущий в озерах, расположенных близ Черного и Мраморного морей, ближе к каспийским видам Clupeonella engrauliformis и Clupeonella grimmi, а не к встречающемуся также и в Черном море Clupeonella delicatula. Более узкий череп и более низкое тело, чему Clupeonella delicatula, исключают вероятность общности происхождения Clupeonella abrau с этим видом, у которого при переходе к пресноводному образу жизни (Clupeonella delicatula caspia m. tscharchalensis) тело становится значительно более высоким, а череп, хотя и незначительно, более широким.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (6). Грудные плавники на конце заостренные. Голова короткая и широкая, межглазничный промежуток не менее 16% длины головы.
2 (5). Тело и в особенности брюшко сжатые с боков, брюшко заостренное, брюшные шипы хорошо заметные.
3 (4). Голова широкая, межглазничный промежуток не менее 17.5% длины головы. Черное, Азовское и Каспийское моря и их бассейн. 1. Clupeonella delicatula (Nordmann)
4 (3). Голова узкая, межглазничный промежуток не более 17.5% длины , головы. Озера Абрау и Абулионд. *3. Clupeonella abrau (Maljatskij)
5 (2). Тело вальковатое, низкое, брюшко закругленное, брюшные шипы слабо заметные. *2. Clupeonella engrauliformis (Borodin)
6 (1). Грудные плавники на конце закругленные. Голова большая и суженная, межглазничный промежуток не более 15.5% длины головы. *4. Clupeonella grimmi Kessler
Ископаемые виды рода Clupeonella по основным из рассматриваемых особенностей более примитивны, чем современные, причем Богачев (1938 : 47) отмечает, что один из известных понтических видов (Clupeonella vexata) имеет значительное сходство с современным Clupeonella engrauliformis. Однако голова у Clupeonella vexata значительно крупнее, чем даже у наиболее большеголового из современных видов (Clupeonella grimmi). У другого ископаемого вида (Clupeonella binagadensis), провизорно относимого тоже к роду Clupeonella, длина головы еще больше. По расстоянию спинного плавника от конца рыла, составляющему около 49—50% длины тела, Clupeonella vexata близок к наиболее примитивному из современных видов.
Помимо изменения ширины в межглазничной области, ширина головы изменяется и в других ее частях, почему выше, ссылаясь на изменение межглазничного промежутка, говорилось об изменении ширины головы вообще. Изменение ширины головы в других ее частях у видов рода Clupeonella видно из данных измерений ширины черепа:

Из видов рода Clupeonella наиболее подвижный образ жизни ведет Clupeonella delicatula, каспийская форма которого (Clupeonella delicatula caspia) совершает миграции из южной части моря в северную и частично заходит даже в реки; в море эта форма держится ближе к берегам. Азовская форма этого вида (Clupeonella delicatula delicatula) совершает меньшие передвижения. Clupeonella engrauliformis держится в некотором отдалении от берегов в южной и средней частях Каспийского моря, иногда подходит и в прибрежную зону. Clupeonella abrau, населяющий небольшое пресноводное озеро, совершает более ограниченные миграции. Наконец, Clupeonella grimmi живет главным образом в южной части Каспийского моря и отчасти в средней, держится в открытом море в галистатической области, никогда не подходя к берегам, и на значительно больших глубинах, чем другие виды рода.
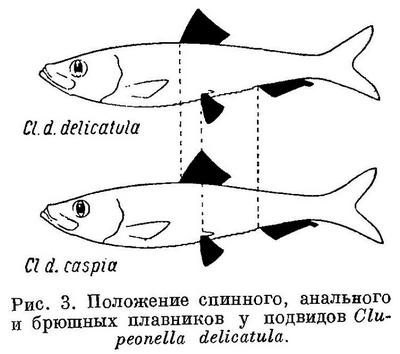
Соответственно положению спинного плавника изменяется положение анального и брюшных плавников и у подвидов в пределах некоторых видов подсем. Clupeinae. Естественно, что степень различия рассматриваемых признаков между подвидами менее степени различий их у видов.
Положение анального и брюшных плавников относительно переднего конца тела, точно так же как первого относительно спинного и второго относительно анального у подвидов Clupeonella delicatula соответственно положению спинного плавника относительно переднего конца тела, изменяется таким же образом, как и у родов и у видов подсем. Clupeinae (табл. 3, рис. 3).

При сопоставлении состава фауны сельдевых Каспийского, Черного и Азовского морей обращают на себя внимание несколько особенностей. Одна из этих особенностей заключается в том, что фауна рыб Черного моря, как известно, значительно более богатая и разнообразная по своему составу, чем Каспийского, более богата и разнообразна и в отношении сельдевых, но не видами и более низкими формами (подвидами), а родами. Здесь, помимо общих с каспийскими родом Clupeonella и подродом Caspialosa, имеются еще 3 рода из этого семейства — Sprattus, Sardina, Sardinella и подрод Alosa s. str., широко распространенных, кроме Черного моря, в Средиземном море и в Атлантическом океане, а некоторые из них, кроме того, в Индийском и Тихом океанах. Эта особенность черноморской фауны сельдевых, свойственная, как отмечалось выше, вообще фауне Черного моря, обусловлена, как известно, инвазией в Черное море средиземноморской фауны после прорыва Босфора.
Еще больший интерес представляет другая особенность черноморских и каспийских сельдевых, не отмеченная до сего времени и заключающаяся в том, что общие обоим морям род Clupeonella и подрод Caspialosa в Каспийском море как в видовом отношении, так и по количеству более низких форм (подвидов) представлены значительно более богато, чем в Черном и Азовском морях. Эта особенность была отмечена автором ранее в предварительном сообщении (Световидов, 1941 : 807, предст. 13 VI 1940). Эта статья должна была быть напечатана одновременно со статьей А. А. Михайловской (1941 : 559, предст. 24 VI 1940), которая была знакома с выводами автора и, говоря о богатстве каспийских сельдей, сделала ссылку на нее. Вследствие того, что статья автора была опубликована позднее: статьи А. А. Михайловской, эта ссылка в ее статье помещена не была.
Состав фауны сельдевых этих родов в Каспийском и Черном и Азовском морях таков (В скобках указано количество подвидов.):

1 Четвертый вид этого рода Clupeonella abrau встречается в оз. Абрау и Абулионд; о нем сказано особо ниже.
Род Clupeonella в Каспийском море представлен 2 эндемичными видами и лишь одним — формой того же вида, что и в Каспийском, — в Черном и Азовском морях. В Каспийском море из рода Alosa имеются 6 видов, из которых 3 свойственны только этому морю; в Черном же и Азовском морях из этого рода встречается только 3 вида, представленные здесь формами видов, общих с каспийскими. Таким образом, в Черном и Азовском морях нет ни одного вида, который не был бы представлен в Каспийском и притом значительно большим количеством форм, чем в Черном и Азовском морях, в то время как в Каспийском море имеются 2 вида из рода Clupeonella и 3 вида из рода Alosa, отсутствующие в Черном и Азовском морях.
Богатство сельдевых обоих этих родов в Каспийском море обусловлено преобладанием главным образом морских, непроходных видов и подвидов. Таковыми из числа видов являются Clupeonella grimmi и Clupeonella engrauliformis и Alosa saposhnikovi, Alosa sphaerocephala и Alosa curensis, в то время как проходная (Alosa kessleri s. l.) и полупроходная (Alosa caspia s. l.) сельди и тюлька (Clupeonella delicatula s. l.) представлены в обоих морях.
Необходимо также отметить некоторые биологические особенности каспийских и черноморско-азовских форм Alosa brashnikovi s. l. и Alosa caspia s. l., а именно то, что каспийские формы этих видов являются более морскими рыбами, чем черноморско-азовские. Все каспийские формы Alosa brashnikovi s. l. избегают пресной воды и размножаются только в море, между тем как черноморско-азовская форма этого вида (Alosa brashnikovi maeotica) заходит в совершенно опресненные участки Таганрогского зал., и в нижнюю часть дельты Дона. Имеются указания и на нерест Alosa brashnikovi maeotica почти в пресной воде. Среди каспийских форм Alosa caspia s. l. есть такие, которые мечут икру в солоноватой воде в море (Alosa caspia salina) или в предустьевых пространствах (Alosa caspia caspia), в то время как все черноморско-азовские формы этого вида мечут икру только в пресной воде, заходя в реки, озера и лиманы. То же самое следует сказать и о Clupeonella delicatula s. l.: Clupeonella delicatula caspia мечет икру как в пресной воде, так почти и по всему Каспийскому морю, между тем как Clupeonella delicatula delicatula в Азовском море нерестует в самых опресненных частях его — в Таганрогском заливе, в дельте Дона, в кубанских опресненных лиманах и лишь отчасти в море.
Заслуживает внимания и еще одна особенность каспийских форм обоих рассматриваемых родов, отмеченная ранее Киселевичем (1918 : 21), — их значительно большие размеры, по сравнению с черноморско-азовскими. Clupeonella delicatula caspia достигает в длину до 14 см, средние размеры ее около 8—10 см, между тем как размеры Clupeonella delicatula delicatula в Азовском море не превышает 9 см, имея в среднем около 7 см.
Наиболее важная особенность сельдевых обоих рассматриваемых родов заключается, однако, в том, что каспийские формы их различаются значительно больше, чем черноморско-азовские формы тех же видов.
Богатство каспийских сельдевых родов Clupeonella и Alosa как в видовом отношении, так и количеством форм общих видов, их гигантизм по сравнению с черноморско-азовскими формами, отмеченные выше биологические особенности и большая степень отличий обусловлены различиями в условиях развития и формообразования сельдевых как в Каспийском и Черном и Азовском морях, так и в тех морях, которые предшествовали им. Поэтому для рассмотрения условий развития и формообразования каспийских и черноморско-азовских сельдевых обоих этих родов и выяснения причин такого резкого различия в составе их, размерах, биологических особенностях и степени отличий необходимо обратиться к истории развития этих морей, так как история развития фауны, по вполне справедливому замечанию Колесникова (1940 : 6), не должна быть оторвана от истории развития бассейна, в котором она обитает.
Понто-каспийский морской бассейн, как известно, претерпел сложную геологическую историю. К концу миоцена, при сокращении Тетиса, одновременно с похолоданием климата, северо-восточная часть его отделилась, образовался сарматский и ряд других пришедших ему на смену бассейнов, фауна которых в дальнейшем развивалась в значительной степени изолированно, находясь, в отличие от фауны образовавшегося к этому времени Атлантического океана и стоявшего в связи с ним Средиземного моря, под сильным воздействием периодических опреснений и осолонений. В меньшей степени опреснения и бассейны с затрудненным сообщением с нормально соленым морским бассейном или даже замкнутые бассейны в северо-восточной части Тетиса были и раньше, в нижнем и среднем миоцене (Жижченко, 1940 : 42, 148).
Биологические взаимоотношения видов Clupeonella и Alosa с другими рыбами как в настоящее время в Каспийском море и Черном и Азовском морях, так несомненно и в морях, предшествовавших им в предыдущие эпохи, различны. Пелагическая фауна открытых частей Каспийского моря состоит исключительно из сельдевых, которые (все виды Clupeonella и формы Alosa caspia s. l.) являются здесь основными потребителями планктона (см. об этом Бенинг, 1938 : 33). Других пелагических рыб, кроме сельдевых, в галистатической области Каспийского моря в сущности нет. Население пелагиали более открытых частей Черного моря составляют в основном средиземноморские иммигранты как из сельдевых, но других родов. Что касается сельдевых из родов Clupeonella и Alosa, то они вместе с остальной автохтонной реликтовой фауной понтического озера-моря встречаются в опресненных частях Черного моря и в Азовском море. Тюлька (Clupeonella delicatula delicatula) встречается здесь только в Азовском море, в лиманах и в опресненных участках у берегов северо-западной части Черного моря, Румынии, Болгарии, в Феодосийском зал. и в оз. Палиастоми. Сельди в зимнее время держатся вдоль берегов Кавказа, у берегов Крыма, летом заходят в опресненные части Черного моря и в Азовское море и в реки.
В галистатической части Черного моря ни один из представителей этих двух родов не найден.
Таким образом, виды рода Clupeonella, живущие только в море и притом в открытых его частях (Clupeonella grimmi и Clupeonella engrauliformis), и морские, непроходные виды рода Alosa (Alosa saposhnikovi, Alosa sphaerocephala, Alosa brashnikovi s. l.), мечущие икру в море, в Черном море не могли образоваться или получить развитие потому, что пелагиаль открытых частей его после прорыва Босфора была заселена более сильными конкурентами, иммигрантами из Средиземного моря. Наоборот, можно полагать, что сельдевые из этих двух родов в настоящую фазу существования Черного моря испытывают в нем сильную депрессию, будучи оттеснены в наиболее опресненные части его и в Азовское море более жизненными средиземноморскими иммигрантами.
Совершенно иные условия оказались в Каспийском море, которое в средней и, в особенности, в южной части в настоящую фазу его существования представляет солоноватоводный, в южной части почти такой же морской бассейн, как Черное море. В этом бассейне пелагических рыб кроме сельдевых из родов Clupeonella и Alosa не оказалось, поэтому фауна пелагиали его, в особенности в южной и средней частях, могла формироваться только из этих рыб, и поэтому же они, в особенности морские непроходные, при отсутствии конкуренции с другими видами и получили здесь пышный расцвет; расцвет этот сопровождается и гигантизмом; аналогичное явление наблюдалось и в понтической реликтовой фауне, например в киммерийском бассейне (Колесников, 1940 : 930).
Подводя итоги всему сказанному об условиях развития и формообразования сельдевых из рода Clupeonella и Alosa, следует подчеркнуть, что богатство их в Каспийском море обусловлено расцветом их в этом море главным образом в настоящую фазу его существования, со времени последнего соединения его с морями, предшествовавшими Черному и Азовскому, которое могло быть в послехвалынское и в бакинское, а ранее в апшеронское и акчагыльское время (см. об этом: Колесников, 1940 : 928), а не в одном из морей, предшествовавших Каспийскому, как полагал Киселевич (1918 : 24). При этом расцвет их обусловили не «значительное протяжение и разнообразие климатических условий в различных частях бассейна» (Киселевич, 1918 : 24), а наличие в Каспийском море незаселенной другими рыбами пелагиали, которую могли заполнить только сельдевые рассматриваемых двух родов. Что касается приводимого К. А. Киселевичем мнения Сарса, что каспийские сельди «находятся в стадии незакончивщегося видообразования» (1918 : 24), то в стадии видообразования находится весь органический мир, даже наиболее древние его представители.
Среди других видов рода Clupeonella по своему распространению особый интерес представляет Clupeonella abrau. Этот вид встречается в небольшом пресноводном озере Абрау и, кроме того, в качестве особого вида (Clupeonella muhlisi), идентичного Clupeonella abrau или самое большее являющегося формой этого вида, описан из пресноводного озера Абулионд (басс. Мраморного моря). Что касается Clupeonella abrau, то он не может считаться эндемичным потому, что, как указывалось выше, найден и в оз. Абулионд. По ряду морфологических особенностей, положенных в основу системы семейства и отдельных видов, Clupeonella abrau относится к числу наиболее примитивных видов рода и наиболее близок не к встречающемуся также и в Черном и Азовском морях виду Clupeonella delicatula, а к видам, встречающимся только в Каспийском море.
Рыбы пресных вод СССР и сопредельных стран. Л.С.Берг 1948 г.
6. Род CLUPEONELLA KESSLER
Clupeonella Кесслер, Рыбы Арало-касп.-понт. обл., 1877, стр. 187, табл. VI, фиг. 24 (тип: Clupeonella grimmi, личинки).
Harengula Берг (non Valenciennes 1847), Мат. к позн. русск. рыбол., II, вып. 3, 1913, стр. 42; IV, вып. 6, 1915, стр. 4; Рыбы пресн. вод России, 1923, стр. 28.
Clupeonella Regan, Ann. Mag-. Nat. Hist. (8), XIX, 1927, p. 377 (ср. также ibidem, XVIII, 1916, p. 3).
Брюхо сжатое с боков, снабженное на всем протяжении от горла до анального отверстия ясно заметными, сильными шипообразными чешуйками, не лежащими в борозде, числом 24—31. В спинном плавнике III—IV 11—15 лучей. Брюшные плавники приблизительно под передней третью спинного, с I 6—7 лучами. Анальный плавник с II—III 14—20 лучами; два последних луча его увеличены, как у сардинок (фиг. 108).

При основании хвостового плавника нет удлиненных чешуй (ala). Зубов на сошнике нет, и вообще рот совершенно беззубый. На межчелюстных костях нет вырезки. На вертикальной части ключицы, смотрящей в жаберную полость, нет выступов. Рот верхний, нижняя челюсть выдается вперед. Ротовая щель небольшая; верхнечелюстная кость доходит лишь до вертикали переднего края глаза; сочленение нижней челюсти с черепом едва заходит за эту вертикаль. Век нет или зачаточные. Радиальных полос на крышечной кости нет. Жаберных тычинок 38—64. Чешуя легко опадающая. Пилорических придатков 8. Позвонков 39—49. (У Clupeonella grimmi, по Кесслеру, 48—49 позвонков; у Clupeonella delicatula я находил 43—44. Е. К. Суворов (1914, стр. 33) приводит для Clupeonella delicatula 40—42, для Clupeonella engrauliformis 44—46; подсчет 20 экз. Clupeonella delicatula из сев. Каспня, сделанный в 1930 г. Е. К. Суворовым, дал 39—43, наичаще 39 и 40, в среднем 40.4.) Яичники розовые или розовато-фиолетовые. Икра пелагическая, с очень большой жировой каплей, с большим околожелтковым пространством (Т. Расс). Пятен на боках тела не бывает. Длина взрослых до 17 см. От родов Clupea и Harengula отличается удлиненными последними лучами анального плавника.
Четыре-пять видов в бассейнах Черного и Каспийского морей. Одни входят из моря в дельты рек для икрометания, другие постоянно живут в пресной воде, третьи мечут икру в море и в реки не заходят. Личинки одного из видов этого рода в Каспийском море зимой встречаются на значительных глубинах, ниже 300 м (Н. М. Книпович, Тр. Касп. эксп. 1914—1915 гг., П., 1921, стр. 728.). В нижеследующем пропорции, если не оговорено, отнесены к длине до конца средних лучей С.
1 (8). Позвонков 39—47. Голова не более 23.5% длины тела.
2 (7). Тело сжатое с боков, не вальковатое. Брюшные шипы хорошо развиты. Грудные плавники два раза или меньше в расстоянии Р—V. Жаберных тычинок 41—64.
3 (6). Тело сравнительно высокое, высота его составляет не менее 20.5% длины тела (до конца средних лучей С).
4 (5). Высота тела 20.5—25% его длины. Жаберных тычинок 52—64 .1. Clupeonella delicatula (Nordmann)
5 (4). Высота тела 23.5—30% его длины. — Оз. Чархал и Волга. 1(1). Clupeonella delicatula morpha tscharchalensis (Borodin)
6 (3). Тело сравнительно низкое, высота его 14—20.5% его длины. Брюшных шипов наичаще 25—26. — Оз. Абрау в зап. Закавказье. 2. Clupeonella abrau Maliatskij
7 (2). Тело вальковатое, низкое, высота тела не более 19% длины его. Брюшные шипы слабо развиты. Грудные плавники более двух раз в расстоянии Р—V. Жаберных тычинок 57—63. Позвонков 44—47. — Каспийское море. 3. Clupeonella engrauliformis (Borodin)
8 (1). Позвонков 44—49, в среднем 47. Голова более 23.5% длины тела, глаза большие. — Каспийское море. 4. Clupeonella grimmi Kessler