(Lophiiformes Garman, 1899) Отряд Лофииформес, Удильщикообразные, Морские черти, Ногопёрые, Рукопёрые, Order Lophiiformes Garman, 1899 (Anglerfishes) 18 семейств
Информация из Интернета
Новый отряд впервые описал в 1899 году американский учёный Самуэль Гарман (Самуэль Уолтон Гарман — Samuel Walton Garman, 1843 — 1927, американский герпетолог, палеонтолог и ихтиолог, в 1870 году окончил Государственный педагогический университет штата Иллинойс (Illinois State Normal University), в 1871 году стал профессором естественных наук в семинарии Ферри-Холл (Ferry Hall Seminary), затем учился у Луи Агассиса (Louis Agassiz), в 1872 году получил диплом, сопровождал Эдварда Копа (Edward Drinker Cope) в 1872 году в Вайоминг для сбора фоссилий (окаменелостей), с Луи Агассисом предпринял исследовательскую поездку к Магелланову проливу, с 1870 года работал помощником директора отделения герпетологии и ихтиологии в Музее сравнительной зоологии Гарвардского университета (Harvard's Museum of Comparative Zoology), его работа касалась классификации рыб, особенно акул, а также рептилий и амфибий, автор описания более 200 новых видов рыб, в его честь названо тринадцать новых таксонов рыб).
Лофииформес, Удильщикообразные, Морские черти, Ногопёрые, Рукопёрые (Lophiiformes) — отряд морских лучепёрых рыб, обитающих по всему Мировому океану. Отряд удильщикообразных делится на 5 подотрядов: Удильщиковидные (Lophioidei), Клоуновидные (Antennarioidei), Хаунаксовидные (Chaunacoidei), Нетопыревидные (Ogcocephaloidei) и Цератиевидные (Ceratioidei). Эта классификация основана на филогенетических выводах Т. Питча и Д. Гробекера (Pietsch & Grobecker, 1987).
Название отряда Lophiiformes образовано от имени типового рода Lophius Linnaeus, 1758, название которого происходит от древнегреческого слова λοφια (lophia) — «грива», «спина», «гребень», «щетина», этимология которого до конца не выяснена. Некоторые ученые связывают его со своеобразным гребнем, проходящим вдоль всей спины. Другие исследователи придерживаются мнения, что челюсти этой рыбы напоминают щетину или гребень. Окончание отрядов «–formes» происходит из латинского языка и означает «похожей формы». Английское название «anglerfishes» — «рыбы-рыболовы», «рыбы-удильщики», соответствует русскому названиям отряда «удильщикообразные», и указывает на длинный видоизменённый первый луч спинного плавника, снабжённый приманкой (эской) и напоминающий удочку рыболова. А благодаря необычному и непривлекательному виду головы — этих хищников прозвали «морскими чертями». Из-за того, что эти рыбы могут передвигаться по морскому дну, отталкиваясь от него несколько видоизменёнными плавниками, их ещё называют «рыбы-лягушки», «ногопёрые» или «рукопёрые».
Найденные окаменелости (fossils) ранних удильщикообразных рыб были датированы нижним третичным периодом (палеоген). Время появления некоторых современных родов отряда Морские черти: начало эоцена — Lophius (Лофиусы), Antennarius (Антеннариусы), Brachionichthys (Брахионихтисы); середина эоцена — Ogcocephalus (Огкоцефалусы); конец эоцена — Dibranchus (Дибранхусы), Chaunax (Хаунаксы). [Sepkoski, Jack. A compendium of fossil marine animal genera // Bulletins of American Paleontology — Paleontological Research Institution, 2002. — Vol. 364. — P. 560.]
Филогенетическое исследование митохондриального генома показало, что Удильщикообразные рыбы диверсифицировались (разнообразились) за короткий период от раннего до среднего мелового периода, между 130 и 100 миллионами лет назад. [Miya, M.; T. Pietsch; J. Orr; R. Arnold; T. Satoh; A. Shedlock; H. Ho; M. Shimazaki; M. Yabe (2010). "Evolutionary history of anglerfishes (Teleostei: Lophiiformes): a mitogenomic perspective". BMC Evolutionary Biology. 10: 58.]
Удильщикообразные рыбы имеют голое тело причудливой формы, часто с большим количеством кожных выростов или покрытое костными бугорками, шипиками или бляшками. Пелагические формы наиболее сжаты с боков, тогда как бентосные формы обычно сильно сжаты дорсовентрально (приплюснуты), часто с большими ртами, направленными вверх. Передняя часть спинного плавника, если он присутствует, расположена прямо над пастью и имеют форму «удочки» (Illicium, иллиций) с приманкой (Esca, эска) для привлечения добычи. Именно эта особенность дала таксону его название. Брюшные плавники, если имеются, с одним шипом и четырьмя (редко) или пятью мягкими лучами, сидят на горле, перед грудными плавниками. Одним из отличительных признаков удильщикообразных являются твёрдые грудные плавники, служащие для опоры. Они поддерживаются 2—3 скелетными элементами, из которых нижний сильно увеличен, обычно расширен на конце и может совершать вращательные движения. Это позволяет донным рыбам использовать грудные плавники, как бы согнутые в локте, для ползания по грунту. Жаберное отверстие небольшое трубчатое, расположено у основания грудного плавника или позади него (редко частично перед ним). Лучей жаберной перепонки — бранхиостегальных лучей (branchiostegal rays) — 5—6. Первый позвонок сросся с черепом. Рёбер нет. Грудные лучевые кости 2—5, узкие и удлинённые. Плавательный пузырь, при его наличии, закрытый и физиологически активный.
Все удильщикообразные рыбы плотоядны и поэтому приспособлены для поимки добычи. У большинства видов широкий рот простирается по всей передней окружности головы, а на обеих челюстях располагаются полосы наклоненных внутрь зубов. Зубы могут быть вогнутые, чтобы не мешать скольжению объекта в желудок, но предотвращать его выход изо рта. Морские черти способны расширять и челюсть, и живот, так как их кости тонкие и гибкие, до огромных размеров, что позволяет проглатывать добычу в два раза больше, чем все их тело. Из-за небольшого количества пищи, доступной в окружающей среде, эта адаптация позволяет удильщикообразным запасать пищу, когда её много. Глубоководные виды по цвету варьируют от тёмно-серого до тёмно-коричневого, с большими головами и огромными ртами в форме полумесяца, заполненными длинными, похожими на клыки зубами, загнутыми внутрь для эффективного захвата добычи. Их длина может варьироваться от 2—18 см, при этом некоторые виды достигают размера 120 см. Различие по длине в значительной степени связано с половым диморфизмом — самки намного крупнее самцов. Ногопёрые, обитающие на мелководье, устраивают засады и часто замаскированы под камни, губки или водоросли.
У большинства взрослых самок цератиидных удильщиков есть люминесцентный орган, называемый эска, на конце модифицированного спинного луча (иллициума или удочки). Предполагается, что этот орган служит очевидной цели — заманивать добычу в тёмной глубоководной среде, но также служит для привлечения внимания самцов к самкам для облегчения спаривания. Источником люминесценции являются симбиотические бактерии, обитающие внутри и вокруг эски, заключённые в чашеобразный отражатель, содержащий кристаллы, вероятно, состоящие из гуанина. У некоторых видов бактерии, размещённые внутри эски, неспособны к люминесценции независимо от рыбы-хозяина, что позволяет предположить, что они развили симбиотические отношения, и бактерии не могут самостоятельно синтезировать все химические вещества, необходимые для люминесценции и зависят от рыбы. Электронная микроскопия этих бактерий у некоторых видов показывает, что это грамотрицательные палочки, в которых отсутствуют капсулы, споры или жгутики. У них двухслойные клеточные стенки и мезосомы. Через поры эска соединяет с морской водой, что позволяет удалять мертвые бактерии и отходы живых клеток, а также позволяет рН оставаться постоянной. Все это, а также постоянная температура батипелагической зоны, в которой обитают эти рыбы, имеют решающее значение для долгосрочной жизнеспособности бактериальных культур. Эска всегда открыта наружу, поэтому не исключено, что бактерии попадают в рыбу из морской воды. Тем не менее, похоже, что каждый вид использует свой собственный вид бактерий, и эти бактерии никогда не были обнаружены в морской воде. Есть предположение, что эска выделяет бактерии во время нереста, и бактерии тем самым переносятся на икру.
Подотряды Ogcocephaloidei и Ceratioidei — объединены наличием таких признаков, как редуцированная до небольшого остаточного элемента вторая спинная колючка (за исключением Diceratiidae, у которых она хорошо развита), отсутствие третьей спинной колючки и её птеригиофора, сращенная с черепом задневисочная кость.
Ogcocephaloidei — нетопыревидные. Вторая спинная колючка редуцирована и погружена под кожу головы; жаберные лепестки на первой дуге отсутствуют (имеются в редуцированном виде у некоторых Ceratioidei); птеригиофор иллиция и третьей колючки спинного плавника без дорсальных расширений, уплощенных с боков; брюшной плавник имеется или отсутствует.
Подотряд Chaunacoidei — хаунаксовидные. Сестринская группа всех вышеприведенных таксонов, поскольку также характеризуется погруженной под кожу головы второй спинной колючкой (удлиненной у Chaunacidae) и отсутствием жаберных лепестков на первой жаберной дуге. Тело шарообразное; кожа покрыта зубчиками; только иллиций, других колючек спинного плавника нет; рот косой; жаберное отверстие позади основания грудного плавника; лучей анального плавника — 5—7; окраска тела от розовой до глубокой красновато-оранжевой. Максимальный размер около 35 см.
Подотряд Antennarioidei — клоуновидные. Сестринская группа всех вышеприведенных таксонов, поскольку также обладает мелкими икринками и личинками (много мельче, чем таковые у Lophioidei), редуцированным (до трёх и менее) числом колючек спинного плавника. Колючий спинной плавник состоит из трёх отдельных головных колючек (первая модифицирована в иллиций [«удилище»]; вторая колючка может быть короткой, но никогда не бывает погруженной под кожу); птеригиофоры иллиция и третьей колючки спинного плавника с сильно уплощенными с боков дорсальными расширениями.
Подотряд Lophioidei — удильщиковидные. Считаются примитивной сестринской группой всех приведенных выше таксонов. Брюшной плавник имеется; колючий спинной плавник за головой, с одной-тремя колючками; четвёртая фарингобранхиалия есть, несёт зубы; ложножабра крупная; тело голое; лобные кости соединяются.
Долгое время считалось, что Удильщикообразные связаны с Батрахообразными (Batrachoidiformes). Но молекулярно-филогенетические исследования Shedlock et al. (2009) и Miya et al. (2010), однако, предполагают, что Lophiiformes связаны с Иглобрюхообразными (Tetraodontiformes) и с рыбами-кабанами из семейства Caproidae.
Распространение (карта): весь Мировой океан. Некоторые из них являются пелагическими (обитают вдали от морского дна), а другие — бентическими (обитают близко к морскому дну), на больших глубинах (Ceratiidae) или на континентальном шельфе (Antennariidae и Lophiidae).
Поскольку удильщикообразные являются приспособленцами-собирателями, они демонстрируют диапазон предпочтительной добычи с рыбой в крайних пределах спектра размеров, показывая при этом повышенную избирательность к определённой добыче. Одно исследование, посвящённое изучению содержимого желудков тонкопёрой рыбы-удильщика у тихоокеанского побережья Центральной Америки, показало, что эти рыбы в основном ели две категории донных жертв: ракообразных и костистых рыб. Наиболее частой добычей были креветки пандалиды. 52% исследованных желудков были пустыми, что подтверждает наблюдения о том, что рыба-удильщик мало потребляет энергии.
Некоторые удильщики отличаются экстремальным половым диморфизмом и половым симбиозом маленького самца с гораздо более крупной самкой, что наблюдается в подотряде Ceratioidei. У этих видов самцы могут быть на несколько порядков меньше самок. Так морские дьяволы (Ceratiidae) используют необычный способ спаривания. Поскольку плотность населения здесь очень небольшая, встречи также очень редки, поэтому найти себе пару проблематично. Когда учёные впервые начали отлавливать цератиидных удильщиков, они заметили, что все экземпляры были самками. Эти особи были размером в несколько сантиметров, и почти все они были с прикреплёными паразитами. Оказалось, что эти «паразиты» были сильно редуцированными мужскими особями. Это указывает на то, что некоторые таксоны удильщиков используют полиандрию (многомужество) в спаривание. Свободноживущие самцы и «незаражённые» самки у этих видов никогда не имели полностью развитых гонад. Таким образом, самцы никогда не созреют, не прикрепившись к самке, и умирают, если не могут ее найти.
При рождении мужские цератииды уже имеют чрезвычайно хорошо развитые органы обоняния, которые обнаруживают запахи в воде. У самцов некоторых видов также развиваются большие узкоспециализированные глаза, которые могут помочь в идентификации партнеров в темноте. Самцы цератиидов значительно меньше самок удильщиков и могут иметь проблемы с поиском пищи в глубоком море. Кроме того, пищеварительный тракт некоторых самцов отстает в росте, что не позволяет им питаться. У некоторых таксонов есть челюсти, которые никогда не подходят или не эффективны для поимки добычи. Эти особенности означают, что самец должен быстро найти самку удильщика, чтобы предотвратить свою смерть. Чувствительные органы обоняния помогают самцу обнаруживать феромоны, которые сигнализируют о близости самки удильщика.
Методы, которые используют удильщикообразные для поиска партнёров, различаются. У некоторых видов крошечные глаза, которые не подходят для идентификации самок, в то время как у других недоразвитые ноздри не позволяют им эффективно находить самок по запаху. Когда самец находит самку, он кусает её кожу и высвобождает фермент, который переваривает кожу его рта и её тела, сплавляя пару до уровня кровеносных сосудов. Самец становится зависимым от самки-хозяина в своем выживании, получая питательные вещества через общую систему кровообращения, а взамен обеспечивает самку спермой. После слияния самцы увеличиваются в объеме и становятся намного крупнее свободноживущих самцов этого вида. Они живут и сохраняют репродуктивную функцию до тех пор, пока живёт самка, и могут принимать участие в нескольких нерестилищах. Этот крайний половой диморфизм гарантирует, что когда самка готова к нересту, у неё сразу же появится половой партнёр. Несколько самцов могут быть одновременно прикреплены к одной самке (до восьми самцов у некоторых видов), хотя некоторые таксоны, по-видимому, имеют правило «один самец на самку».
Симбиоз — не единственный способ размножения удильщиков. В самом деле, многие семейства, в том числе Melanocetidae, Himantolophidae, Diceratiidae и Gigantactinidae, не показывают никаких признаков мужского симбиоза. У самок некоторых из этих видов есть большие развитые яичники, а у свободноживущих самцов большие семенники, что позволяет предположить, что эти половозрелые особи могут нереститься во время сексуальной привязанности, не связанной со слиянием тканей. У самцов этих видов также есть зубчатые челюсти, которые намного эффективнее при охоте, чем у симбиотических видов.
Половой симбиоз может быть необязательной стратегией у некоторых видов удильщиков. У Oneirodidae самки, у которых ещё не были развиты полностью функциональные гонады, имели прикреплённых симбиотических самцов, были зарегистрированы у представителей родов Leptacanthichthys и Bertella. Одна теория предполагает, если самка не является половозрелой, самцы прикрепляются к ним независимо от их собственного репродуктивного развития, но когда и самец, и самка становятся зрелыми, они нерестятся, а затем разделяются.
Одно из объяснений эволюции полового симбиоза состоит в том, что относительно низкая плотность самок в глубоководной среде оставляет мало возможностей для выбора партнера. Самки остаются крупными, чтобы увеличить плодовитость, о чём свидетельствуют их крупные яичники и икра. Ожидается, что самцы уменьшатся в размерах, чтобы снизить метаболические затраты в условиях ограниченных ресурсов и разовьют узкоспециализированные способности к поиску самок. Если самцу удается найти самку, то симбиотическая привязанность, в конечном счёте, с большей вероятностью улучшит жизнеспособность на протяжении всей жизни по сравнению со свободной жизнью, особенно когда шансы найти будущих помощников невелики. Дополнительным преимуществом симбиоза является то, что сперму самца можно использовать для множественных оплодотворений, так как он всегда остается доступным для спаривания. Более высокая плотность встреч самцов и самок может коррелировать с видами, которые демонстрируют факультативный симбиоз или просто используют более традиционное временное контактное спаривание.
Нерестовая кладка удильщиков рода Lophius состоит из тонкой полосы прозрачного студенистого материала шириной 25 см и длиной более 10 м. Икринки на этой полосе находятся в одном слое, каждое в своей полости. Такие икряные кладки среди рыб встречается редко. Личинки свободно плавают, их брюшные плавники вытянуты в нити.
Виды Lophius северо-западной Европы внесены в список ICES как «находящиеся вне безопасных биологических пределов». Кроме того, известно, что удильщикообразных иногда выносит на поверхность во время Эль-Ниньо, оставляя большие скопления плавающих мертвых рыб.
Семейство Lophiidae представляет коммерческий интерес в водах Западной Европы, восточной части Северной Америки, Африки и Восточной Азии. В Европе и Северной Америке мясо хвоста рыб из рода Lophius, известного как морской чёрт, широко используется в кулинарии, и его часто сравнивают с хвостом омара по вкусу и текстуре. В Азии, особенно в Японии, печень морского чёрта, известная как анкимо (ankimo), считается деликатесом. Удильщик особенно интенсивно потребляется в Южной Корее, где он является основным ингредиентом таких блюд, как Агуджим (Agujjim).
Рыбы открытого океана. Н.В.Парин 1998 г.
Еще одна весьма примечательная группа макропланктонных рыб — церациевидные или глубоководные удильщики (подотряд Ceratioidei), очень большой вклад в изучение которых внесен Э. Бертельсоном и Т. Питчем, весьма характерна для бати- и абиссопелагиали. Сюда относятся 11 семейств и около 120 видов, довольно разнообразных по размерам и внешнему облику (рис. 23), но обладающих одной общей особенностью — видоизмененным в «удилище» (иллиций) первым лучом спинного плавника. На конце иллиция находится «приманка» (эска), которая у разных видов сильно различается по строению, величине и форме. Все удильщики имеют довольно короткое тело (исключение составляет семейство Gigantactinidae) с непропорционально большой головой, огромный, как правило, зубастый рот и сильно растяжимый желудок. Окраска однотонная черная, реже темно-коричневая. Органов свечения на туловище и голове нет, и у большинства видов единственной светоизлучающей структурой служит железа, входящая в состав эски. Исключение составляют только линофрины (род Linophryne), у которых в дополнение к удочке имеются подбородочные усики со светящимися придатками. В заполненном слизью просвете эскальной трубчатой железы живут светящиеся симбиотические бактерии, которых до сих пор не удалось размножить в культуре и изучить детально. Эскальная железа и ее придатки (иногда сложно устроенные) изолированы от наружной среды отражающим и пигментным слоями, прободаемыми «окнами» и «щелями», которые обеспечивают видоспецифичность иллюминации эски. Регуляция интенсивности свечения обеспечивается только изменением интенсивности кровоснабжения, т. е. скорости подачи в железу кислорода, необходимого для жизнедеятельности бактерий. Функциональная роль эски как приманки, привлекающей кормовые объекты удильщиков, совершенно очевидна, однако серьезные межвидовые различия в строении органа заставляют предполагать и его коммуникационное значение.
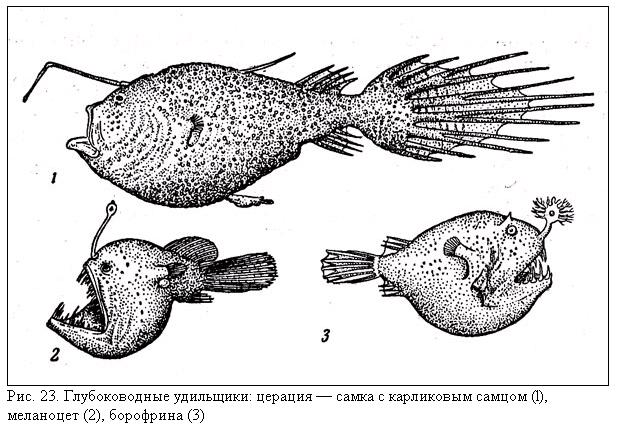
Взрослые удильщики наиболее обычны на глубине 1500—2500 м. Они известны, впрочем, и из более глубоких горизонтов, хотя их личинки проходят развитие в приповерхностных горизонтах (туда всплывает имеющая положительную плавучесть икра) и лишь по мере роста опускаются все глубже и глубже. Возрастные изменения горизонта обитания удильщиков представляют собой один из классических примеров вертикальных онтогенетических миграций. В глубинах океана образ жизни разнополых удильщиков имеет очень мало общего. Половозрелые самки, к которым, собственно говоря, и относятся все приведенные выше сведения об удильном аппарате, как правило, имеют длину от 5—8 до 20 см, но у гигантактиса (Gigantactis gargantua) она достигает 41 см, криптопсара (Cryptopsaras couesi) — 44 см, гимантолофа (Himantolophus groenlandicus) — 60 см, а у церации (Сеrаtias holboelli) — даже 145 см. Их пищу составляют различные глубоководные рыбы (миктофиды, топорики, меламфаи), иногда настолько крупные, что попытка проглотить их приводит к гибели самого хищника. Совсем недавно, например, на поверхности океана была обнаружена самка дицерации (Diceratias bispinosus) длиной 11 см; подавившаяся 37-сантиметровым долгохвостом (по заглоченному ею экземпляру был, кстати говоря, описан новый вид Ventrifossa johnboborum).
Самцы всех видов удильщиков отличаются от самок отсутствием иллициума, намного меньшими размерами, более прогонистым телом, сильным развитием обонятельных органов, наличием своеобразных вторичных зубов, сидящих на внешней стороне челюстей, направленных вперед и действующих как щипчики. В большинстве семейств они, несмотря на свою карликовость (длина 15—25 мм), ведут вполне активную самостоятельную жизнь, питаясь мелкими планктонными рачками и отыскивая самок по запаху. В то же время представители четырех семейств — Caulophrynidae, Ceratiidae, Linophrynidae и Neoceratiidae — демонстрируют совершенно уникальные отношения между полами, не известные ни у каких других позвоночных животных. Встретив самку своего вида, самцы этих удильщиков сначала накрепко вцепляются в нее зубами, а затем полностью срастаются с ее телом, теряя ненужные теперь органы (челюсти, глаза, кишечник), подключаясь к ее кровеносной системе и превращаясь в результате этих преобразований в самых настоящих паразитов, выполняющих только одну функцию — продуцирование спермы. Как правило, различия в размерах самок и паразитических самцов очень впечатляющи: так, на крупной церации длиной 119 см и массой 7 кг был обнаружен самец длиной 16 мм и массой всего 14 мг (по-видимому, он лишь недавно обрел свою «хозяйку», так как у других особей того же вида самцы-паразиты вырастают за счет самки до длины 16 см). У криптопсара известен, однако, случай прикрепления 10-миллиметрового самца к совсем юной, еще не половозрелой самочке длиной 15,5 мм, находившейся, по примерной оценке, в возрасте между 6 и 12 месяцами. Наибольший возраст крупной самки удильщика гигантактиса определен в 18 лет.
Видовое разнообразие батиабиссопелагического ихтиоцена значительно меньше, чем мезопелагического. Здесь отмечено, впрочем, не так уж мало видов — почти 200, которые, как уже указывалось, все относятся к макропланктону. Для этой зоны особенно характерны рыбы-удильщики подотряда Ceratioidei (около 100 видов, т. е. половина видового состава ихтиоцена). Типичными обитателями глубинных однородных вод являются также «черные» циклотоны, миктофиды рода Taaningichthys, китовидки, глубоководные угреобразные.
Икра и ранние личинки самых разных рыб глубоководной пелагиали — светящихся анчоусов, циклотон, удильщиков — проходят развитие в поверхностных горизонтах, но размножающихся особей в эпипелагиали никогда не наблюдали. Логично заключить, что нерест происходит на глубине обычного обитания взрослых рыб, откуда оплодотворенные икринки, обладающие положительной плавучестью, быстро всплывают. Вылупившиеся личинки по мере развития и роста постепенно опускаются все глубже и глубже.
Насколько сейчас известно, почти все глубоководные пелагические рыбы относятся к порционно нерестующим. Число икринок в порции подвержено довольно сильной изменчивости. В яичниках «светлых» циклотон насчитывали 0,1—0,9 тыс. ооцитов, «черных» циклотон — около 2—10 тыс., угря Derichthys serpentinus — 4,1 тыс., рыб-удильщиков, как правило, 7,5—10,8 тыс., но у очень крупной (65 см) самки цератии (Ceratias holboelli) — почти 5 млн. ооцитов. Суммарная абсолютная плодовитость у рыб мезо- и батипелагиали в общем невелика и заметно уступает плодовитости эпипелагических рыб, которые, впрочем, имеют гораздо более крупные средние размеры. Относительная плодовитость (число икринок на 1 г массы тела) у глубоководных видов также достоверно меньше, чем у приповерхностных.
Размножение глубоководных пелагических рыб в тропических водах, по всей вероятности, продолжается в течение всего года, но у краев тепловодной зоны либо имеет пик в летне-осеннее время, либо вообще ограничено этим сезоном. В то же время в субполярных водах существуют как зимненерестующие, так и летненерестующие виды.
Некоторые из упоминавшихся специфических особенностей батипелагических рыб, связанные с их размножением, в частности, половой диморфизм по размерам (и крайнее его проявление — самцовый паразитизм) и по строению органов обоняния, а также синхронный гермафродитизм, обеспечивающий возможность нереста любой пары случайно встретившихся производителей, несомненно, объясняются трудностями отыскания партнеров в условиях разреженного существования.
Преимущественными ихтиофагами являются и батиабиссопелагические удильщики, которые также подманивают добычу светящимися эскальными приманками и в некоторых случаях светящимися придатками усиков.
Следующий отряд — удильщикообразные — характерен преимущественно для мезобентали, в которой он представлен тремя семействами и примерно сотней видов. Большинство удильщиков (сем. Lophiidae), которых называют также морскими чертями, обитает на глубине до 500—600 м, но некоторые, например европейский морской черт (Lophius piscatorius), могут опускаться гораздо глубже - даже до 2000 м. В том же диапазоне глубин (200—500 м) обычны другие «удящие» рыбы, также относящиеся к числу бентических — жабовидные хаунаксы (сем. Chaunacidae) длиной до 40 см и морские нетопыри (Ogcocephalidae), редко превышающие 15—20 см в длину.
Рыбы Японского моря и сопредельных частей Охотского и Желтого морей. Часть 7. (Dactylopteriformes — Pegasiformes). Г.У.Линдберг, В.В.Федоров, З.В.Красюкова 1997 г.
XXXVI. Отряд Lophiiformes (Pediculati) — Удильщикообразные
Колючий спинной плавник у наиболее примитивных форм из 6 колючих лучей. Из них передние 3 находятся на черепе и первый луч видоизменены в удочку (иллициум) с приманкой (эска) на дистальном конце. Голова сильно уплощена впереди, в связи с чем отмечаются многочисленные случаи специализации: потеря носовых костей, затылочной комиссуры в сейсмосенсорной системе головы, изменения в соответствии со специализацией мускулатуры и иннервации. Epiotici отдельны от parietalia и соединены посередине назад до supraoccipitale. Жаберные отверстия в виде небольшого удлиненного, трубковидного отверстия, расположенного сразу же выше, позади или книзу (очень редко и частично впереди) основания грудного плавника. Urale 2 слито с urale I и praeurale I, и с hypuralia образует единую пластинку (иногда с глубокой щелью позади). Нижние radiale в грудном плавнике значительно удлинено кзади. Икра выметывается в двойной спиралевидной кожистой оболочке (Pietsch, Grobecker, 1987 : 268—269).
Широко распространены в Мировом океане, преимущественно в тропических и субтропических водах, обитатели шельфа, батиали, абиссали, эпи-, мезо-, бати- и абиссопелагиали. 5 подотрядов, 18 семейств , 63 рода и около 283 видов. В Японском море 4 подотряда, 4 семейства.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ПОДОТРЯДОВ И СЕМЕЙСТВ ОТРЯДА LOPHIIFORMES
(Lophiiformes Pietsch, Grobecker, 1987 : 273, с изменениями; Ceratioidei — Bertelsen, 1986 : 1371.)
1 (2). Колючий спинной плавник позади головы имеет 1—3 колючки. Верхнеглоточные зубы на pharyngobranchiale IV имеются. Cleithrum с заметным задним шипом. Подкрышечная кость с большим дополнительным отростком, присоединенным к переднему краю крышечной кости. Ложножабра хорошо развита. Икра и личинки большие; голова личинок по сравнению с телом небольшая (1. Подотряд Lophioidei). CCXV. Сем. Lophiidae
2 (1). Лучи колючего спинного плавника позади головы отсутствуют. Верхне глоточные зубы на pharyngobranchiale IV отсутствуют. Шип на cleithrum отсутствует. Дополнительный отросток подкрышечной кости отсутствует или редуцирован до небольшого выступа. Ложножабра обычно редуцирована или отсутствует. Икра и личинки небольшие; голова личинок по сравнению с телом большая.
3 (10). На черепе сверху 3 луча колючего спинного плавника. Птеригиофор удочки и 3-го луча колючего спинного плавника с сильно сжатым бритвообразным верхним выступом. Interhyale со срединным, направленным назад и в стороны отростком, который входит в контакт с предкрышечной костью. Межкрышечная кость плоская и широкая (2. Подоотряд Antennarioidei).
4 (5). В грудном плавнике 2 радиалии. V I 4. Parietalia слиты по средней линии и с supraoccipitale. Ectopterygoideum овальный по форме или отсутствует. Ceratobranchialia на I—III жаберных дугах с одной или больше зубными пластинками. Hypobranchiale II простое, hypobranchiale III отсутствует. [Сем. Brachionichthyidae]
5 (4). В грудном плавнике 3 радиали. V I 5. Parietalia хорошо отделены друг от друга supraoccipitale. Ectopterygoideum с 3 радиусами, верхний отросток этой кости прикрывает среднюю поверхность metapterygoideum. Ceratobranchialia на I—IV жаберных дугах без зубных пластинок. Hypobranchiale II и hypobranchiale III вильчатые.
6 (7). Сошник широкий, без заднего срединного отростка. Расстояние между наружными краями ethmoidalia lateralia почти такой же величины, как и между боковыми краями sphenotici. Верх головы широкий, ее ширина равна или превышает ширину metapterygoideum. Задний конец предчелюстной кости острый. Крышечная кость хорошо развита, простирается кзади. Pharyngobranchiale I и epibranchiale I с зубными площадками. Между вершинами остистых отростков гемальных дуг с 14-го по 16-й позвонки имеется костное соединение. Птеригиофор удочки (иллициум) удлиненный, сжат с боков, задний конец расширяется в стороны. [Сем. Lophichthyidae]
7 (6). Сошник узкий, с задним срединным отростком. Расстояние между наружными краями etmoidalia lateralia значительно меньше, чем между боковыми краями sphenotici. Верх головы узкий, ее ширина меньше ширины metapterygoideum. Задний конец предчелюстной кости лопатовидный. Крышечная кость редуцирована. Pharyngobranchiale I и epibranchiale I без зубных площадок. Костные соединения между вершинами остистых отростков гемальных дуг отсутствуют. Птеригиофор удочки короткий, его задний конец цилиндрический.
8 (9). Глаза на голове расположены сверху. Лучи колючего спинного плавника редуцированы. Рот маленький. Грудной плавник с 2 лопастями. Верхний луч нижней лопасти прикреплен кожистой перепонкой к боку тела, верхняя лопасть прикреплена перепонкой к лучам брюшного плавника. В мягкой части спинного плавника 16—17 лучей; в анальном плавнике 11—12. [Сем. Tetrabrachiidae]
9 (8). Глаз на голове расположен сбоку. Лучи колючего спинного плавника хорошо развиты. Рот большой. Грудной плавник не имеет лопасти, не прикреплен перепонкой к бокам тела или лучам брюшного плавника. В мягкой части спинного плавника 11—15 лучей, в анальном плавнике 6—9. CCXVI. Сем. Antennariidae
10 (3). На черепе сверху 2 или 3 колючих луча спинного плавника, но только передний из них приподнят над верхним профилем головы (2-й и 3-й лучи редуцированы и прикрыты кожей или отсутствуют). Птеригиофор удочки и 3-го колючего луча спинного плавника, если он имеется, без бритвообразного верхнего выступа. Interhyale без срединного, направленного назад и в стороны, отростка. Межкрышечная кость удлиненная и широкая.
11 (12). Второй колючий луч спинного плавника удлиненный. Третий колючий луч и его птеригиофор имеются. Epibranchiale I со срединным отростком, прикрепленным связкой к проксимальному концу epibranchiale II (3. Подотряд Chaunacoidei). CCXVII. Сем. Chaunacidae
12 (11). Второй колючий луч спинного плавника редуцирован до остатка под кожей головы или потерян; птеригиофор сразу же позади основания удочки, если она имеется. Третий луч и его птеригиофор отсутствуют. Epibranchiale I простое, без связки с epibranchiale II.
13 (14). Нижнеглоточные зубы имеются (ceratobranchiale V с зубами, кость имеет проксимальное расширение). Брюшной плавник имеется. Половой диморфизм не проявляется, самцы не паразитируют на самках (4. Подотряд Ogcocephaloidei). CCV1II. Сем. Ogcocephalidae
14 (13). Нижнеглоточные зубы отсутствуют (ceratobranchiale V без зубов, кость редуцирована до тонкого, палочковидного элемента). Брюшной плавник отсутствует (исключая личинки Caulophrynidae). Половой диморфизм сильно развит, у небольшой части таксонов самцы паразитируют на самках. [Подотряд Ceratoidei].
A (15—32). ТАБЛИЦА ОПРЕДЕЛЕНИЯ СЕМЕЙСТВ ПО САМКАМ
15 (16). Утолщение приманки (эски) отсутствует, удочка (иллициум) оканчивается длинными отростками. Наибольший луч спинного и анального плавников превышает 60 % SL. [Сем. Caulophrynidae]
16 (15). На вершине удочки имеется приманка в виде утолщения со светящимся органом. Наибольший луч спинного и анального плавников меньше 60 % SL.
17 (18). В спинном плавнике более 11 лучей. [Сем. Melanocetidae]
18 (17). В спинном плавнике менее 11 лучей.
19 (20). 2—3 передних луча спинного плавника имеют утолщение на дистальном конце с внутренними фотофорами. Ротовая щель вертикальная или очень косая. [Сем. Ceratiidae]
20 (19). Утолщенных лучей спинного плавника нет. Ротовая щель почти горизонтальная.
21 (22). На голове имеется второй колючий луч спинного плавника с дистальным фотофором (обычно булавовидный, но у крупных экземпляров некоторых видов скрыт под кожей головы и проявляется как пора). [Сем. Diceratiidae]
22 (21). Второй колючий луч спинного плавника (или пора) на голове отсутствует.
23 (24). Верхняя челюсть простирается далеко вперед нижней. Утолщение (эска) с 1—3 большими крючковидными зубчиками. [Сем. Thaumatichthyidae]
24 (23). Челюсти впереди на одном или почти на одном уровне. Утолщение без больших зубчиков.
25 (26). Удочка расположена на вершине рыла. Длина головы менее 35 % SL. Длина хвостового стебля более 20 % SL. [Сем. Gigantactinidae]
26 (25). Удочка расположена на голове позади вершины рыла. Длина головы более 35 % SL. Длина хвостового стебля менее 20 % SL.
27 (30). На теле в коже имеются колючки.
28 (29). Колючки в коже многочисленные, мелкие, сидящие близко друг к другу. [Сем. Centrophrynidae]
29 (28). Колючки в коже в виде больших относительно малочисленных костных пластинок со срединной заостренной вершиной. [Сем. Himantolophidae]
30 (27). Кожа на теле голая, без колючек.
31 (32). Жаберных лучей 5 (редко 4). D 3. А 3. [Сем. Linophrynidae]
32 (31). Жаберных лучей 6. В спинном и анальном плавниках более чем по 3 луча. [Сем. Oneirodidae]
Б (33—44). ТАБЛИЦА ОПРЕДЕЛЕНИЯ СЕМЕЙСТВ ПО САМЦАМ
33 (34). В анальном плавнике более 9 лучей. [Сем. Caulophrynidae]
34 (33). В анальном плавнике менее 9 лучей.
35 (36). В спинном плавнике более 11 лучей. [Сем. Melanocetidae]
36 (35). В спинном плавнике менее 11 лучей.
37 (38). Обонятельные органы очень малы. Глаза большие, шарообразные. [Сем. Ceratiidae]
38 (37). Обонятельные органы большие. Глаза не шарообразные.
39 (40). Глаза слегка трубчатые, направлены более или менее вперед. D 3. A 3. [Сем. Linophrynidae]
40 (39). Глаза боковые и не трубчатые. В спинном и анальном плавниках более чем по 3 луча (исключая Rhynchactis).
41 (42). Глаза маленькие, их диаметр менее 5 % SL. На челюстях зубы сильные, крючковидные: на верхней — 3, на нижней — 4. [Сем. Gigantactinidae]
42 (41). Диаметр глаза более 5 % SL. На верхней челюсти более 3 зубов.
43 (44). Кожа с шипиками. Передние ноздри по бокам головы. [Сем. Himantolophidae]
44 (43). Кожа голая, без шипиков. Передние ноздри направлены вперед и находятся около вершины рыла. [Сем. Oneirodidae]
Иллюстрированная энциклопедия рыб. Ст.Франк 1983 г.
Отряд Удильщикообразные (Lophiiformes)
У представителей отряда удильщикообразные (Lophiiformes) большая и расширенная передняя часть тела. Жаберные щели небольшие. На теле кожистые мясистые выросты, и рыбы напоминают разодранные пучки водорослей. Грудные плавники служат для передвижения по дну. Заостренная часть спинного плавника придвинута к голове или превращена в ряд самостоятельных шипов, либо вообще отсутствует. Эти рыбы живут в тропических, субтропических и умеренно теплых водах морей и океанов. Это малоподвижные, плохо плавающие рыбы, большей частью находящиеся на дне или в зарослях водорослей. Делятся на три подотряда и несколько семейств
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
ОТРЯД УДИЛЬЩИКООБРАЗНЫЕ (LOPHIIFORMES)
Первый луч колючего спинного
плавника сдвинут к верхней челюсти и
превращен в своеобразное «удилище»
(иллиний), несущее на конце «приманку»
(эска), которая служит для привлечения
добычи. Брюшные плавники,
если имеются, сидят на горле. Грудные
плавники поддерживаются 2—3
скелетными элементами, из которых нижний
сильно увеличен, обычно расширен на
конце и может совершать вращательные
движения. Это позволяет донным рыбам
использовать грудные плавники, как бы
согнутые в локте, для ползания по
грунту. Жаберные отверстия небольшие.
Тело голое, подчас с большим
количеством кожных выростов или покрыто
костными бугорками, шипиками или
бляшками. Все удильщикообразные — морские
хищные рыбы. Отряд удильщикообразных включает три подотряда — Удильщиковидных, Клоуновидных и
Глубоководных удильщиков с 16 семействами,
объединяющими свыше 70 родов и свыше
225 видов.
ПОДОТРЯД УДИЛЬЩИКОВИДНЫЕ (LOPHIOIDEI)
Удильщиковидные — это крупные
малоподвижные рыбы с большой уплощенной
головой, огромным ртом и огромным
желудком. Передний спинной плавник из
6 колючек, первые три колючих луча
обособлены друг от друга, а три задние,
меньшие, связаны перепонкой.
Грудные плавники большие.
Подотряд содержит одно семейство —
Удильщиковые.
ПОДОТРЯД КЛОУНОВИДНЫЕ (ANTENNARIOIDEI)
У клоуновидных рыб в первом
спинном плавнике имеется не более 3 колючих
лучей. Помимо того, они отличаются от
удильщиковидных формой тела и рядом
особенностей в строении скелета. Этот
подотряд включает 3—4 семейства,
важнейшие из них семейства клоуновых,
хаунаксовых и нетопыревых.
ПОДОТРЯД ЦЕРАЦИЕВИДНЫЕ, ИЛИ ГЛУБОКОВОДНЫЕ УДИЛЬЩИКИ (CERATIOIDEI)
Этот подотряд, включающий 11 семейств,
40 родов и около 120 видов, внешне
отличается от остальных удилщикообразных
прежде всего отсутствием брюшных
плавников. В то время как другие удильщикообразные обитают на дне или, в
немногих случаях, у поверхности,
глубоководные удильщики постоянно живут на
большой глубине, обитая в открытом
океане в толще воды. Замечательным образом
в этом подотряде проявляется половой
диморфизм, а в отдельных семействах,
и, по-видимому, независимо, выработался
паразитизм самцов.
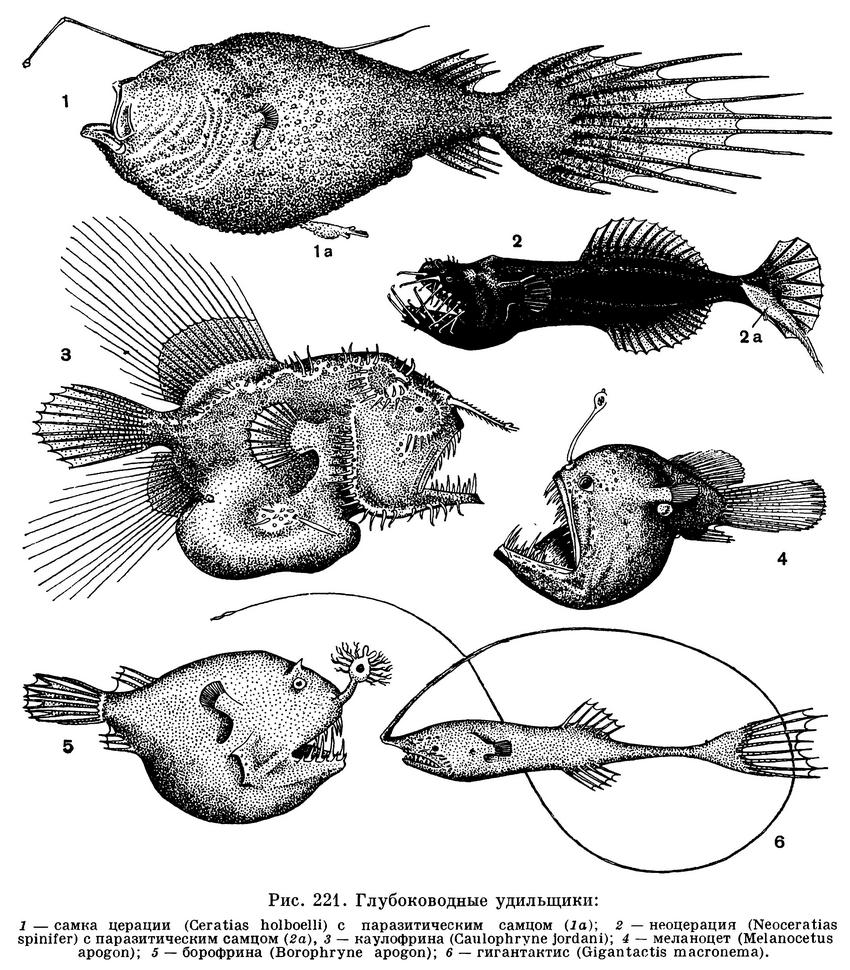
Несмотря на то что взрослые
глубоководные удильщики живут на глубинах,
куда не проникает свет и где отсутствуют
какие-либо сезонные изменения, в
Северной Атлантике, где они изучены наиболее
полно, все виды размножаются в весеннее
или летнее время. Нерест происходит,
по-видимому, на больших глубинах.
Развивающиеся икринки постепенно
поднимаются кверху, и личинки длиной 2—3 мм
выклевываются в приповерхностном слое
30—200 м, где питаются
преимущественно веслоногими рачками и планктонными
щетинкочелюстными червями (Chaetognatha). К началу метаморфоза молодь
успевает опуститься на глубину свыше 1000 м.
По-видимому, ее погружение совершается
быстро, так как самки на стадии
метаморфоза встречаются в слое 2—2,5 тыс. м,
а самцы на этой же стадии — на
глубине 2 тыс. м. В слое 1500—2000 м
обитают оба пола, прошедшие метаморфоз
и достигшие половозрелости, но иногда
взрослые особи встречаются и на
меньших глубинах. Взрослые самки питаются
в основном глубоководными батипелагическими рыбами (миктофовыми, гоностановыми, хаулиодами, топориками, меламфаями и др.), ракообразными и
реже головоногими, а взрослые самцы,
подобно личинкам, — веслоногими
рачками и щетинкочелюстными. Связанные с
индивидуальным развитием вертикальные
миграции глубоководных удильщиков
имеют очень большое приспособительное
значение, так как только в
приповерхностном слое их малоподвижные и
многочисленные личинки могут найти достаточно корма, чтобы накопить запасы для
предстоящего метаморфоза. Огромные
потери вследствие поедания икры и
личинок хищниками компенсируются
у удильщиков очень большой
плодовитостью. Их икра мелкая (не более 0,5—0,7 мм в диаметре). Их прозрачные
личинки напоминают крошечные
баллончики, благодаря тому что они одеты в
кожный чехол, раздутый студенистой тканью.
Эта ткань увеличивает плавучесть и
размеры личинок, что наряду с
прозрачностью оберегает их прежде всего от
мелких хищников, наиболее опасных для
них своей многочисленностью.
Все глубоководные удильщики
характеризуются очень резко выраженным
половым диморфизмом. Уже на самых
ранних личиночных стадиях, когда по форме
тела, размерам, степени развития глаз
и обонятельных органов оба пола вплоть
до стадии метаморфоза сходны между
собой, самки отличаются от самцов
наличием «удочки» (иллиция), в которую
превращен измененный первый луч спинного
плавника.
Во время и особенно после
метаморфоза у самок относительная величина
головы и рта сильно увеличивается,
обонятельные органы далее не
развиваются, глаза, как правило, становятся
маленькими, а у наиболее крупных видов
почти вовсе дегенерируют. У самцов,
напротив, тело становится более прогонистым, относительная величина головы
и челюстей сильно уменьшается, глаза
остаются крупными, а обонятельные
органы достигают очень больших размеров.
У самок, прошедших метаморфоз, на
челюстях, сошнике и верхнеглоточных
костях развиваются тонкие острые зубы,
своими загнутыми вершинами
направленные кзади и нередко способные
складываться внутрь. У многих видов они
мелкие и многочисленные, но у некоторых
челюстные зубы очень длинные (виды Linophryne, Lasiognathus и др.). У тауманихта (Thaumanichthys pagidostomus)
они особенно сильно развиты на верхней
челюсти, а у неоцерации (Neoceratias
spinifer) появляются дополнительные
огромные зубы, сидящие прямо на рыле
и снаружи нижней челюсти.
У самцов, прошедших метаморфоз,
вместо личиночных зубов в передней
части рта развиваются особые зубы, не
связанные с челюстями и сливающиеся
своими основаниями. Они действуют как
щипчики для захвата мелкой добычи
и для прикрепления к самкам.
Пожалуй, наиболее ярко половой
диморфизм проявляется в размерах. Все
свободноживущие самцы достигают в
длину не более 16—46 мм. Самки много
крупнее. Хотя у большинства видов они
не превышают в длину 5—10 см, у
нескольких видов самки достигают 20 см,
у криптопсара (Cryptopsaras couesi) до
44 см, а у гимантолофа (Himantolophus
groenlandicus) — до 60 см и веса 4 кг.
Самый крупный самец у гимантолофа
имеет длину всего 46 мм и вес 0,82 г,
т. е. по длине он меньше самки в 13
раз, а по весу в 5000 раз. Однако
крупнее всех самки церации (Ceratias
holboelli), достигающие длины свыше 1 м.
В четырех семействах глубоководных
удильщиков (Caulophrynidae, Ceratiidae,
Neoceratiidae и Linophrynidae) существуют
совершенно необычные отношения между
полами, не встречающиеся среди других
рыб и неизвестные для позвоночных
животных. Они выражаются в том, что
карликовые самцы живут в виде паразитов
на теле самок. Самец внедряется в кожу
самки острыми щипчикообразными
зубами. Вскоре губы и язык такого самца
полностью сливаются с телом самки, а его
челюсти, зубы, глаза и кишечник
постепенно редуцируются так, что в конце
концов он превращается в придаток,
вырабатывающий сперму. Питание
паразитирующего самца осуществляется за
счет крови самки, так как ее
кровеносные сосуды соединяются с сосудами
самца. Но жабры самца и крошечные
жаберные отверстия сохраняются, вследствие
чего поступающая к нему кровь может
обогащаться кислородом. Различия
в размерах самки и паразитирующего на
ней самца могут быть огромными. Так,
например, у самки церации (Ceratias
holboelli) длиной 119 см и весом 7 кг
прикрепившийся к ней самец имел длину
16 мм и весил всего 14 мг. Правда,
паразитирующий самец, питающийся за
счет самки, у этого вида может достигать
в длину 16 см. У остальных 7 видов, у
которых обнаружен паразитизм самцов (Caulophryne jordani, Neoceratias spinifer,
Cryptopsaras couesi, Edriolychnus schmidti, Photocorinus spiniceps, Borophryne
apogon и Linophryne argyresca), после
прикрепления к самкам самцы,
по-видимому, уже не увеличиваются в размерах
и не превышают в длину 14—22 мм.
Паразитирующие самцы
прикрепляются только ко взрослым самкам и лишь
после того, как они сами завершат стадию
метаморфоза. На одной и той же самке
на разных участках ее тела может
одновременно прикрепляться до трех самцов.
Однажды прикрепившись, такие самцы
навсегда утрачивают самостоятельность
и, судя по всему, в течение ряда лет
участвуют в размножении. До перехода
к паразитизму такие самцы обладают
хорошо развитыми глазами и очень
крупными обонятельными органами. Это
позволяет предполагать, что они отыскивают
самок по специфическому запаху, следы
которого сохраняются долгое время
в практически неподвижной воде больших
глубин. Приблизившись к самке, самец,
по-видимому, может визуально
«уточнить» ее принадлежность к своему виду
по строению эски, освещаемой
вспышками ее светящегося органа, или же по
цвету и частоте самих вспышек,
закодированных соответствующим образом.
Самцы остальных шести семейств
глубоководных удильщиков, по-видимому, ведут
свободный образ жизни, но не исключена
вероятность, что и они в период нереста
временно прикрепляются к самкам.
Только у самок глубоководных
удильщиков сохраняется «удилище» (иллиций)
со светящейся «приманкой» (эской),
которая у различных видов сильно
варьирует по форме и величине и бывает
снабжена самыми разнообразными кожными
придатками. У самок большинства видов
иллиций короткий, но у особей
некоторых родов (Rhynchactis, Lasiognathus
и Gigantactis) он очень длинный. Так,
у гигантактиса (Gigantactis macronema)
иллиций в 4 раза превышает длину тела.
У самок
некоторых семейств (Diceratiidae и Сеratiidae) имеется дополнительный
булавовидный светящийся орган на втором
головном луче, причем у самок церациевых, кроме того, перед мягким
спинным плавником есть 1—3 подобных
образования. В семействе линофриновых у
самок имеются особые придатки на
подбородке. Возможно, что у линофрины
(Linophryne arborifera) огромный
древовидный подбородочный придаток тоже
несет светящиеся железы.
Взрослые удильщики окрашены в темно-коричневый или черный цвет, их тело
обычно голое, и преобразованные чешуи
имеются лишь у отдельных видов. Так, у
тауманихта (Thaumanichthys pagidostomus) густо сидящие шипики имеются
на брюшной стороне тела, а у гимантолофов (Himantolophus) на теле имеются
костные бляшки, подчас очень крупные.
Желудок самок глубоководных
удильщиков способен очень сильно
растягиваться, благодаря чему они могут
заглатывать и переваривать очень крупную
добычу, нередко превосходящую их по
своим размерам. Жадность удильщиков
подчас приводит к гибели не только их
жертву, но и их самих. Иногда на
поверхности находят мертвых удильщиков
с заглоченной рыбой, превышающей их по
размерам более чем в два раза. Захватив
столь крупную добычу, удильщик не
может ее выпустить благодаря строению
своих зубов, и ему ничего не остается,
как продолжать заглатывать рыбу,
стремящуюся освободиться из капкана и
увлекающую его вверх. К тому времени,
когда удильщик заканчивает обед, он
погибает, по-видимому, от потери сил.
Личинки глубоководных удильщиков
встречаются лишь в тропической и
умеренно теплой зонах Мирового океана,
лежащих между 40° с. ш. и 35° ю. ш. и
ограниченных летними изотермами 20° С
в поверхностных водах. В более высоких
широтах, включая субарктические и
субантарктические воды, встречаются только
взрослые особи, которые попадают туда
благодаря выносу их течениями.
Обитатели бездны. Уильям Кроми 1971 г.
РЫБЫ-РЫБОЛОВЫ
Может случиться, что в темных глубинах рыба-топорик, бросившись на предмет, показавшийся ей сочной копеподой или светящейся креветкой, сама окажется влекомой в темную, жуткую пропасть. Если маленькая рыбка начинает вырываться, спасая свою жизнь, острые, искривленные зубы выпрямляются, преграждая жертве выход. Затем меньшие зубы заталкивают добычу в эластичный желудок. Рыба-удильщик раздобыла себе очередной «кусок хлеба».
Одновременно смешные и свирепые, эти маленькие немыслимые существа добывают себе пропитание рыбной ловлей, как заправские рыбаки. Будучи плохими пловцами, они висят неподвижно или лежат в засаде на дне океана на глубине от 900 до 3600 метров. Первый луч колючего спинного плавника у таких рыб, обособившись от остальных, удлинился и превратился в удилище, нависшее над головой рыбы. У одних видов удилище это короткое и толстое, у других — в несколько раз длиннее тела. У одного вида удильщика имеется «складное» удилище, которое может скользить по спине, подтягивая приманку все ближе и ближе к огромной пасти.
В результате поисков и ошибок эволюционного развития удильщики извлекли из кладовой природы целый арсенал экзотических приманок. Иногда они используют лишь мясистое утолщение на конце удочки, но чаще всего удочка разветвлена и напоминает вкусного червя или креветку. Для того чтобы жертва без труда могла отыскать приманку, почти у всех 80 видов удильщиков (за исключением двух) приманка снабжена «освещением». У одного вида светятся зубы. Ученые рыболовы с «Галатеи» извлекли на свет ранее неизвестную разновидность удильщика с раздвоенным светящимся органом, свешивающимся с неба. Доктор Бруун назвал черное, длиной в полметра широкоголовое существо «несомненно, самым удивительным трофеем экспедиции...» Ему дали название Galatheathauma axeli в честь экспедиционного судна и датского принца Акселя.
С помощью мышц удильщик забрасывает удочку и шевелит приманкой, имитируя некое движущееся существо. Когда жертва оказывается достаточно близко, нижняя часть пасти удильщика опускается вниз, челюсти выпячиваются, а жаберные крышки внезапно расширяются. Таким образом создается мощное течение, которое затягивает жертву в разинутую пасть хищника. Большинство таких удильщиков не больше человеческого кулака, но они могут проглотить добычу величиной с себя.
Сосредоточенные на глубине от 1500 до 1950 метров, глубоководные удильщики, должно быть, произошли от мелководных видов, как, например, морской черт (Lophis piscatorius). Такие удильщики в изобилии водятся среди скал и водорослей у берегов в тропических и полутропических водах, где они удят с помощью раздвоенной складки кожи, укрепленной на втягивающемся удилище. Самцы мелководных удильщиков имеют хорошее рыболовное оборудование: самцы же большинства глубоководных удильщиков предоставляют заниматься добычей пищи самкам. Достигнув брачного возраста, глубоководные самцы впиваются своими выпяченными зубами в брюхо, бока, лицевую и какую-либо другую часть самки. Постепенно губы и рот самца врастают в ее кожу, и все органы, кроме органов размножения, атрофируются. Системы кровообращения соединяются, и самец получает необходимое питание из крови самки. За это он оплодотворяет ее икру. Представляя собой немногим больше, чем наружный половой орган, самец продолжает расти, но достигает лишь небольшой части длины самки. Чем меньше самец, тем лучше, поскольку тем меньше потребуется пищи для того, чтобы супруги могли просуществовать.
Это необычный вид полового сожительства является приспособлением к условиям жизни в темноте, где рыбье «население» невелико и подходящую пару найти нелегко. Самцы-удильщики обладают удивительно острым нюхом, позволяющим им находить самок. Возможно, им помогает в этом и зрение: впиваясь во тьму крупными выпуклыми глазами, они отыскивают вспышки света, излучаемые световыми органами будущей супруги. Глубоководные обитатели, вероятно, флиртуют, подмигивая огоньками, подобно светлякам на лугах, подающим световые сигналы в брачную пору.