(Gadiformes Goodrich, 1909) Отряд Гадиформес, Трескообразные, Order Gadiformes Goodrich, 1909 (Anacanthini, Codfishes) 10 семейств
Материал подготовил Фактистов И. Е., 2022 г. Сайт www.fishbiosystem.ru
Новый отряд Gadiformes впервые описал в 1909 году английский учёный Эдвин Гудрич (Эдвин Стефен Гудрич — Edwin Stephen Goodrich, 1868 — 1946, английский биолог, специалист в области сравнительной анатомии, эмбриологии, палеонтологии и в эволюционных теориях, получил образование в Школе изящных искусств Феликса Слейда (Slade School of Fine Art), после обучался в Университетском колледже Лондона (University College London), в 1894 году окончил с отличием Мертон-коллежд (Merton College) в Оксфорде, в 1905 году был избран членом Королевского общества, с 1921 по 1946 год преподавал на кафедре зоологии в Оксфордском университете (University of Oxford), в 1932 году награждён медалью Линнея — одной из высших наград Лондонского Линнеевского общества).
Гадиформес, Трескообразные (Gadiformes) — отряд морских и пресноводных (2 вида) лучепёрых рыб, обитающих по всему миру. Типовой вид — Gadus morhua Linnaeus, 1758. Молекулярные данные (Wiley et al., 2000; Miya et al., 2001, 2003, 2005) дают основания считать, что Gadiformes и Zeiformes являются сестринскими таксонами и образуют монофилетическую группу. Из 10 семейств этого отряда, включающих более 600 видов, в водах России представлены 7 семейств (см. Рыбы России), в том числе многие коммерчески важные виды — трески, макрурусы, хеки, минтаи, пикша и путассу.
Название отряда Gadiformes образовано от имени типового рода Gadus Linnaeus, 1758, название которого происходит от латинизации древнегреческого слова γαδος (gados) — «треска», введённое греческим оратором Афинеем (Ateneo — около 170 г.) для обозначения этой рыбы, которая в основном продавалась в финикийском городе Гадес (Gades). Окончание отрядов «–formes» происходит из латинского языка и означает «похожей формы». Английское название «codfishes» — «тресковые рыбы», соответствует русскому названиям отряда «трескообразные», и указывают на то, что к данному отряду относятся рыбы, похожие на треску. Раньше отряд трескообразных называли бесшипными, бесколючими, неколючими или мягкопёрыми — Anacanthini (anacanthine fishes), от гречкских слов αν- (an) — «без» и ακανθα (akanta) — «колючка», «шип», отмечая отсутствие колючих лучей на плавниках представителей этой группы рыб.
Время появления ранних трескообразных рыб было датировано поздним меловым периодом (роды †Rankinian — Ранкиниани, †Paratichthys — Паратихтисы). Хронология вымерших родов (см. диаграмму): начало эоцена — †Rhinocephalus (Риноцефалусы), †Trichurides (Трихуриды), †Palaeogadus (Палеогадусы); конец эоцена — †Eophycis (Эофицисы); начало олигоцена — †Pseudoraniceps (Псевдораницепсы), †Palaeomolva (Палеомольвы); начало миоцена — †Bregmacerinia (Брегмацеринии), †Trichiurichthys (Трихиурихтисы); середина миоцена — †Eclipes (Эклиресы), †Bolbocara (Болбокары).
Время появления некоторых современных родов отряда Трескообразные: начало эоцена — Urophycis (Урофицисы), Raniceps (Раницепсы), Nezumia (Незумии), Merluccius (Мерлуциусы), Melanonus (Меланонусы); середина эоцена — Bregmaceros (Брегмацеросы); конец эоцена — Tripterophycis (Триптерофицисы), Macruronus (Макруронусы), Coelorinchus (Целоринхусы), Bathygadus (Батигадусы); начало олигоцена — Trachyrincus (Трахиринкусы), Squalogadus (Сквалогадусы), Phycis (Фицисы), Hymenocephalus (Гименоцефалусы), Gaidropsarus (Гайдропсарусы), Gadiculus (Гадикулусы); середина олигоцена — Trisopterus (Тризоптерусы), Micromesistius (Микромезистиусы), Coryphaenoides (Корифеноидесы); начало миоцена — Ventrifossa (Вентрифоссы), Physiculus (Физикулюсы); середина миоцена — Molva (Мольвы), Lepidion (Лепидионы), Merlangius (Мерлангиусы), Laemonema (Лемонемы); конец миоцена — Gadus (Гадусы), Gadomus (Гадомусы), Brosme (Бросмы); начало плейстоцена — Pseudophycis (Псевдофицисы), Gadella (Гаделлы), Theragra (Терагры), Microgadus (Микрогадусы), Melanogrammus (Меланограммусы); конец плейстоцена — (Поллахиусы).
Ранние трескообразные рыбы были похожи по строению на ранних перкопсообразных (Percopsiformes), и почти все оставались морскими обитателями, впоследствии специализировавшиеся в различных средах. Они имели уменьшенный хвостовой скелет; удлинённое тело. Отличительные признаки современных трескообразных: непарные (спинные, анальные) и брюшные плавники не имеют колючих лучей (за исключением первых лучей спинного плавника у некоторых долгохвостов); если есть брюшные плавники, то они обычно находятся под грудными или впереди них (торакальные или югулярные), редко позади грудного плавника (у Macrouridae). Лучей брюшного плавника до 11. У большинства видов спинной и анальный плавники длинные.
Трескообразные рыбы характеризуются удлинённым, сужающимся к хвосту, более или менее конусообразным телом. Плавники лишены настоящих колючих лучей. Брюшные плавники, если они есть, находятся под или впереди грудных плавников (редко сзади, у макрурид) и имеют максимум 11 лучей. Один спинной плавник, очень длинный, или два или три смежных или отдельных спинных плавника. Один очень длинный анальный плавник или два смежных или отдельных плавника. Скелет хвостового плавника симметричный. Наблюдается редукция в заднем отделе позвоночника, приводящая к тому, что число задних птеригиофоров спинного и анального плавников превосходит число хвостовых позвонков. Очень характерен для многих трескообразных усик на подбородке. Плечевой пояс прикреплён к черепу. Межчелюстные и верхнечелюстные кости подвижны. Нижние глоточные кости отделены друг от друга. Предчелюстная кость (premaxilla) образует весь край верхней челюсти, выдвижная у некоторых таксонов. Наружная крыловидная кость (ectopterygoid) без зубов. Глазнично-клиновидная кость (orbitosphenoid) и основная клиновидная кость (basisphenoid) отсутствуют. Жабры гребёнчатые. Лучей жаберной перепонки — бранхиостегальных лучей (branchiostegal rays) — 6—8. Плавательный пузырь не имеет протока (не соединен с кишечником, закрытопузырные рыбы), у некоторых видов отсутствует (плавательный пузырь отсутствует у Melanonus и Squalogadus). Чешуя обычно циклоидная (гладкая), редко ктеноидная. Трескообразные рыбы обладают отолитом уникального строения. Однако у представителей семейства Phycidae отолит ещё более специализирован и отличается от такового у тресковых. По положению брюшных плавников трескообразные рыбы сходны с окунеобразными (Perciformes), по строению плавников — с карпообразными (Cypriniformes), от которых отличаются отсутствием воздушного протока, ведущего из плавательного пузыря. Размеры от 2 см (брегмацеры — род Bregmaceros Thompson, 1840) до 2 м (атлантическая треска — Gadus morhua Linnaeus, 1758).
Распространение (карта), варьируется в зависимости от семейства: морские воды по всему миру и редко пресные воды (налим — Lota lota). Подавляющее большинство видов (бенто- и батипелагические) холодноводные и придонные рыбы, обитающие главным образом в глубинах океана и в морях умеренных областей обоих полушарий или более холодным климатом (тропические виды обычно глубоководны). Некоторые виды могут заходить в эстуарии, но только налим, и еще некоторые популяции у одного вида, приурочены к пресной воде. Многие из представителей этого отряда осуществляют сезонные миграции, связанные с воспроизводством или поисками важных пищевых объектов. Наблюдения за фактическим нерестовым поведением немногочисленны, как и данных о различиях в их поведении днём и ночью. Кормление также меняется в зависимости от сезона для многих видов с характерной картиной, связанной с прекращением кормления в сезон нереста. Все виды плотоядны. Молодые особи ведут пелагический образ жизни, а взрослые особи — придонный образ жизни. Многие трескообразные рыбы питаются добычей, находящейся в субстрате, и им помогает в поисках пищи присутствие тактильных усиков на подбородке.
Больше известно о размножении и развитии икринок и личинок представителей семейств Gadidae и Merlucciidae, поскольку эти таксоны содержат коммерчески важные виды, которым уделяется наибольшее внимание. Трескообразные рыбы, как правило, выпускают массу икринок, которые затем оплодотворяются самцами. Почти у всех видов икра пелагическая (хотя о размножении глубоководных видов известно немного). Атлантическая треска (Gadus morhua) — одна из самых плодовитых рыб в мире. Самка весом 5 кг способна произвести 2,5 миллиона икринок, а более крупные самки могут произвести ещё больше. Не отстаёт и пикша (Melanogrammus aeglefinus). Самка длиной чуть меньше метра может произвести около 2 миллионов икринок. Размер икры трескообразных варьируются от примерно 0,5 мм в диаметре у некоторых Moridae и Phycidae до примерно 2,0 мм у некоторых Gadidae и Macrouridae. Хорион (внешняя оболочка) у большинства видов гладкий, но у Macrouridae может иметь шестиугольный узор. Большинство представителей Gadiformes имеют в икринке единственную небольшую жировую каплю, хотя икра гадид её не имеют. Стадии раннего периода жизни известны менее чем для трети видов, описанных в отряде, но некоторые описания доступны для каждого семейства. Личинки обладают разнообразной формой и специализацией. Согласно Fahay и Markle (1984), «похоже, не существует каких-либо уникальных или диагностических признаков для молодых Gadiformes. Особенности формы тела, морфологии ануса и развития брюшных плавников в сочетании с конкретными признаками семейства, представляются наиболее полезными для первоначальной идентификации. Трансформация постепенная и прямая, без заметных изменений в онтогенезе». Кишечник большинства личинок трескообразных рыб закручивается в начале онтогенеза и в сочетании с сужающейся задней частью и округлой головой, делает её по общему внешнему виду похожей на головастика. Это не было зарегистрировано во всех семействах трескообразных и не всегда легко наблюдается, но у очень молодых личинок анус выходит латерально (боковое расположение) через плавниковую складку, а не по её краю, как у большинства личинок рыб. Другой особенностью личинок трескообразных рыб является то, что некоторые вторичные хвостовые лучи развиваются раньше некоторых первичных (у видов, имеющих хвостовой плавник).
МСОП перечисляет три вида трескообразных рыб: Physiculus helenaensis отнесен к категории находящихся под угрозой исчезновения, а Gadus morhua и Melanogrammus aeglefinus отнесены к категории уязвимых. В Соединенных Штатах Америки нет никаких рыб, находящихся под угрозой исчезновения. Однако среди факторов, угрожающих устойчивости жизнеспособных популяций трескообразных рыб, высокие показатели перелова, особенно в отношении атлантической трески. В некоторых важных районах, например, в восточных провинциях Канады, треска коммерчески вымерла, а это означает, что численность её популяции настолько низка, что она больше не может поддерживать промысел. Утрата различных морских местообитаний, критически важных для выживания молодых рыб (а также более старых), часто упоминается как фактор, способствующий сокращению популяций рыб. Почти для всех рыб наши знания о важнейших функциях этих местообитаний недостаточны или поверхностны, и расширенные исследования в этих областях часто упоминаются как необходимые для надлежащего управления морскими ресурсами.
Многие основные виды пищевой рыбы находятся в этом отряде. К трескообразным, имеющим большое хозяйственное значение, относятся многочисленные виды, распространённые преимущественно в холодных морях. Тем не менее, Gadiformes не испытывают недостатка в тёплых и умеренных водах. Трескообразные имеют большое практическое значение: они дают 10—15% мирового улова, занимая второе место вслед за сельдеобразными. Например, в конце 1980-х годов около 15 101 665 тонн, что составляет 17% мирового вылова морских рыб. Сюда относятся много важных промысловых рыб, особенно из семейства тресковых (95%), за которыми следуют Merlucciidae, Macrouridae и Moridae.
Важно отметить, что среди ихтиологов отсутствует согласие относительно состава, происхождения, иерархии или отношений внутри Gadiformes. Нет согласия и в отношении внешних взаимоотношений членов этой, довольно произвольной, группы рыб. Не существует групп характеристик, которые можно было бы отнести только к Gadiformes; следовательно, синапоморфии (производные символы, общие для всех членов рассматриваемой группы) не были идентифицированы для всех членов отряда, за исключением нечленов.
Рыбы открытого океана. Н.В.Парин 1998 г.
Очень много донно-придонных океанических видов содержит отряд трескообразных рыб, две трети состава которого обитает на значительных глубинах. Преимущественно глубоководным является, в частности, семейство моровых (Moridae), впервые обособленное известным отечественным ихтиологом А. Н. Световидовым (1903—1985).
Атлас-определитель рыб: Книга для учащихся. Н.А.Мягков 1994 г.
Отряд Трескообразные (Gadiformes)
В основном морские рыбы. Ценнейшие объекты океанического промысла (в середине века их доля в европейской добыче составляла около 1/3). В наших водах обычны представители одного семейства.
Непарные и брюшные плавники без колючих лучей. Брюшные плавники лежат под грудными или даже впереди них. Есть 1—3 спинных плавника. У многих видов развит подбородочный усик, иногда несколько. За редким исключением (налим обыкновенный) морские рыбы.
Иллюстрированная энциклопедия рыб. Ст.Франк 1983 г.
Отряд Трескообразные (Gadiformes)
Представители отряда трескообразных (Gadiformes), закрытопузырьные рыбы, плавательный пузырь у них не связан с кишечником. Брюшные плавники расположены перед грудными. У плавников нет твердых лучей. Треска морская рыба амфибореального распространения. Согласно Бергу и Световидову, рыба эта не родственна с окуневидными, к которым раньше, причислялась. Она, видимо, возникла в тропических водах. Промысловое значение ее очень велико. Например, в 1936 - 37 годах ее годовой улов составил 2400 тыс. тонн, т. е.около 14 процентов всего мирового улова.
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
ОТРЯД ТРЕСКООБРАЗНЫЕ (GADIFORMES)
В отряде трескообразных объединены
такие рыбы, у которых непарные и
брюшные плавники не имеют колючих лучей
(за исключением первых лучей спинного
плавника у некоторых долгохвостов),
брюшные плавники находятся под
грудными или впереди них, плечевой пояс
прикреплен к черепу, скелет хвостового
плавника симметричный, плавательный пузырь
не соединен с кишечником. Отмечая
отсутствие колючих лучей в плавниках, трескообразных называли ранее
«неколючими» или «бесшипыми» — Anacanthini (от
греческого an — «без», acantha — «колючка,
шип»). Очень характерен для многих трескообразных усик на подбородке.
Трескообразные — морские,
преимущественно холодноводные и придонные рыбы,
распространенные главным образом в
глубинах океана и в морях умеренных
областей обоих полушарий. Этот отряд
включает четыре подотряда — паркетниковидных, тресковидных, долгохвостовидных
и ошибневидных, содержащих 10—11
семейств, всего около 700 видов. Свыше
половины из них глубоководные, к
которым относятся долгохвостовидные (около
300 видов), часть тресковидных (около
100 видов) и большая часть (свыше
100 видов) ошибневидных.
Трескообразные имеют большое практическое значение:
они дают 10—15% мирового улова,
занимая второе место вслед за
сельдеобразными. Свыше 9/10 этого улова дают тресковидные, остальное составляют
долгохвосты и ошибневидные.
ПОДОТРЯД ПАРКЕТНИКОВИДНЫЕ (MURAENOLEPOIDEI)
Семейство Паркетниковые (Muraenolepidae)
Паркетниковые представляют собой
своеобразное семейство трескообразных,
выделяемое в особый подотряд паркетниковидных.
ПОДОТРЯД ТРЕСКОВИДНЫЕ (GADOIDEI)
Подотряд содержит семейства брегмацеровых, тресковых, моровых и меланоновых.
ПОДОТРЯД ДОЛГОХВОСТОВИДНЫЕ (MACROUROIDEI)
Долгохвостовидные близки к тресковым.
Это, пожалуй, самая многочисленная
группа придонных глубоководных рыб,
насчитывающая до 300 видов, из которых
около 200 обитает в одном только
Тихом океане. Их ареал целиком
охватывает акватории Индийского,
Атлантического и Тихого океанов, причем в двух
последних он простирается от Арктики до
Антарктиды. Долгохвосты не обнаружены
лишь в Северном Ледовитом океане, чему
до сих пор нет еще удовлетворительного
объяснения. Подотряд представлен одним
семейством.
Рыбы Японского моря и сопредельных частей Охотского и Желтого морей. Часть 2. (Acipenseriformes — Polynemiformes). Г.У.Линдберг, М.И.Легеза 1965 г.
XIX. Отряд GADIFORMES
N. olfactorii не тянутся в глазничной полости по соседству с межглазничной перегородкой (как обычно у большинства Teleostei), а помещаются в канале, который лежит над межглазничной перегородкой и сообщается с черепной полостью; lobi olfactorii в непосредственном соседстве с носовыми капсулами (как у Galaxiiformes и у многих Cypriniformes); таким образом, мозг продолжается далеко вперед. Opisthoticum очень велико и разделяет prooticum от occipitale laterale; оно прободено отверстием для n. glossopharyngeus. Есть обособленный симметричный хвостовой плавник (псевдокавдальный). Брюшные плавники впереди грудных. Тазовой пояс соединен связкой с ключицами. Колючек в плавниках нет. Нет миодома, орбитосфеноида и базисфеноида. Нет мезокоракоида. Закрытопузырные. Чешуя циклоидная. В костях нет костных клеток. Sacculus очень велик. Первый позвонок прикреплен к черепу. Межмышечных косточек нет (верхние ребра имеются). Лопаточное отверстие между лопаткой и коракоидом. Главным образом морские. (Берг, 1940 : 283).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА СЕМЕЙСТВ ОТРЯДА GADIFORMES
(Сем. Muraenolepidae, распространенное в антарктических и субантарктических морях и представленное 1 родом, отличается от других семейств отряда Gadiformes отсутствием свободного хвостового плавника, полностью слитого со спинным и анальным, сильно удлиненными чешуйками, расположенными под углом друг к другу (по типу Anguilla), и большим числом, не менее 10 (10—13), радиалий в грудных плавниках. У остальных семейств этого отряда хвостовой плавник отделен от спинного и анального плавников, реже частично слит, а грудных радиалий 4—6.)
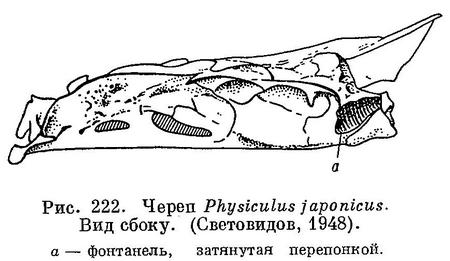
1 (2). 2 спинных плавника (У рыб из рассматриваемого района морских вод.) соприкасающихся основаниями, но разделенных глубокой выемкой; первый спинной короткий, начинается на спине позади жаберного отверстия; второй спинной очень длинный. Фонтанелли сзади черепа в occipitalia lateralia имеются; через них плавательный пузырь своими рожкообразными отростками соприкасается с ушными капсулами. Обнаружить это можно, как указывает А. Н. Световидов (1948 : 66), приподняв жаберную крышку и сделав над жабрами продольный разрез вдоль основания черепа; приподняв отросток можно увидеть фонтанелль (рис. 222). Глубоководные рыбы. LXV. Moridae.
2 (1). 3 спинных плавника, а если 2, то первый спинной расположен или на голове, или если на спине, то над жаберными отверстиями и состоит из 1 усикообразного луча и нескольких десятков коротких кожных выростов, скрытых в глубокой канавке; второй спинной длинный, отделен от первого заметным промежутком. Фонтанелли сзади черепа в occipitalia lateralia нет, и плавательный пузырь не соприкасается с ушными капсулами. Рыбы обычно умеренных глубин.

3 (4). Первый спинной плавник расположен на голове в области затылка и состоит лишь из 1 тонкого длинного луча, без дополнительных коротких кожистых лучей. Второй спинной плавник с очень глубокой выемкой и производит впечатление 2 плавников (рис. 228). LXVI. Bregmacerotidae.
4 (3). Первый спинной плавник расположен на спине и начинается заметно позади жаберных отверстий, а если над ними, то плавник особой формы; у него позади длинного, усикообразного луча имеется несколько десятков коротких кожных лучевидных выростов, скрытых в глубокой канавке. LXVI1. Gadidae.
XX. Отряд MACRURIFORMES
Как Gadiformes, но lobi olfactorii расположены у переднего мозга; n. olfactorii не входят в полость глазницы, проходя внутри перепончатой части межглазничной перегородки. У некоторых в первом спинном плавнике колючка; иногда на последнем неветвистом луче первого спинного плавника бывают фулькры. Чешуя ктеноидная или циклоидная. Radialia грудных плавников в числе 3—6. Хвостовой плавник слит со спинным и анальным, симметричный. Брюшные плавники под грудными или немного впереди их, с 5—17 лучами. Первый позвонок не прикреплен к черепу. Глубоководные рыбы. (Берг, 1940 : 286).
1 или 2 семейства.
Фауна СССР. Рыбы. Том IX, вып.4. Трескообразные. А.Н.Световидов 1948 г.
КРАТКИЙ ИСТОРИЧЕСКИЙ ОЧЕРК СИСТЕМЫ ОТРЯДА
В отряд Anacanthini J. Müller (1846 : 177), впервые установивший его, объединил рыб, имеющих плавники без колючих лучей, брюшные плавники, если они имеются, на горле или груди, плавательный пузырь, если он имеется, не соединенный с кишечником. Всю обширную группу рыб, объединяемую в один отряд по этим признакам, J. Müller делит следующим образом (l. с. : 202):
Ordo Anacanthini.
Subordo I. Anacanthini subbrachii.
Fam. 1. Gadoidei.
2. Pleuronectides.
Subordo II. Anacanthini apodes.
Fam. 1. Ophidini.
В таком объеме отряд Anacanthini, как указывает сам J. Müller (l. с. : 177, 199), состоит частично (Anacanthini subbrachii) из Malacopterygii subbrachii, частично (Anacanthini apodes) из Malacopterygii apodes системы Cuvier (1817 : 212, 229). По внутреннему строению J. Müller считает Anacanthini близкими к отряду Anacanthopterygii, с которыми они, точно так же как и с Pleuronectidae, сближались долгое время и впоследствии.
Günther (1862 : 317), принимая отряд Anacanthini в объеме, установленном J. Müller'ом, считает, что колючие лучи у них имеются только в вертикальных плавниках, и делит отряд на два подотряда несколько иначе:
Anacanthini Gadoidei. Голова симметричная. Семейства: Gadopsidae, Lycodidae, Gadidae, Ophidiidae (группы: Brotulina, Ophidiina, Fierasferina, Ammodytina, Congrogadina), Macruridae, Ateleopodidae.
Anacanthini Pleuronectoidei. Обе стороны головы несимметричны. Семейство: Pleuronectidae.
Система Günther'a отличается и большим подразделением внутри подотрядов.
Соре (1872 : 341) определяет Anacanthini как рыб, у которых основание черепа простое (без миодома), posttemporale вильчатое, лопаточное отверстие между scapula и coracoideum; pharyngialia три, горизонтальные, третья более длинная; лучи спинных плавников мягкие; хвостовой плавник дифицеркальный. По этим признакам он относит к Anacanthini, считаемым им за одну из групп отряда Percomorphi, только семейства Gadidae и Macruridae, выделяя большинство сближаемых с ними семейств в особую группу того же отряда Scyphobranchii. Камбал Соре выделяет в самостоятельный отряд Heterosomata, в качестве которого они приняты почти во всех более поздних системах. Система Anacanthini Соре является значительным шагом вперед, так как объем и многие положенные им в основу ее признаки приняты в большинстве более поздних систем.
Jordan и Gilbert (1883 : 783) в подстрочном примечании указывают, что в отряде Acanthopteri семейства Gadidae, Macruridae, Lycodidae, Congrogadidae, Fierasferidae и Ophidiidae образуют особую группу или подотряд Anacanthini или Jugulares, Отличаемый от других Acanthopteri длинными спинными и анальными плавниками, состоящими только из мягких лучей. Семейство Gadidae они разделяют на подсемейства Gadinae, Merlucciinae, Chiasmodontinae, Brotulinae, хотя последнее в определительных таблицах (р. 79) выделено ими в самостоятельное семейство.
Gill (1884 : 167) дает следующие подразделения Anacanthini или Jugulares:
Gadоidеа. Орбито-ростральная часть черепа длиннее, чем задняя, черепная полость широко открыта спереди, supraoccipitale хорошо развито и с килем сзади, лопаточное отверстие между scapula и coracoideum. Семейства: Gadidae, Merlucciidae, Bregmacerotidae, Ranicipitidae, Macruridae.
Ophidioidea. Орбито-ростральная часть черепа сжата и короче, чем задняя, черепная полость частично замкнута расширением и соединением parasphenoideum и frontalia, scapula с отверстием в середине. Семейства: Brotulidae, Brotulophididae, Ophidiidae, Fierasferidae, Congrogadidae.
Lycodoidea. Орбито-ростральная часть черепа сжата и короче, чем задняя, черепная полость спереди открыта, но ограничена с боков расширением соприкасающихся parasphenoideum и frontalia, supraoccipitale наклонно и крышевидно сзади, scapula с отверстием в середине. Семейства: Lycodidae, Ammodytidae, Ateleopodidae, Xenocephalidae,
Все группы семейства Ophidiidae и ряд родов семейств Gadidae, Ophidiidae и Ateleopodidae системы Günther'a Gill считает семействами; семейство Gadopsidae и род Chiasmodus он считает семействами Acanthopterygii. Весьма существенным в системе Gill'а является то, что в основе разделения отряда на группы им указаны новые остеологические признаки, имеющие значение и в настоящее время.
Значительно ближе к современному объему, взяв в основу признаки группы Gadoidea системы Gill'a, принимают Anacanthini Jordan и Evermann (1898 : 2528), считая их точно так же одним из подотрядов Acanthopterygii. К Anacanthini они относят семейства Gadidae, Merlucciidae, Macruridae. Семейство Ranicipitidae не включено в состав подотряда Anacanthini, вероятно, только потому, что его у берегов Сев. Америки нет. Семейства Bregmacerotidae, Brotulidae, Ophidiidae, Lycodidae, Fierasferidae и некоторые другие объединены в группу Ophidioidea, сближаемую с группой Blennioidea подотряда Xenopterygii из отряда Acanthopterygii. Камбалы являются особым подотрядом (Heterosomata) отряда Acanthopterygii, сближаемым с Anacanthini. Семейство Ammodytidae отнесено к подотряду Rhegnopteri, сближаемому с подотрядом Percesoces.
Boulenger (1901 : 268) объединяет тресковых вместе с семействами Trachinidae, Callionymidae и Nototheniidae в подотряд Jugulares, относимый им к отряду Acanthopterygii по общности положения у них лопаточного отверстия между scapula и coracoideum и брюшных плавников на горле. Эта система, как увидим ниже, признания не получила, так как сближать рыб в один подотряд только по положению брюшных плавников и лопаточного отверстия невозможно.
В 1903 г. Regan (1903 : 459) определяет Anacanthini следующим образом: parietalia отделены друг от друга посредством supraoccipitale; prooticum и occipitalia lateralia разделены увеличенным opisthoticum; плечевой пойс прикреплен к черепу; mesocoracoideum нет; непарные и брюшные плавники без колючих лучей (за исключением первого луча спинного плавника у некоторых Macruridae); брюшные плавники под грудными или впереди них; тазовый пояс позади ключичного симфизиса и прикреплен к нему лишь при помощи связки; жабры гребневидные; плавательный пузырь не соединен с кишечником.
Положению лопаточного отверстия между scapula и coracoideum Regan не придает существенного значения, так как оказалось, что у Gadomus longifilis из семейства Macruridae отверстие это находится в scapula. Точно так же и у Trematomus из семейства Nototheniidae, которое Boulenger по положению лопаточного отверстия между scapula и coracoideum сближал с семействами Anacanthini, отверстие это оказалось в scapula.
К Anacanthini Regan относит семейства Gadidae, Macruridae и Muraenolepidae. Роды Merluccius и Raniceps, выделяемые некоторыми в особые семейства, включены им в семейство Gadidae. К семейству Gadidae относит Regan и род Bregmaceros, у которого, по его исследованиям, кости брюшного пояса слабо прикреплены к плечевому поясу, хвостовой плавник симметричный и лопаточное отверстие находится между scapula и coracoideum, а не в scapula, как считали Jordan и Evermann (1898 : 2526), выделявшие Bregmaceros в самостоятельное семейство и сближавшие его с семействами Brotulidae и Ophidiidae.
В настоящее время система Anacanthini Regan'a является общепринятой. С небольшими изменениями принята она и Jordan'oм (1923 : 162).
СИСТЕМА
Goodrich, принимая в Vertebrate Craniata (1909 : 478) Anacanthini, называемых им Gadiformes, в объеме и определении Regan'a, обращает внимание на давно известную (Stannius, 1854 : 63), но забытую особенность строения их межглазничной области и передней части мозга, считая эту особенность наиболее характерным для них признаком.
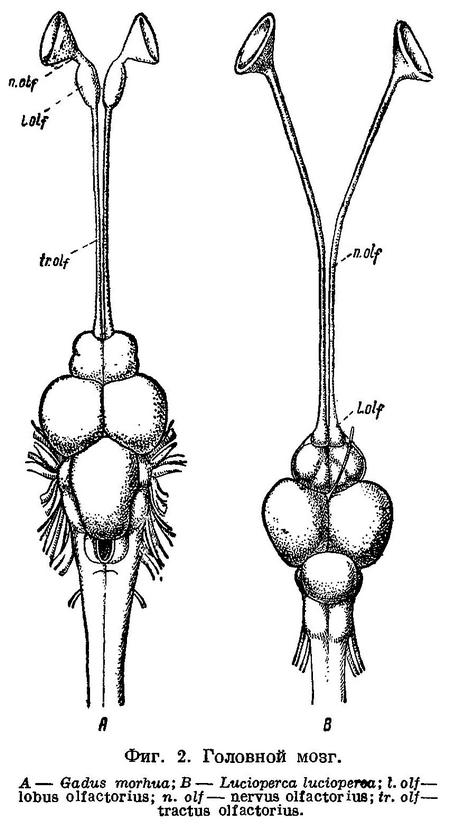
Черепная полость у Gadiformes продолжена вперед над septum interorbitale в виде узкого канала, где расположены обонятельные нервы и через посредство которого черепная полость оказывается сближенной с носовыми капсулами. Благодаря этому, обонятельные нервы у Gadiformes не проходят через орбиту и на всем протяжении расположены друг около друга (фиг. 1, А) в отличие от громадного большинства остальных Teleostei, у которых обонятельный нерв каждой стороны проходит через орбиту и отделен межглазничной перегородкой от нерва другой стороны (фиг. 1, С). Исключением среди Teleostei, подобно Gadiformes, являются далеко стоящие от них семейства Galaxiidae (Swinnerton, 1903 : 61), Siluridae, Cyprinidae (Sagemehl, 1891 : 489), некоторые Characinidae (Sagemehl, 1885 : 1) и Plectognathi (Diodon) (Owen, 1846 : 184, fig. 47), у которых межглазничная область устроена сходным образом. Кроме того, lobi olfactorii у Gadiformes далеко отстоят от переднего мозга, располагаясь в канале над septum interorbitale около носовых капсул, вследствие чего n.n. olfactorii у них очень короткие, а с передним мозгом lobi olfactorii соединяются при помощи длинных tractus olfactorii (фиг. 2, А), что среди Teleostei известно лишь у Galaxiidae (Swinnerton, 1903 : 63), некоторых Cypriniformes (Sagemehl, 1885 : 74; 1891 : 489) и Plectognathi (Owen, 1846 : 184).
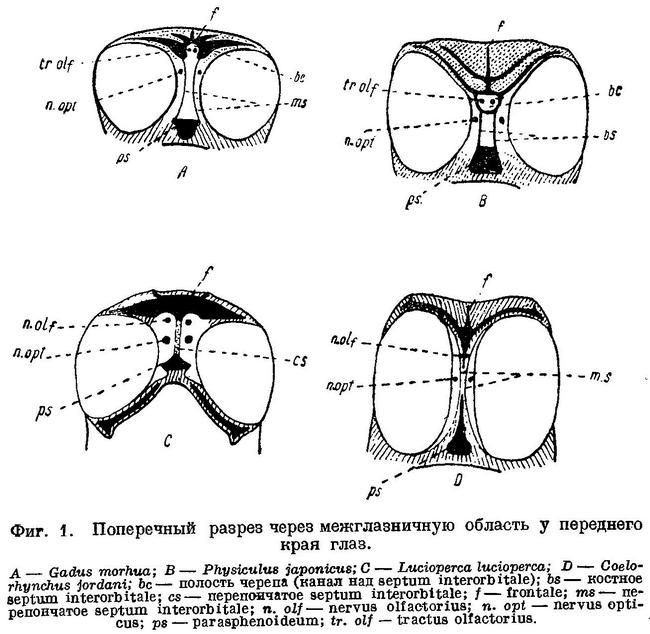
Помимо упомянутых семейств и отрядов рыб, обонятельные нервы, по указанию Dietz'a (1913 : 86), не проходят через орбиту еще у Liparis и Cyclopterus. В межглазничную же область полость черепа продолжена только у Liparis, у которого она здесь такой же ширины, как и сзади, и доходит до самых носовых капсул; что касается Cyclopterus, то у него полость черепа оканчивается спереди на уровне заднего края глаз, далее же обонятельные нервы разделены хрящом, заполняющим всю межглазничную область. Обонятельные нервы через орбиту не проходят у ряда других исследованных нами представителей подотряда Scleroparei (Муохоcephalus, Cottocomephorus, Comephorus, Sebastes, Trigla, Occa), куда относятся обе упомянутые выше рыбы, и Blennioidei (Blennius, Lumpenus, Ophidion), но полость черепа у всех них в межглазничную область не продолжена, за исключением Myoxocephalus, у которого она в виде канала тянется до самых носовых капсул. Однако, что является самым существенным, lobi olfactorii у всех перечисленных рыб расположены непосредственно около переднего мозга и с носовыми капсулами соединены при помощи длинных n.n. olfactorii, которые расположены между перепонкой septum interorbitale и костями или хрящом межглазничной области (см. фиг. 1, D) и через орбиту, таким образом, не проходят. (Указание Dietz'a (1921 : 436) на положение lobi olfactorii у Cyprinidae (фиг. 2) около переднего мозга и у Liparis (фиг. 5) около носовых капсул неверно: как это общеизвестно, у Cyprinidae lobi olfactorii находятся около носовых капсул, у Liparis, по нашим исследованиям, — около переднего мозга. Что касается скелета хвостового плавника, то у Gadiformes он совершенно иного устройства, чем у Cyclopterus, Liparis и Cottus, сближаемых Dietz'eм по этому признаку с Gadiformes, хотя у этих рыб он несколько менее асимметричен, чем у других Scleroparei. Наиболее существенный признак Gadiformes — большое opisthoticum с отверстием для n. glossopharyngeus Dietz не рассматривает.) Канал над septum interorbitale с расположенными в нем обонятельными нервами имеется, как обнаружено нами, и у сем. Brotulidae (Hoplobrotula armata), у представителей которого он, как и у сем. Cyprinidae, костный, но lobi olfactorii тесно примыкают к переднему мозгу, а к носовым капсулам тянутся по каналу длинные n.n. olfactorii.
Таким образом, межглазничная область иногда даже у одной и той же группы рыб устроена довольно разнообразно и в некоторых группах существенного значения как систематический признак иметь не может. Что касается положения lobi olfactorii, то около носовых капсул они расположены, насколько это известно в настоящее время (см. Lissner, 1918 : 127), только у семейств Galaxiidae, Cyprinidae, Siluridae, у некоторых Characinidae, у отряда Plectagnathi и у Gadiformes. На некотором расстоянии от носовых капсул, приблизительно посередине между ними и передним мозгом, расположены lobi olfactorii из Cyprinidae лишь у Carassius и из Gadidae — у Raniceps, вследствие того, что в передней части канала около носовых капсул у этих рыб разрастается хрящ и тем самым lobi olfactorii как бы отодвигаются от носовых капсул. Весьма характерно, что у Brotulidae lobi olfactorii расположены около переднего мозга, несмотря на наличие у них в межглазничной области канала, доходящего до самых носовых капсул. Возможно, что и у тех Characinidae, у которых lobi olfactorii расположены не около носовых капсул, это обусловлено разрастанием здесь хряща.
Lobi olfactorii расположены около носовых капсул, однако, не у всех Gadiformes. У Macrurus cavernosus (Pfüller, 1914 : 48, pi. 1—2), Coelorhynchus jordani, Coelorhynchus coelorhynchus, Coelorhynchus japonicus, Trachirinchus trachirinchus, Coryphaenoides rupestris, Coryphaenoides altipinnis и Abyssicola macrochir (наши исследования) из семейства Macruridae lobi olfactorii тесно примыкают к переднему мозгу, полость черепа в виде канала над septum interorbitale далеко не продолжена, вследствие чего tractus olfactorii отсутствует, а n.n. olfactorii длинные. Кроме того, у всех них оба обонятельных нерва, расположенные вначале друг около друга между перепонками septum interorbitale правой и левой сторон черепа, затем разделяются при помощи frontale и mesethmoideum и проходят к носовым капсулам, располагаясь между этими костями и перепонкой septum interorbitale и минуя, таким образом, орбиту (фиг. 1, В). У семейства Gadidae вдоль черепа с нижней стороны frontalia тянутся два костных гребня, ограничивающие с боков канал над septum interorbitale. У семейства Macruridae посередине черепа с нижней стороны frontalia тянется один непарный гребень, вдоль которого с боков расположены n.n. olfactorii. Нужно полагать, что подобным же образом расположены lobi olfactorii и устроена межглазничная область и у остальных представителей этого семейства. Следовательно, по одним из наиболее важных признаков — положению lobi olfactorii и строению межглазничной области — семейство Macruridae существенно отличается от других семейств Gadiformes. Из признаков, свойственных семейству Macruridae и другим семействам Gadiformes, следует отметить увеличенное opisthoticum с отверстием для n. glossopharyngeus (В этом отношении из Macruridae нами исследован только один вид — Coelorhynchus anatirostris Jord. et Gilb.); остальные признаки свойственны многим другим группам Teleostei.
Таким образом, по некоторым признакам семейство Macruridae должно быть отнесено к Gadiformes, по другим признакам оно должно быть из этого отряда выделено. И то и другое будет справедливо почти в одинаковой степени. Однако, после исключения из Gadiformes семейства Macruridae многие признаки (положение лопаточного отверстия между scapula и coracoideum, только мягкие лучи в непарных плавниках, положение брюшных плавников впереди грудных), не считаемые Regan'ом характерными, становятся общими для всех семейств отряда, и Gadiformes (Anacanthini) можно считать действительно мягкоперыми, объединенными в отряд большим количеством признаков. Поэтому обширное семейство Macruridae, повидимому распадающееся в свою очередь на несколько семейств, следует выделить в самостоятельный отряд, близкий, с одной стороны, к Gadiformes, с другой — к некоторым Acanthopterygii.
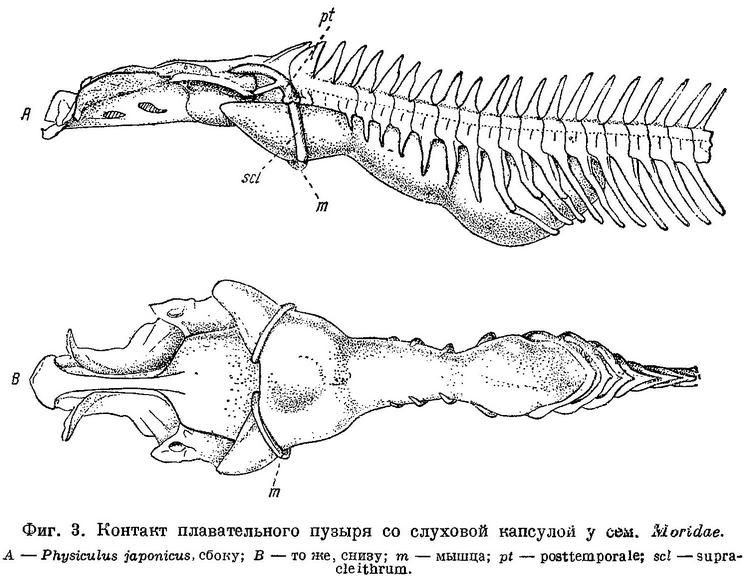
По исследованиям Stannius'a (1844 : 63) и по Goodrich'у (1909 : 478), septum interorbitale и нижняя часть канала n.n. olfactorii у Gadiformes перепончатые (фиг. 1, А). По нашим исследованиям, у значительной части представителей семейства Gadidae канал n.n. olfactorii целиком, a septum interorbitale на значительном протяжении костные (фиг. 1, В). Кроме того, у рыб, с костной septum interorbitale и каналом n.n. olfactorii, neurocranium спереди открывается лишь узким отверстием, будучи почти замкнут расширением и соединением parasphenoideum и frontalia, которые образуют и septum interorbitale. В neurocranium у рыб с костной septum interorbitale сзади с каждой стороны от foramen magnum имеется по большой, затянутой перепонкой фонтанелли, через которые при помощи двух длинных рожкообразных выростов плавательный пузырь находится в контакте с ушными капсулами (фиг. 3, А и В). Соединение плавательного пузыря с ушной капсулой у одного из представителей семейства Gadidae — Lotella bacchus было обнаружено давно (Parker, 1883 : 234), но оставалось мало известным. О том, что подобное соединение плавательного пузыря с ушной капсулой имеется и у некоторых других тресковых рыб, не было известно, и поэтому значения для системы эта важная анатомическая особенность не получила. Целиком костный канал n.n. olfactorii, костное на значительном протяжении septum interorbilale, фонтанелли сзали neurocranium и соединение через них плавательного пузыря с ушными капсулами, — все эти признаки имеют весьма существенное значение, и роды, которые обладают ими, должны быть выделены в особое семейство Moridae.
К семейству Moridae относятся роды: Mora, Lepidion, Uraleptus, Lotella, Physiculus, Laemonema, Antimora и ряд других.
Таким образом, отряд Gadiformes (Anacanthini) состоит из семейств Gadidae, Moridae, Bregmacerotidae. Последнее близко к семейству Gadidae, но по ряду признаков должно быть выделено в самостоятельное семейство. Что касается рода Мuraenolepis, который Regan (1903 : 466) выделяет в особое семейство Muraenolepidae и причисляет к отряду Gadiformes (Anacanthini), то по строению переднего мозга и черепа он не отличается от Gadiformes, но благодаря увеличенному числу грудных radialia (не менее 10) и некоторым другим особенностям (жаберные отверстия узкие, простирающиеся вверх не выше уровня основания грудных плавников, чешуи удлиненные и расположены под углом друг к другу; детальнее см. Световидов, 1939 : 583) он занимает несколько обособленное положение. Увеличенное число грудных radialia имеет важное значение и дает основание для выделения сем. Muraenolepidae в особый подотряд отряда Gadiformes. Таким образом, отряд Gadiformes разделяется на два подотряда: 1) Muraenolepidoidei с одним семейством Muraenolepidae и 2) Gadoidei с семействами Moridae, Bregmacerotidae и Gadidae. Последнее наиболее естественно, по строению взрослых, личинок и икринок, разделяется на подсемейства Lotinae, Мerlucciinae и Gadinae.
Происхождение и родство Gadiformes со всеми сближаемыми, с ними семействами и группами рыб весьма отдаленно. На отдаленность Gadiformes от Pleuronectiformes (Heterosomata) наиболее решительно и достаточно убедительно указывают Соре (1872 : 341) и Cunningham (1897 : 498), выводы которых Boulenger (1902 : 296) и Regan (1910а : 484) подкрепили новыми анатомическими данными. Regan (1903 : 460), указавший ряд новых признаков, хорошо отделяющих Gadiformes от сближаемых с ними семейств Blenniidae, Zoarcidae, Brotulidae, Ophidiidae и других, и Boulenger (1904 : 646, 702, 703) считают, что сходство между этими двумя группами рыб должно скорее быть приписано конвергенции, чем близкому родству. Позднее (1910 : 11) Regan высказывает предположение о происхождении Gadiformes от примитивных Iniomi, родственных ныне живущим Aulopidae. На отдаленность от Gadiformes семейства Aulopidae, имеющих соприкасающиеся parietalia, не разделенные посредством supraoccipitalia, и иное устройство жаберной крышки, указал Gregory (1933 : 207, 380, fig. 85). Кроме того, следует добавить, что канала над septum interorbitale у Aulopidae нет и n.n. olfactorii проходят через орбиту. Наиболее же существенным отличием семейства Aulopidae, равно как и других сближаемых с Gadiformes семейств и групп рыб, является положение lobi olfactorii около носовых капсул. Эта важная анатомическая особенность Gadiformes, характерная лишь для далеко стоящих от них Cyprinidae и Galaxiidae, указывает на раннее отщепление их от ныне живущих групп рыб общего с ними происхождения. При этом следует отметить, что некоторые характерные морфологические особенности, в частности дифицеркальный хвост (Whitehouse (1935 : 70) считает его гомоцеркальным и у взрослых. Barrington (1935 : 270) предлагает для хвостов такого типа название псевдокавдадьный.) и положение lobi olfactorii около носовых капсул, а не около переднего мозга, представляют собой явления вторичные: как известно, у молодых тресковых хвост гомоцеркальный, a lobi olfactorii, по указанию Т. С. Расса (Световидов, 1937 : 1288), у них располагаются чрезвычайно близко от переднего мозга. Л. С. Берг (1940 : 284), помещая Gadiformes в системе почти сразу за Cypriniformes, ниже Perciformes, считает их как низкоорганизованный отряд, происшедший от форм, родственных Pachycormidae. Для Gadidae он дает несколько иное, чем принятое здесь, разделение на подсемейства, основанное на строении только взрослых; в особые подсемейства им выделяются Eleginus (парапофизы расширены и содержат выросты плавательного пузыря) и Raniceps (lobi olfactorii от носовых капсул несколько удалены; о вторичном характере этого у Raniceps и Carassius см. выше).
Система семейства Gadidae разработана очень мало. Даже в работах, посвященных специально семейству Gadidae или отряду Gadiformes, вопросы системы семейства не затронуты или им уделяется мало внимания. Вследствие этого в определителях и в сводках по рыбам тех или иных морей роды тресковых располагаются в самом разнообразном порядке и последовательности.
Наибольшего внимания из всех попыток классифицировать тресковых заслуживает система, принятая Jordan'oм и Evermann'oм (1898 : 2531), которые разбивают все роды семейства на 5 групп: Gadinae, Lotinae, Phycinae, Gaidropsarinae и Brosminae, положив в основу этой группировки весьма существенный признак — строение спинных плавников. Внутри каждой из этих групп роды расположены бессистемно и, кроме того, всю систему несколько путает то, что в нее включены роды, выделяемые нами в семейство Moridae, а род Merluccius принят за особое семейство.
Попытки классифицировать Gadinae, наиболее богатую видами и родами группу семейства Gadidae, сводятся к разделению в свою очередь и их на ряд групп. Williamson (1909 : 111) предлагает ряд группировок Gadinae по многим признакам, причем для каждого из признаков получено различное число групп и распределение по ним родов, вследствие того, что взаимосвязи со средой и функциональное значение этих признаков самые разнообразные (положение рта, начало анального плавника, цвет перитонеум, размеры глаз, размеры усика, длины парных плавников, размеры и даже окраска рыб, — всего 20 признаков).
Классификация, по мнению автора, должна основываться на комплексе признаков, функционально связанных с наиболее характерной биологической особенностью данной группы.
Одной из морфологических особенностей тресковых рыб, свойственной в такой сильно выраженной и изменчивой форме, пожалуй, только им одним, является строение их непарных плавников. Среди них имеются виды, имеющие один, два и три спинных и один и два анальных плавника. Эта особенность тресковых частично нашла свое отражение в системе Jordan'a и Evermann'a. Характерной биологической особенностью тресковых, не менее изменчивой, чем рассматриваемая морфологическая, следует считать их способность к обитанию в различных частях водной среды и неодинаковую приспособленность к движению. Преобладающее большинство тресковых держится у дна, но среди представителей семейства есть и такие, которые живут в толще воды, ведя пелагический образ жизни (Micromesistius). Среди представителей семейства, наряду с видами, держащимися на значительной глубине (Brosme), имеются виды, живущие и на небольшой глубине у берегов. Держась у берегов, одни представители ведут крайне неподвижный образ жизни (Ratiiceps), другие, как, например, Eleginus, значительно более подвижны. Наряду с видами, живущими локально, среди тресковых есть и такие, которые совершают отдаленные миграции. Несомненно, что указанные морфологические и биологические особенности тресковых находятся в теснейшей функциональной связи.
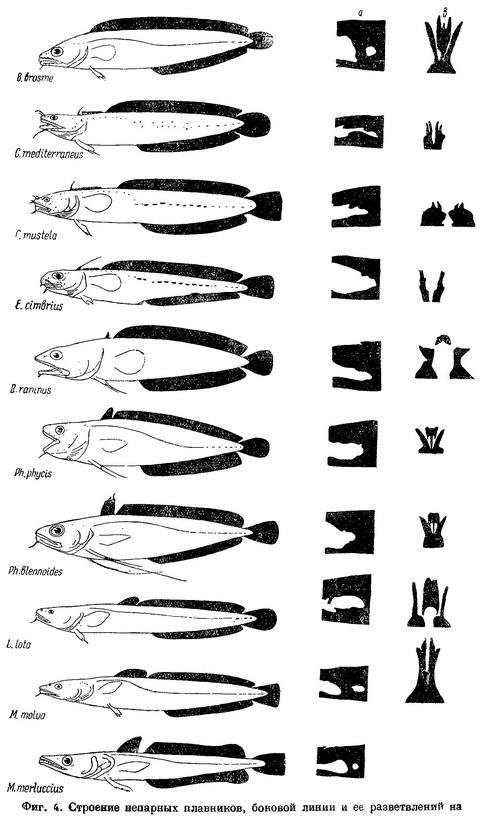
Наиболее совершенной формой строения непарных плавников, как показали экспериментальные исследования de Houssay (1910 : 13), являются не длинные сплошные анальный и спинной плавники, а разделенные на несколько, так как такое строение их наиболее приспособлено для обтекания теми токами, которые при движении рыбы возникают вокруг ее тела. По строению непарных плавников всех тресковых можно расположить в ряд (фиг. 4). Из этого рисунка видно, как, начиная от Brosme и кончая Micromesistius, постепенно дифференцируются спинной и анальный плавники. Brosme имеет один длинный спинной и один анальный плавник, тесно соприкасающиеся, отчасти слитые с хвостовым. У Gaidropsarus, Ciliata и Enchelyopus имеются уже два спинных плавника: первый состоит всего лишь из одного нормально устроенного луча. Остальные лучи первого спинного плавника Gaidropsarus mediterraneus по строению (Bogoljubsky, 1908 : 327) не являются настоящими плавниковыми лучами. Raniceps имеет первый спинной плавник несколько больших размеров (3 луча), a Phycis, Urophycis и Molva — нормальных для тресковых размеров. Merluccius, имеющий в задней части анального и второго спинного плавников по выемке, является как бы переходной формой к тресковым с тремя спинными и двумя анальными плавниками. Далее, у рыб, имеющих три спинных и два анальных плавника, плавники эти, первоначально соприкасающиеся, разделяются все более и более широкими промежутками, причем у Trisopterus luscus анальные плавники часто соединены небольшой перемычкой. К концу ряда анальные плавники снова несколько сближены. Таким образом, мы имеем ряд рыб с постепенно дифференцирующимися непарными плавниками.
В пределах подсемейства Lotinae рыбы в этом ряду располагаются соответственно расчленению первоначально одного спинного плавника на два и началу образования третьего спинного и второго анального плавников. Gadinae дают продолжение этого ряда соответственно дальнейшему разделению спинных плавников увеличивающимися промежутками между ними. При этом совершенно очевидно, что у тех рыб, у которых имеется более чем один спинной и анальный плавник, точно так же как и у рыб с большими промежутками между этими плавниками, общая длина оснований всех спинных и обоих анальных плавников меньше, чем у рыб с нерасчлененными плавниками. Основным руководящим признаком в этом ряду является, как показали подсчеты, все же расчленение плавников и величины промежутков между ними, а не суммы длины оснований их и тем более не количество лучей в них. Так, например, сумма оснований спинных плавников у Trisopterus esmarkii приблизительно равна сумме оснований их у остальных двух видов этого рода, между тем как число лучей в этих плавниках у обоих других видов значительно больше. В еще большей степени относится сказанное к количеству лучей и к сумме длин оснований анальных плавников.
В пределах ряда тресковых, расположенных по степени дифференциации их непарных плавников, наблюдается несколько закономерностей и по другим признакам, причем каждый из этих плавников на протяжении ряда делится на два параллельных ряда, соответственно двум подсемействам Gadidae — Lotinae и Gadinae; подсемейство Merlucciinae занимает промежуточное положение.
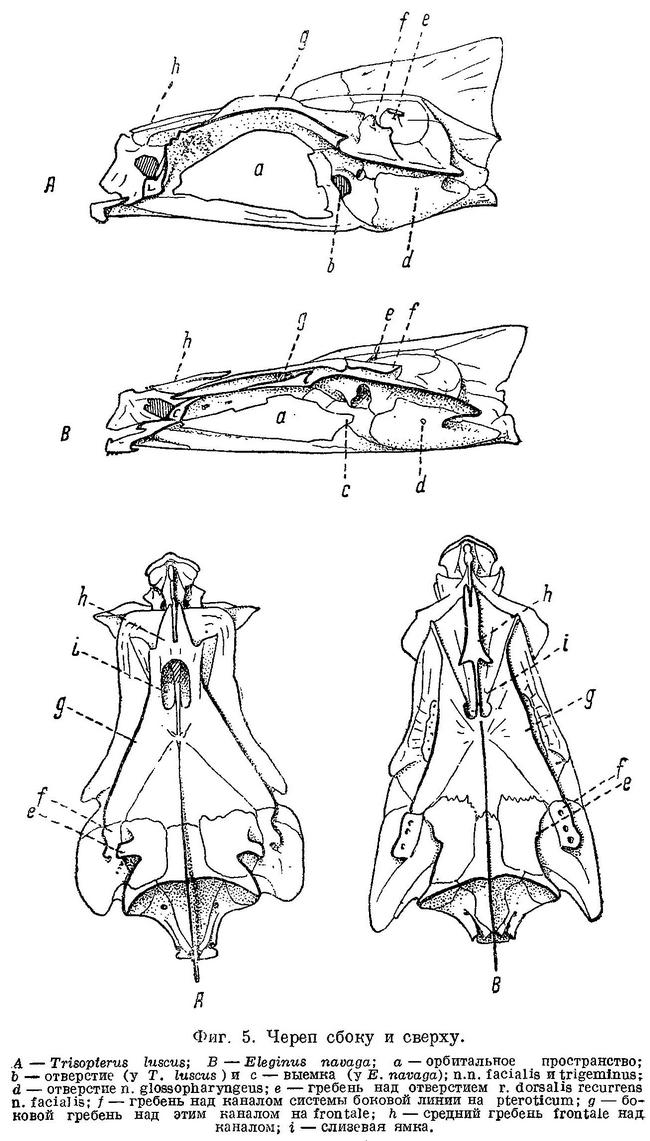
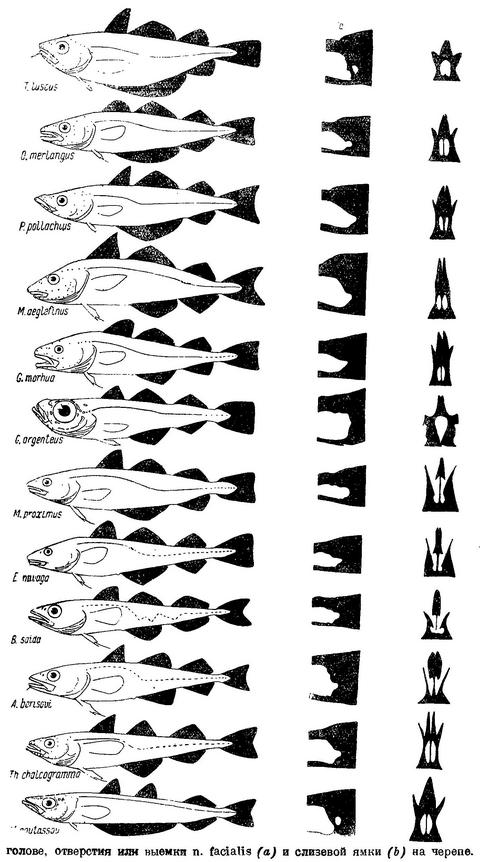
N. facialis выходит из черепа у тресковых вместе с n. trigeminus через отверстие или различной глубины выемку в передней части prooticum сбоку черепа (см. фиг. 4,а и фиг. 5,b,с). У Brosme brosme нерв этот выходит через отверстие (фиг. 4,а), а у следующего за ним по ряду Gaidropsarus mediterraneus — через глубокую выемку в prooticum, края которой настолько сближены, что почти замыкают ее спереди. У Ciliata mustela и далее следующих за ним в ряду рыб выемка становится спереди более широкой и менее глубокой. У Lota lota выемка так же глубока и края спереди сужены почти так же, как у Gaidropsarus mediterraneus, а у всех видов Molva и Merluccius n. facialis из черепа выходит опять через отверстие. Через отверстие n. facialis выходит и у следующих далее в ряду видов рода Trisopterus. При этом ширина костной перемычки у Trisopterus luscus несколько шире, чем у Trisopterus minutus, а у последней шире, чем у Trisopterus esmarkii. Odontogadus merlangus и далее следующие за ним представители семейства имеют в prooticum выемку, которая последовательно становится спереди более широкой и менее глубокой. У обеих форм Gadiculus argenteus выемки почти нет. Далее, у видов родов Eleginus, Microgadus, Boreogadus, Arctogadus и Theragra выемка постепенно углубляется, а края ее становятся более сближенными. Micromesistius имеет сбоку черепа отверстие. Таким образом, по этому признаку ряд тресковых, расположенных по степени дифференциации их непарных плавников, разделяется в свою очередь на два параллельных ряда, соответственно делению семейства на подсемейства. Такой же закономерности подчиняется изменение и некоторых других признаков.
Сверху черепа тресковых на каждом parietale расположено по отверстию, через которое выходит r. dorsalis recurrens n. facialis (фиг. 5,е и рис. В на табл. XXI—LXVII). У одних тресковых каждое из этих отверстий прикрыто у каждого вида в различной степени развитыми гребнями; у других гребень не развит, и отверстие сверху открыто. У Brosme brosme и далее следующих за ним в ряду Gaidropsarus mediterraneus, Ciliata mustela, Enchelyopus cimbrius гребни, прикрывающие эти отверстия, имеются. У Raniceps ranius и всех видов родов Phycis и Urophycis отверстия r. dorsalis recurrens n. facialis не прикрыты, так как гребни у них имеют вид едва заметных выростов, расположенных несколько сзади отверстий. У Lota, всех видов Molva и Merluccius гребни развиты и прикрывают отверстия. В пределах подсемейства Gadinae гребни на parietalia развиты сильнее. У всех видов родов Trisopterus, Odontogadus, Pollachius, Gadus и Melanogrammus гребни развиты, размеры их уменьшаются от Trisopterus к Melanogrammus. Нет гребней у Gadiculus. Далее гребень есть, постепенно увеличиваясь к концу ряда, у Eleginus, Microgadus, Boreogadus, Arctogadus, Theragra и Micromesistius, причем у одних особей Eleginus navaga гребни есть, у других они не развиты.
N. glossopharyngeus (Stensiö, 1921 : 155) выходит из черепа через небольшое отверстие на opisthoticum (фиг. 5,d и рис. А на табл.XXI—LXVII). Величина этого отверстия, изменяясь в пределах ряда, подчиняется той же закономерности. У Brosme brosme отверстие почти не заметно. У Gaidropsarus mediterraneus, Ciliata mustela, Enchelyopus cimbrius, Raniceps raninus оно небольшое, у видов родов Phycis и Uropkycis несколько больше, у Lota и Molva большое и у Merluccius вновь небольших размеров. Далее, у всех видов рода Trisopterus отверстие едва заметно, размеры его постепенно увеличиваются до Gadus и Melanogrammus и, начиная с Gadiculus, уменьшаются вновь.
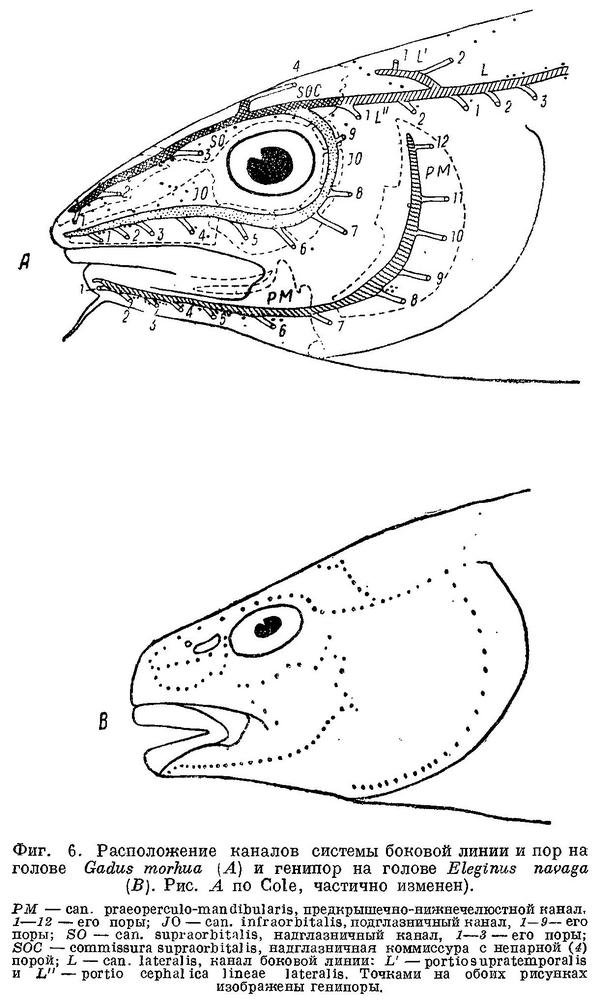
Боковая линия у тресковых образует на голове с каждой стороны каналы (фиг. 6): предкрышечно-нижнечелюстной, подглазничный и надглазничный с короткой комиссурой, соединяющей правый и левый каналы. Сама боковая линия имеет на голове надвисочную часть (L '), над которой образуется окостенение из 4—5 косточек ossa tabularia (= supratemporale). Каждый канал системы боковой линии на голове прикрыт сверху почти на всем своем протяжении гребнем тех костей, на которых эти каналы расположены (frontale, pteroticum, sub- и praeorbitale, nasale, praeoperculum, tabulare). Среди этих гребней наиболее существенное значение имеет средний гребень frontale (фиг. 5, h) и боковые гребни frontale и pteroticum (фиг. 5, f и g). Межглазничная комиссура надглазничного канала расположена в особом углублении frontale — слизевой ямке (фиг. 5, i). Слизевая ямка у одних представителей Gadidae спереди замкнута сросшимися краями боковых передних гребней над каналами системы боковой линии на frontale, у других широко открыта, так как края боковых и средних гребней frontalia укорочены и не срастаются впереди слизевой ямки. У Brosme brosme края боковых и средних гребней frontalia настолько сближены, что почти замыкают спереди небольших размеров слизевую ямку. У Gaidropsarus mediterraneus, Ciliata mustela и Enchelyopis cimbrius слизевая ямка спереди открыта, очень небольших размеров. Raniceps raninus имеет слизевую ямку спереди открытой. У одних видов родов Phycis и Urophycis слизевая ямка спереди открыта, у других закрыта; оба эти рода представляют в этом отношении небольшое отклонение от общего правила, о причинах чего будет сказано ниже. У Lota lota слизевая ямка спереди почти замкнута сближенными краями боковых и средних гребней frontalia, которые у видов рода Molva, срастаясь, образуют хорошо замкнутую ямку. Слизевая ямка у всех видов рода Merluccius представляет собой обширное углубление посередине frontalia и имеет несколько уклоняющийся от обычного для тресковых вид. Слизевая ямка замкнута далее, начиная с Trisopterus и кончая Gadiculus. У всех далее следующих в ряду видов слизевая ямка спереди открыта и замкнута вновь у стоящих в конце его Theragra chalcogramma и Micromesistius poutassou и Micromesistius australis. При этом следует отметить, что закрытая или открытая спереди слизевая ямка обычно свойственна всем видам любого рода тресковых, за исключением, как указывалось выше, родов Phycis и Urophycis. Кроме того, есть один вид — Eleginus navaga, — у которого сближенные края боковых и средних гребней frontalia иногда бывают сращены с одной стороны, а на одном из исследованных черепов и с обеих сторон. Таким образом, эта особенность находится в неустойчивом состоянии в первом случае в пределах рода, во вторых — в пределах вида, стоящих в середине каждого из параллельных рядов.
Подобная же закономерность наблюдается и в строении гребней над самыми каналами боковой линии на frontalia. У Brosme, Gaidropsarus, Ciliata, Enchelyopus, Molva, Trisopterus, Micromesistius, т. е. у рыб, стоящих в начале и конце каждого ряда, края гребней на frontalia тесно примыкают к этим костям, почти замыкая канал. Более того — у некоторых из этих рыб (Trisopterus) края гребней местами соединяются с frontalia при помощи костных перемычек. У всех остальных расположенных в ряду между этими рыбами тресковых края гребней над слизевыми каналами на frontalia обычно приподняты. Несколько уклоняется в этом признаке род Merluccius, у которого гребни развиты очень слабо.
Вдоль каждого из каналов системы боковой линии на голове расположены многочисленные группы нервных окончаний (фиг. 6, В) — генипор (pit-organs), или каналы эти открываются наружу рядом пор (фиг. 6, А), причем единичные генипоры имеются и в этом случае. Боковая линия на теле у тресковых тянется от головы до хвостового плавника в виде непрерывной или прерывистой, разделенной небольшими промежутками, линии (фиг. 4). У Brosme brosme боковая линия тянется непрерывной полосой от головы до конца спинного плавника, становясь прерывистой лишь на хвостовом стебле (фиг. 4). У Gaidropsarus mediterraneus, Ciliata mustela и Enchelyopus cimbrius боковая линия прерывиста на всем протяжении, у Raniceps raninus ее, повидимому, нет совершенно. У видов родов Phycis и Urophycis и у Lota lota боковая линия непрерывна почти до конца анального плавника, далее прерывиста; у видов рода Moiva она прерывиста сзади от конца анального плавника, у видов Merluccius — непрерывна на всем протяжении. У видов родов Trisopterus, Odontogadus, Pollachius и Melanogrammus боковая линия непрерывна на всем протяжении; у видов родов Gadus, Gadiculus (Экземпляры Gadiculus обычно бывают без чешуй, и поэтому о строении их боковой линии судить трудно, однако до середины третьего спинного плавника она тянется без перерывов.) и Microgadus она прерывиста сзади от середины или конца третьего спинного плавника, у видов рода Eleginus прерывиста уже сзади от начала второго спинного плавника и у видов родов Boreogadus и Arctogadus боковая линия прерывиста на всем протяжении. Далее, у Theragra chalcogramma боковая линия прерывиста лишь сзади от начала второго спинного плавника и у Micromesistius poutassou и Micromesistius australis без перерывов на всем протяжении.
Подобной же закономерности подчиняется в ряду тресковых и наличие у них на голове пор или генипор (см. точки на голове, фиг. 4). Роды, стоящие в начале и конце подсемейств Lotinae и Gadinae, образующих два параллельных ряда в общем ряду тресковых, имеют на голове поры: у родов, расположенных посередине каждого из тих параллельных рядов (Raniceps и Lota в первом, Microgadus, Eleginus, Boreogadus и Arctogadus во втором), имеются только генипоры. Исключением в первом ряду являются лишь роды Phycis и Urophycis, стоящие в середине ряда Lotinae, но имеющие поры. Оба эти рода представляют некоторое исключение, как указывалось выше, и по строению слизевой ямки. Следует отметить, что все виды, за исключением некоторых из родов Phycis и Urophycis, имеющие слизевую ямку спереди открытой, имеют на голове лишь генипоры. Все эти виды, опять-таки за исключением родов Phycis и Urophycis, живут в прибрежной области, многие заходят в пресную воду или даже живут здесь постоянно. Несколько уклоняющийся строением слизевой ямки от обычного для тресковых типа род Merluccius отличается от стоящих около него в системе родов и отсутствием слизевых пор.
В ряду тресковых, расположенных по степени дифференциации их непарных плавников, промежутки между анальными плавниками у Gadinae в пределах ряда, как указывалось выше, первоначально увеличиваясь, к концу ряда вновь сближены (фиг. 4). Параллельно с этим перемещается и положение начала анального плавника: у Trisopterus оно расположено под первым спинным плавником, у стоящих далее в ряду постепенно отодвигается назад, начиная с Eleginus, вновь перемещается вперед, и у обоих видов Micromesistius анальный плавник начинается впереди первого спинного.
Таким образом, ряд тресковых рыб, расположенный по степени дифференциации непарных плавников, по всем остальным признакам распадается соответственно делению семейства на Lotinae и Gadinae на два параллельных ряда; подсем. Merlucciinae занимает промежуточное положение. В каждом из рядов все признаки изменяются в общем параллельно один другому.
Изменчивость каждого из этих признаков, в особенности отверстия или выемки n. facialis сбоку черепа, ограничена довольно узкими пределами. У трески, например, выемка n. facialis на всех исследованных черепах и рисунках их в других работах вариирует мало. Наиболее вариирующим оказывается отверстие n. glossopharyngeus на opisthoticum: на одном из черепов у Pollachius virens отверстие справа менее отверстия слева. Точно так же и на рисунке черепа Pollachius pollachius (Williamson, 4902, табл. VII, фиг. 41) отверстие это небольшое, а на фотографии (l. с., табл. IX, фиг. 28) — нормальной для этого вида величины. То же самое наблюдается и у других видов, изученных по нескольким или многим черепам, но в общем закономерное изменение как этой, так и других рассматриваемых особенностей совершенно очевидно даже и при большей степени их изменчивости.
Из фактов онтогенеза известно, что при развитии личинки и малька любой тресковой рыбы происходит постепенная дифференциация эмбриональной плавниковой складки (См. таблицы в работах J. Schmidt'a (1905, 1905а, 1906, 1906а, 1907, 1907а).). В тянущейся вдоль всего тела эмбриональной плавниковой складке личинки, соответственно положению хвостового, спинных и анальных плавников взрослой рыбы, закладываются лучи, причем у Brosme (J. Schmidt, 1905а, табл. I, фиг. 1—12) лучи эти закладываются равномерно вдоль всей спины сверху и задней половины тела снизу, а у рыб с более чем одним спинным и анальным плавниками на месте промежутков между будущими плавниками лучи укорочены (если у взрослых плавники соприкасаются) или отсутствуют совершенно (J. Schmidt, 1905, табл. I—III). Крайние лучи двух соседних плавников в эмбриональной плавниковой складке у мальков всех тресковых, даже у Micromesistius poutassou, у которой во взрослом состоянии плавники разделены широкими промежутками, соприкасаются или разделены очень узкими промежутками. Одновременно с закладкой лучей, в эмбриональной плавниковой складке против перерывов между плавниковыми лучами, на месте будущих промежутков между плавниками взрослой рыбы образуются выемки, которые, углубляясь, постепенно разделяют эмбриональную плавниковую складку на отдельные плавники. Спинные и анальные плавники соприкасаются у вполне сформировавшегося малька и сеголетка, но по мере роста его у тех видов, у которых во взрослом состоянии плавники разделены промежутками, плавниковые перепонки у заднего края плавников постепенно редуцируются, и, таким образом, образуются промежутки, увеличивающиеся по мере роста рыбы.
Строение каналов боковой линии на голове имеет важное филогенетическое значение. У Holostei и низших Teleostei (Cypriniformes, Clupeiformes) эти каналы расположены в костях и сообщаются с наружной средой посредством узких отверстий, пронизывающих местами канал (Goodrich, 1930 : 294—295). У более высокоорганизованных Teleostei кожные кости погружаются глубже под кожные покровы, каналы менее замкнуты в костях и, как у Gadiformes, представляют бороздку, прикрытую гребнями и лишь частично замкнутую в костный канал. Слизевая ямка у мальков менее 60 мм длины, повидимому, спереди не замкнута, так как у мальков длиной 60—65 мм средние и боковые гребни frontalia соединены при помощи перепонки, которая у рыб размерами 85 мм и более окостеневает. Как происходит развитие остальных признаков — неизвестно.
Таким образом, ряд тресковых, расположенный по этим признакам, до некоторой степени является отражением и филогенетических отношений представителей семейства. Однако нельзя считать, как это делают Holt и Calderwood (1895 : 440), что от стоящего в начале ряда и более примитивно устроенного Brosme последовательно произошли все остальные члены ряда. Наиболее близко к исходной форме всех тресковых стоят, повидимому, роды Phycis и Urophycis, сохранившие и во взрослом состоянии некоторые черты строения личинки: брюшные плавники у всех видов этих родов, подобно личинкам Lotinae, длинные и состоят всего из трех лучей. Это предположение находит себе подтверждение, нам кажется, и в том, что одна из важных из числа рассматриваемых особенностей черепа тресковых — слизевая ямка — у видов родов Phycis и Urophycis, как сказано выше, имеет строение, свойственное всем видам семейства. Вместе с тем Phycis и Urophycis имеют черты и более высокой организации, свойственной всем Gadinae: frontalia у всех исследованных в этом отношении видов обоих родов срастаются в одну непарную пластинку, связывая, таким образом, подсемейства тресковых.
Предположение, что роды Phycis и Urophycis наиболее близко стоят к исходной форме семейства, находит себе еще подтверждение и в том, что среди тресковых не известно такой рыбы с одним спинным плавником, у которой была бы в нем выемка, подобная наблюдаемой у Merluccius в задней части второго спинного и анального плавников. Наоборот, у Raniceps и Gaidropsarus происходит постепенное редуцирование задней части первого спинного плавника, отделяемого вследствие этого от второго спинного плавника промежутком. У мальков Raniceps первый спинной плавник более длинный и соприкасается со вторым; по мере роста малька задняя часть этого плавника редуцируется и вследствие этого между ним и вторым спинным плавником образуется промежуток (Schmidt, 1907а, табл. I, фиг. 9—13). У мальков Gaidropsarus первый спинной плавник устроен нормально, но по мере роста задняя часть плавника постепенно редуцируется (D'Ancona, 1932, табл. XIII, фиг. 15—20, табл. XIV, фиг. 13—22), и, кроме того, строение его у взрослой рыбы показывает, что это явление вторичное, произошедшее путем регресса (Bogoljubsky, 1898 : 382). Следовательно, у одних тресковых спинные плавники подвергаются дальнейшей дифференциации — в задней части второго спинного плавника отделяется третий, у других упрощаются — задняя часть первого спинного плавника редуцируется, и этот плавник у рассматриваемых родов почти исчезает.
Как указывалось выше, Gadinae, имеющие более расчлененные плавники, в общем ведут и более подвижный образ жизни, чем Lotinae со слабо дифференцированными плавниками. Соответственно периодическому изменению в ряду остальных признаков, в общем параллельно изменяются и биологические особенности тресковых рыб. Brosme brosme и все виды родов Molva и Merluccius, стоящие в начале и конце ряда Lotinae, живут на значительных и даже на больших глубинах. Все остальные представители этого подсемейства являются значительно более мелководными рыбами. На более значительных глубинах живут и все виды родов Trisopterus, Odontogadus, Pollachius, Melanogrammus, Gadus, Thera- gra, Micromesistius, стоящие в начале и в конце ряда Gadinae, в то время как Microgadus, Eleginus, Boreogadus и Arctogadus живут на значительно меньших глубинах прибрежной области. Исключением среди Gadinae являются обе формы Gadiculus argenteus, живущие на значительных глубинах, и отчасти Pollachius virens, держащийся в поверхностных слоях воды, среди Lotinae — некоторые виды Gaidropsarus, встречающиеся на глубинах.
Классификация икринок и личинок тресковых, основывающаяся на приспособлении их к пелагическому образу жизни, целиком совпадает с разделением семейства на два параллельных ряда. Икринки Lotinae и Merlucciinae имеют жировую каплю (Расс, 1936 : 300), а у личинок их брюшные плавники удлинены и состоят лишь из трех лучей. У всех Gadinae икринки без жировой капли и брюшные плавники у личинок не удлинены. У Merlucciinae, занимающего промежуточное положение между Lotinae и Gadinae, брюшные плавники удлинены не так сильно, как у Lotinae, Личинки подсемейства Gadinae классифицируются, главным образом, по пигментации, причем их разделяют на три группы: 1) Pollachius pollachius, Pollachius virens, Gadus morhua, Eleginus navaga, Eleginus gracilis, Boreogadus saida, 2) Trisopterus luscus, Trisopterus minutus, Trisopterus esmarkii, Melanogrammus aeglefinus, Odontogadus merlangus, Micromesistius poutassou, 3) Gadiculus argenteus argenteus, Gadiculus argenteus thori. С предложенной здесь системой семейства Gadidae система личинок подсемейства Gadinae согласуется мало, так как основывается она совершенно на иных признаках — пигментации. Но все же следует отметить, что в первую группу входят виды, стоящие в середине ряда Gadinae, а во вторую, главным образом, виды, стоящие по краям ряда.
Таким образом, параллелизм в рассматриваемых морфологических особенностях и в значительной степени в экологии тресковых обоих родов довольно полный. Экологическим исключением являются упомянутые выше и отчасти некоторые другие виды. Относительно морфологических особенностей следует отметить, что одни из них более резко выражены у Lotinae (глубина выемки в prooticum), другие у Gadinae (гребни на parietalia), а некоторые признаки (слизевая ямка) у Lotinae имеют несколько уклоняющееся строение.
Как видно из всего вышеизложенного, изменение рассматриваемых признаков происходит не одинаково: один признак — строение непарных плавников — на протяжении ряда изменяется в сторону последовательной дифференциации, изменение всех остальных признаков обнаруживает циклические отклонения в сторону возрастания и убывания их, как если бы они были расположены по восходящей спирали, стержнем которой является возрастающая дифференциация строения непарных плавников. При таком расположении род Brosme ряда Lotinae пришелся бы под стоящим в начале ряда Gadinae родом Trisopterus, род Merluccius пришелся бы под родом Micromesistius. Средние роды Lotinae пришлись бы под аналогичными им родами Gadinae. Следует отметить, что роды, расположенные в середине каждого ряда, обладают некоторыми и другими сходными признаками: у Boreogadus и Arctogadus на верхней части черепа позади слизевой ямки есть поперечные гребни, которые имеются и у Phycis и некоторых видов рода Urophycis.
Какова же взаимная связь всех этих разнообразных и на первый взгляд несущественных по их функциональному значению признаков?

Непарные плавники, как мы видели, играют существенную роль в движении рыб. Под гребнями слизевых каналов и в слизевой ямке на frontalia расположены каналы системы боковой линии, при помощи которых, как известно, рыба ориентируется в струях воды при движении. Через отверстие или выемку в prooticum и отверстие в parietalia сверху черепа выходят ветви n. facialis, который, как известно, иннервирует разветвления органа боковой линии на голове и спинной и грудной плавники (фиг. 7, be, hmf, so, rfd). Через отверстие в opisthoticum у тресковых выходит n. glossopharyngeus (Stensio, 1921 : 125), через якобсонову анастомозу, являющуюся частью r. palatinus этого нерва, находящийся в связи с n. facialis (Cole, 1898 : 148).
В комплексе этих признаков, следовательно, имеется приспособление к движению в различных водных слоях, к ощущению движения и к передаче ощущений в мозг посредством нервов, выходящих из черепа через различные по размерам и форме отверстия и выемки в черепе. Таким образом, все лежащие в основе системы тресковых признаки находятся в тесной функциональной взаимной связи. Возможно, что подобным же образом соответственно изменяется и строение мозга, в устройстве некоторых частей которого известны изменения в связи с образом жизни и питания (Evans, 1936 : 367).
По устройству непарных плавников представителей семейства Moridae можно расположить в такой же ряд, как и представителей семейства Gadidae. В начале этого ряда будет стоять род Brosmiculus, в конце — Tripterophycis. Для построения системы семейства Moridae нет серии скелетов, охватывающих если не большинство видов, то хотя бы все роды семейства, чтобы иметь возможность выбрать признаки, на которых основывать систему. Судя по небольшому количеству черепов, приходящихся лишь на немногие роды семейства Moridae, можно полагать, что взаимосвязь в некоторых признаках, подобная таковой семейства Gadidae, имеется и здесь. Более того, здесь имеются роды Laemonema и Laemonemodes, брюшные плавники у которых, как и у родов Phycis и Urophycis из семейства Gadidae, состоят из трех лучей.
Наиболее примитивным из всех семейств Gadiformes является семейство Muraenolepidae, увеличенное число radialia в грудных плавниках которого, свойственное еще лишь семействам Gymnotidae и Anguillidae, следует рассматривать как признак низкой организации. По строению черепа Muraenolepis более всего напоминает некоторые Gadidae из подсемейства Lotinae, но, вопреки мнению Regan'a, ближе всего он не к Onos (= Enchelyopus), а к Phycis и Urophycis. Кроме того, семейство Muraenolepidae, точно так же как и семейство Moridae, сходно с подсемейством Lotinae и по ряду других особенностей: непарные плавники у них дифференцированы не в такой сильной степени, как у Gadinae, икринки всех изученных в этом отношении Moridae имеют жировую каплю, а личинки — удлиненные брюшные плавники.
Тропическое семейство Bregmacerotidae, отличающееся от семейства Gadidae не в такой сильной степени, как семейство Moridae, наиболее близко систематически к одному из самых теплолюбивых представителей семейства Gadidae — роду Gaidropsarus.
ИСКОПАЕМЫE ТРЕСКОВЫЕ
Многочисленные остатки в виде отолитов, относимые ко многим родам семейства Gadidae, встречаются во всех третичных отложениях почти всех европейеких стран, начиная с верхнего палеоцена (Frost, 1931 : 105; 1934 : 500; Leriche, 1910 : 229; Koken, 1884 : 546; 1885 : 113; 1891 : 83; Chaine et Duvergier, 1928 : 190). По этим отолитам описано большое количество родов и видов тресковых, но, вообще говоря, трудно на основании одних отолитов устанавливать не только виды, но даже и роды ископаемых тресковых, чему прекрасным примером могут служить исследования Weiler'a (1935 : 35), установившего принадлежность почти всех известных из нижнетретичных отложений Merluccius к другому роду — Nemopteryx.
Несомненные остатки в виде отпечатков, по которым с достоверностью можно сказать о принадлежности их к тому или иному вымершему или ныне живущему роду тресковых, известны лишь начиная с нижнего олигоцена. Представители семейства Bregmacerotidae (род Bregmaceros) найдены в эоценовых отложениях. Семейство Moridae известно в ископаемом состоянии только по отолитам из олигоценовых отложений Новой Зеландии.
Сем. GADIDAE
Ниже перечислены представители семейства тресковых, известные по отпечаткам. Виды и роды тресковых, описанные по одним лишь отолитам, не упоминаются.
Род BROSME (CUVIER) OKEN
Brosme susedanus Kner (= Brosmius susedanus Kner, 1863 : 145; Kramberger, 1883 : 68 = Brosmius cfr. susedanus Erasmo, 1930 : 82). Верхний миоцен Хорватии (Podsused).
Brosme fuchsianus Kramberger (= Brosmius fuchsianus Kramberger, 1883 : 68). Верхний миоцен Хорватии (Podsused).
Brosme strossmayeriKvamberger (= Brosmius strossmayeri Kramberger, 1883 : 69). Верхний миоцен (?) Хорватии (Beocin).
Brosme elongatus Kramberger (= Brosmius elongatus Kramberger, 1883 : 69; 1885 : 63; Леднев, 1914 : 3; Смирнов, 1936 : 27; = ? Onobrosmius elongatus ponticus Богачев, 1938 : 38). Верхний миоцен Хорватии (Dolje, Sused), верхний миоцен — нижний плиоцен Апшеронского п-ова (рыбные глины и кремнистые мергели окрестностей Бинагадов, понтические глины Кировского района) и Северного Кавказа (глинистые сланцы окрестностей Дзауджикау).
Brosme murdjadjensis Arambourg (= Brosmius murdjadjensis Arambourg, 1937 : 59). Верхний миоцен (ярус Sahelien) Алжира (пров. Оран, Raz-el-Ain).
Brosme oligocaenicus Bogacev (= Brosmius oligocaenicus Богачев, 1933 : 27). Нижнемайкопская свита (олигоцен?) Кавказа (окрестности Сумгаита).
Brosme andrussovi Bogacev (= Brosmius andrussovi Богачев, 1933 : 45). Средний миоцен Апшеронского п-ова (спириалисовые слои).
Brosme sp. Lednev (= Brosmius sp. Леднев, 1914 : 5). Верхний миоцен — нижний плиоцен Апшеронского п-ова (рыбные глины и мергели окрестностей Бинагадов).
Род PHYCIS RÖSE
Phycis suessi Steindachner (= Brosmius Heckel, 1851 : 157 Phycis suessi Steindachner, 1860 : 566). Нижний плиоцен окрестностей Вены (Inzersdorf).
Отолиты, относимые к современному виду Urophycis tenuis, найдены в миоценовых отложениях восточной Германии (Chaine et Duvergier, 1928 : 191).
Род PSEUDOLOTA BOGACEV
Pseudolota abbreviata Bogacev (= Lota sp. Леднев, 1914 : 36 = Pseudolota abbreviata Богачев, 1933 : 28). Нижний миоцен Апшеронского п-ова (верхнемайкопская свита окрестностей Баку) и Шабандага.
У описанного Ледневым Lota sp. Богачев по рисунку устанавливает, что второй спинной и анальный плавники разделены на два. Мною исследован самый отпечаток и деления этих плавников на две части не обнаружено. Второй спинной плавник в указанном Богачевым месте разделения его на два имеет несколько лучей, прижатых к спине и на вершине прикрытых породой, вследствие чего на рисунке и кажется, что плавник разделен; эти лучи ни в коем случае не могут быть очень тонкими и на отпечатках едва заметными последними лучами плавника. Анальный плавник у 6-го луча не может быть разделен на два, потому что 5-й, 6-й и 7-й лучи на отпечатке наиболее длинные. Несколько больший, чем между другими лучами, промежуток между 5-м и 6-м лучами обусловлен тем, что они несколько более раздвинуты; почти такой же промежуток виден далее между 7-м и 8-м лучами.
К описанию этого отпечатка Ледневым следует добавить, что грудные плавники на отпечатке очень длинные, заходят за начало анального плавника; брюшные короче, конец их сохранился плохо. Остистые отростки первых 12—15 позвонков у основания расширены, соприкасаются друг с другом, коротки и не сильно наклонены назад. Строение остистых отростков ближе напоминает род Molva, чем Lota. От современных представителей рода Molva эта рыба отличается значительно меньшим (45) количеством позвонков (у Molva molva их 63—65, Molva dipterygia dipterygia 76—79, Molva dipterygia elongata 82—84).
Род LOTA (CUVIER) OKEN
Lota hulai Pietschmann (1934 : 48). Нижний плиоцен окрестностей Вены. Единственный представитель ископаемых тресковых, найденный в пресноводных или солоноватоводных (Congerien-Tegeln) отложениях.
Род NEMOPTERYX AGASSIZ
Близок к роду Merluccius, от которого отличается узкими, нормальными для тресковых парапофизами и несоприкасающимися друг с другом, суженными при основании остистыми отростками. Грудные плавники сильно удлинены; два спинных и один анальный с выемками в задней части второго спинного и анального плавников. Вымерший род.
Nemopteryx crassus Agassiz (= Nemopteryx crassus Agassiz, 1842 : 75 = Nemopteryx elongatus Agassiz, 1842 : 76 = Palaeogadus troschelii v. Rath, 1859 : 126 = Palaeobrosmius elongatus v. Rath, 1859 : 130 = Nemopteryx troscheli Wettstein, 1886 : 59 = Nemopteryx elongatus Rychlycki, 1909 : 760 = Gobius elongatus Rychlycki, 1909 : 759). Нижний — средний олигоцен Швейцарии (кантон Гларус) и нижние слои среднего олигоцеиа галицийских Карпат.
Nemopteryx emarginatus (Koken) [= Otolitus (Merluccius) emarginatus Koken, 1884 : 547 = Merluccius emarginatus Weiler, 1928 : 50 = Nemopteryx emarginatus Weiler, 1935 : 34]. Средний олигоцен Германии (Майнцский бассейн).
Nemopteryx baschkaensis (Kramberger) (= Megalolepis baschkaensis Kramberger, 1879 : 61 = Merluccius baschkaensis Kramberger, 1882 : 111 = Nemopteryx baschkaensis Weiler, 1935 : 34). Нижние слои среднего олигоцена (Ваschka, Krakowiza).
Nemopteryx latus (Kramberger) (= Megalolepis latus Kramberger, 1879 : 62 = Merluccius latus Kramberger, 1882 : 11 = Nemopteryx latus Weiler, 1935 : 34). Нижние слои среднего олигоцена Карпат (Baschka).
Nemopteryx leptosomus (Kramberger) (= Gobius leptosomus Kramberger, 1879 : 63 = Nemopteryx leptosomus Weiler, 1935 : 34). Повидимому, идентичен с Nemopteryx baschkaensis. Нижний олигоцен Карпат (Baschka).
Nemopteryx athanasiui Pauca (Pauca, 1932 : 149; 1934 : 639; Weiler, 1933 : 20). Нижние слои среднего олигоцена Карпат (Dukla, Jaslo, Krosno, Suslanesti), Моравии (Nikoltschitz) и Венгрии (Eger).
Nemopteryx elongatus (Simonescu) (= Gobius elongatus Simonescu, 1905 : 120 = Nemopteryx elongatus Pauca, 1934 : 641 = Nemopteryx elongatus Weiler, 1935 : 34). Нижние слои среднего олигоцена Карпат (окрестностей Piatra, Молдавия).
Nemopteryx kubacskai Weiler (Weiler, 1935 : 28). Олигоцен Венгрии (Будапешт).
Nemopteryx sp. Weiler (= Merluccius sp. Weiler, 1928 : 77 = Nemopteryx sp. Weiler, 1935 : 34). Нижние слои среднего олигоцена Молдавии (Blatt Mainest, Jud. Bacau).
Nemopteryx sp. Bogacev (Богачев, 1933 : 28). Олигоцен — нижний плиоцен Сев. Кавказа (верхнемайкопская свита близ Перкишкюля).
Помимо нижнего и среднего олигоцена, Nemopteryx, возможно, встречается и в верхнем олигоцене, а может быть и в миоцене, где найдены отолиты, напоминающие отолиты Nemopteryx (Weiler, 1935 : 39).
Род MERLUCCIUS RAFINESQUE
Merluccius macroactus (Kramberger) (= Gobius macroactus Kramberger, 1879 : 64, pl. XVI, fig. 5 = Merluccius macroactus Weiler, 1935 : 34). Нижние слои среднего олигоцена Карпат (Wola radziszowska).
Merluccius lednevi Bogacev (Богачев, 1933 : 35). Олигоцен — нижний миоцен Апшеронского п-ова (верхнемайкопская свита окрестностей Сумгаита).
Merluccius maicopicus Bogacev (Богачев, 1933 : 26) Нижнемайкопская свита окрестностей Сумгаита (олигоцен? Кавказа).
Merluccius errans (Smirnov) (= Spinogadus errans Смирнов, 1936 : 68 = Merluccius maeoticus Богачев, 1942 : 89). Идентичность Merluccius maeoticus установлена П. Г. Данильченко, которому автор признателен за это сообщение. Merluccius sp. Леднева (1914 : 37) отождествляется Богачевым (1933 : 35) с Merluccius lednevi. По указанию П. Г. Данильченко, он ближе к Merluccius errans. Олигоцен — нижний миоцен Сев. Кавказа (верхнемайкопские отложения окрестностей Дзауджикау и мэотические Таманского п-ова).
Почти все ископаемые виды рода Merluccius из средней Европы после детального исследования оказались принадлежащими к роду Nemopteryz. Точно так же при сравнении и отолитов обоих родов оказалось, что все палеоценовые и эоценовые и большинство олигоценовых отолитов, по которым описан ряд видов рода Merluccius, принадлежат к роду Nemopteryx; к роду Merluccius принадлежат отолиты лишь из верхнетретичных отложений средней Европы (Weiler, 1935 : 35).
Род GADUS L. (s. lat.)
Ныне живущие тресковые, относившиеся ранее к роду Gadus, в настоящее время разделяются на несколько самостоятельных родов. Для ископаемых тресковых, описанных большей частью по несовершенным остаткам, это сделать затруднительно, и поэтому в нижеследующем они отнесены к роду Gadus s. l.
Gadus jonas (De Bosniaski) (in: Sauvage, 1880 : 12 = Gadus jonas Erasmo, 1930 : 78). Верхний миоцен Апеннинского п-ова (Gabbro, пров. Ливорно).
Gadus aeglefinoides (Kner et Steindachner) (= Morrhua aeglefinoides Kner u. Steindachner, 1863 : 34; Kramberger, 1883 : 65). Верхний миоцен Хорватии (Podsused).
Gadus extensus (Kramberger) (= Morrhua extensa Kramberger, 1885 : 51; 1886 : 135). Верхний миоцен Семиградья (Szakadat).
Gadus lanceolatus (Kramberger) (= Morrhua lanceolata Kramberger, 1883 : 67). Верхний миоцен Хорватии (Podsused).
Gadus macropterygius (Kramberger) (= Morrhua macropterygia Kramberger, 1883 : 65 = Gadus sp.? aff. aeglefinoides Богачев, 1928 : 38 = Gadus macropterygius Богачев, 1933 : 33). Верхний миоцен Хорватии (Dolje) и Ашперонского п-ова (диатомовые глины окрестностей Бинагадов).
Gadus minimus (Kramberger) (= Morrhua minima Kramberger, 1885 : 63; 1891 : 104). Верхиий миоцен Хорватии (Sused).
Gadus szagadatensis (Steidachner) (= Morrhua szagadatensis Steindachner, 1863 : 139). Верхний миоцен Семиградья (Szakadat).
Gadus atropatanus Bogacev (Богачев, 1933 : 37). Средний миоцен Апшеронского п-ова (спириалисовые слои).
Gadus kiplingi Bogacev (Богачев, 1928 : 36; 1933 : 40). Верхний миоцен Апшеронского п-ова (диатомовые слои окрестностей Бинагадов).
Gadus caspius Bogacev (Богачев, 1928 : 36; 1933 : 41). Верхний миоцен Апшеронского п-ова (диатомовые слои окрестностей Бинагадов).
Gadus kwitkae Bogacev (Богачев, 1933 : 43; 1938 : 40). Нижний плиоцен Апшеронского п-ова (понтические глины окрестностей Бинагадов и Кировского района).
Gadus spoliatus Bogacev (Богачев, 1938 : 41). Нижний плиоцен Апшеронского п-ова (понтические мергели Кировского района).
Gadus iniistius (Jordan et Gilbert) (= Arnoldina iniistia Jordan et Gilbert, 1919 : 63 = Arnoldina iniistia Jordan, 1921 : 262 = Arnoldites iniistia Jordan, 1923 : 164). Плиоцен южной Калифорнии. Известен лишь один отпечаток с несохранившимися обоими задними спинным и анальным плавниками. Jordan и Gilbert выделяют его в самостоятельный род, близкий к Theragra или Pollachius.
Gadus sp. (Pauca, 1935 : 103). Плиоцен (понтические слои Oltenie).
Gadus sp. (Pauca, 1935а : 228). Миоцен Семиградья (Gemeinde Desmir, Bezirk Cojoecna).
Некоторые из перечисленных видов идентичны, часть их известна по остаткам посредственной сохранности. Кроме того, остатки рода Gadus s. l. известны в виде отдельных частей скелета из верхнего плиоцена южной Великобритании, а мелкие рыбки — из ледниковых или послеледниковых глин Норвегии (Woodward, 1901 : 599). В ледниковых глинах Швеции близ Лунда найдены кости, идентичные с костями ныне живущей Boreogadus saida (Smitt, 1893 : 484), а в ледниковых же глинах близ Карлсборга (около оз. Веттерн) — кости, идентичные с костями Melanogrammas aeglefinus (Munthe, 1910 : 1). К какому семейству принадлежит Eclipes veternus Jordan et Gilbert (= Merriamina ectens Jord. et Gilb.), Eclipes manni Jordan et Gilbert, Eclipes extensus Jord., относимые Jordan'oм в его последней работе (1921 : 260) к семейству Gadidae, решить трудно.
Таким образом, в олигоцене и миоцене Gadidae были представлены теми же родами, что и современные. В настоящее время из ископаемых тресковых не встречается лишь Nemopteryx.
Сем. BREGMACEROTIDAE
Из представителей этого семейства в ископаемом состоянии известен род Bregmaceros.
Род BREGMACEROS THOMPSON
Bregmaceros albyi (Sauvage) (= Podopteryx albyi Sauvage, 1880 : 17 = Podopteryx bosnjaski Sauvage, 1881 : 19 = Erethmophorus sp. Stefano, 1918 : 85 = Bregmaceros albyi Arambourg, 1925 : 25; Erasmo, 1930 : 51). Верхний миоцен Апеннинского п-ова, Сицилии (Ликата) и Алжира (Sidi-Brahim, Raz-el-Ain).
Bregmaceros filamentosus (Priem) (= Problennius filamentosus Priem, 1908 : 17 = Bregmaceros filamentosus Arambourg, 1925 : 58). Эоцен Ирана (Pouchet-e-Kuuh).
Сем. MORIDAE
Представители этого семейства в ископаемом состоянии в виде отпечатков не известны.
Strinsia alata Steindachner (1859 : 771) сюда отнесена быть не может, так как род Strinsia из современных фаунистических сводок исключен. В верхнеолигоценовых (ярус Hutchinsonian) отложениях Новой Зеландии найдены отолиты, которые Frost (1933 : 129) относит к Physiculus [G. (Physiculus) terakohensis].
Таким образом, с самого начала третичного периода семейства Gadidae и Bregmacerotidae, насколько можно судить по ископаемым остаткам, были достаточно резко дифференцированы. Хорошо дифференцированы были и многие современные роды подсемейства Lotinae (Brosme, Lota, Merluccius). Насколько резко были дифференцированы в миоцене современные роды Gadinae, по ископаемым остаткам судить трудно.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ
Географическое распространение каждого из семейств отряда Gadiformes имеет существенные особенности, обусловленные главным образом их экологическими различиями. Представители семейства Moridae, обитающие на значительных глубинах, как и вся основная масса глубоководной фауны, являются космополитами; они довольно равномерно распространены почти во всех глубоких морях (табл. 1).
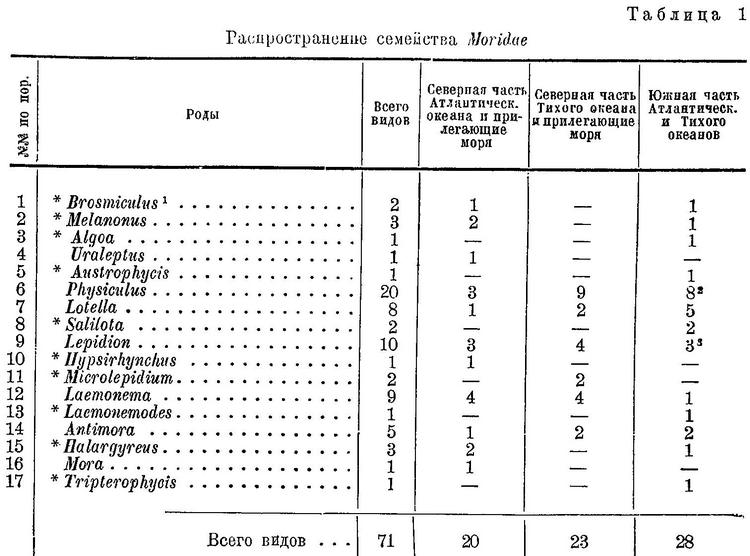
1 Звездочками отмечены роды, не исследованные остеологически и предположительно относимые к семейству Moridae.
2 Включая 3 вида из Индийского океана.
3 Включая 2 вида из Индийского океана.
Распространение теплолюбивых представителей семейства Bregmacerotidae ограничено тропическими и субтропическими морями.
Распространение семейства Gadidae, более холодноводного, чем семейство Bregmacerotidae, и держащегося на значительно меньших глубинах, чем семейство Moridae, ограничено в основном северным полушарием, исключая некоторых представителей родов Gaidropsarus, Urophycis, Merluccius и Micromesistius, встречающихся у берегов Новой Зеландии, у южной оконечности Африки и у Южн. Америки (см. табл. 2 и фиг. 8). В тропической области, где найдены на глубинах лишь единичные представители родов Merluccius ж Urophycis (возможно, есть и Gaidropsarus), представители семейства Gadidae, как правило, не встречаются. В пределах занимаемого ареала не все Gadidae имеют одинаковое распространение: некоторые из них встречаются только в эротических морях, большинство — в умеренных и немногие — в субтропических.
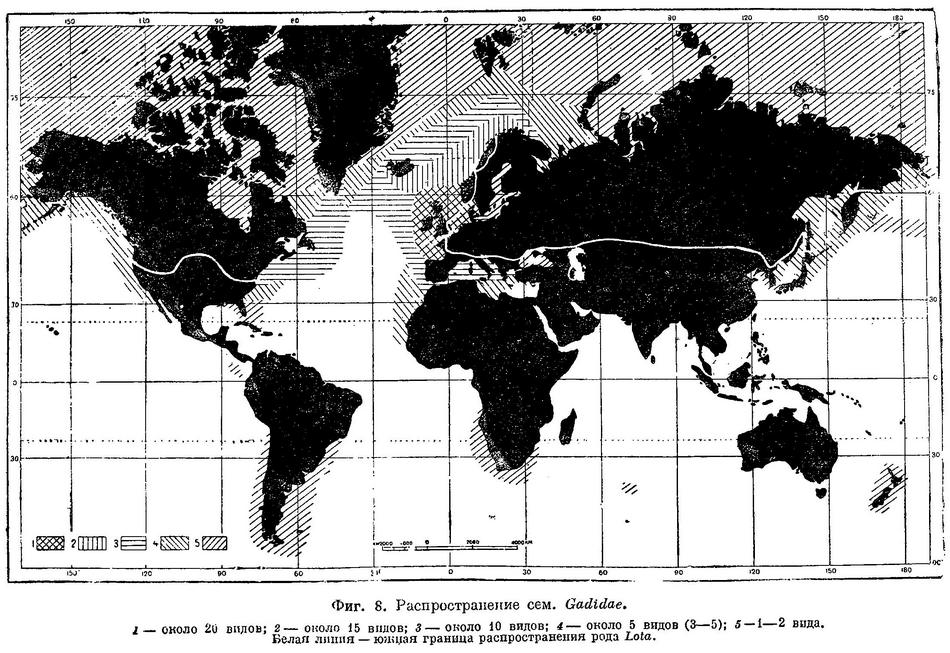
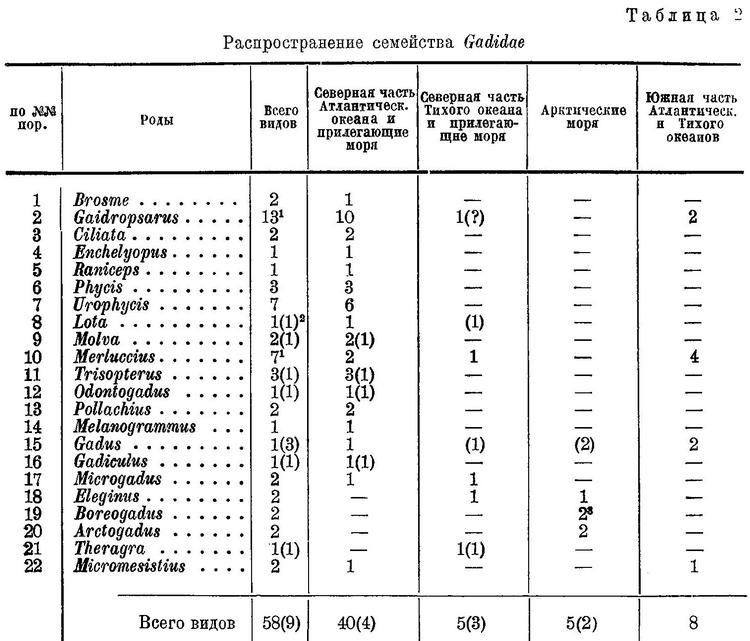
1 Большинство этих видов, несомненно, является только подвидами.
2 В скобках указано количество подвидов.
3 Boreogadus saida встречается в самой северной части Берингова моря, в основном будучи распространенной в арктических морях.
Современное распространение отдельных представителей семейства Gadidae в пределах занимаемого ими ареала, как и распространение отдельных семейств отряда, помимо причин исторических, обусловливается в основном температурой. Всех их можно разбить, на несколько групп соответственно основным зоогеографическим подразделениям Ortmann'a (1896) (Звездочками обозначены формы, держащиеся на значительных глубинах.):
I. Арктическо-циркумполярная подобласть: Arctogadus borisovi, Arctogadus pearyi, Boreogadus agilis, Boreogadus saida, Eleginus navaga, Gadus morhua ogac, Gadus morhua maris-albi, Gadus morhua morhua n. hiemalis. Сюда же следует отнести, повидимому, и Gadus morhua kildinensis из оз. Могильного на о. Кильдине, точно так же, как и прибрежную мурманскую расу Gadus morhua morhua.
II. Атлантическо-бореальная подобласть: Brosme brosme*, Gaidropsarus mediterraneus (Другие встречающиеся здесь сомнительные в видовом отношении представители Gaidropsarus не упомянуты.), Ciliata septentrionalis, Ciliata mustela, Enchelyopus cimbrius, Raniceps raninus, Phycis phycis, Phycis blennoides, Phycis chesteri, Urophycis regius, Urophycis tenuis, Urophycis chuss, Urophycis floridanus, Urophycis earlli, Urophycis cirratus, Lota lota lota, Lota lota leptura, Lota lota maculosa (пресноводные), Molva molva*, Molva dipterygia dipterygia*, Molva dipterygia elongata*, Merluccius merluccius*, Merluccius bilinearis*, Trisopterus luscus*, Trisopterus minutus minutus, Trisopterus minutus capelanus, Trisopterus esmarkii, Odontogadus merlangus merlangus, Odontogadus merlangus euxinus, Pollachius pollachius, Pollachius virens, Melanogrammus aeglefinus, Gadus morhua morhua, Gadiculus argenteus argenteus*, Gadiculus argenteus thori*, Microgadus tomcod, Micromesistius poutassou*.
III. Тихоокеанско-бореальная подобласть: Gaidropsarus pacificus, Merluccius productus*, Gadus morhua macrocephalus, Eleginus gracilis, Theragra chalcogramma chalcogramma, Theragra chalcogramma chalcogramma n. fucensis, Microgadus proximus.
IV. Антарктическая (умеренная) область: Gaidropsarus capensis, Gaidropsarus novae-zealandiae, Urophycis brasiliensis, Merluccius capensis*, Merluccius gayi*, Merluccius australis*, Merluccius hubbsi*, Micromesistius australis*.
В первую группу включены формы, распространение которых лежит в пределах арктическо-циркумполярной подобласти Ортманна; все эти формы, кроме подвидов Gadus morhua и за возможным исключением Arctogadus реаryi и Boreogadus agilis, в значительной степени евригалинны. Южная граница распространения этой группы рыб (Boreogadus saida, Eleginus navaga) в Атлантическом океане ближе совпадает с границей арктической области, очерченной Broch'oм (1933 : 17), чем с границей Ортманна. Некоторые из видов арктической области (род Arctogadus, Boreogadus agilis) населяют самые холодные моря и могут быть названы высоко-арктическими (Hofsten, 1915 : 209). Сравнительно небольшое количество видов, встречающихся как в этой, так и в других подобластях, не дает возможности для дальнейшего их разделения.
Распространение второй группы рыб соответствует атлантическо-бореальной подобласти Ортманна с той лишь разницей, что сюда следует включить Средиземное и Черное моря, и, кроме того, южная граница ее у восточных берегов Атлантического океана простирается несколько южнее — приблизительно до 15° с. ш., что близко совпадает с южной границей распространения средиземноморско-атлантической фауны S. Ekman'a (1935 : 113). У берегов Сев. Америки граница распространения этой группы тресковых почти совпадает с южной границей атлантическо-бореальной подобласти Ортманна. Правильнее говоря, как справедливо отмечает Ekman (1935 : 144) и что в особенности относится к тресковым, это будут не границы, а скорее переходные области, так как в зависимости от сезонных колебаний температуры некоторые виды со струями Гольфштрема далеко проникают в арктическую область. Ряд видов, в особенности держащихся на значительных глубинах, при соответствующих условиях проникает в воды к югу от указанных границ. Сказанное относится к границам и между другими областями.
Выделять средиземноморских тресковых в особую средиземноморско-атлантическую фауну (Ekman, 1935 : 113) с прилегающей к ней частью атлантической оснований нет, так как многие средиземноморские тресковые (Micromesistius poutassou, Phycis blennoides, Trisopterus luscus) за единичными исключениями (Odontogadus merlangus euxinus и Trisopterus minutus capelanus) распространены в Атлантическом океане далеко на север — до границ с арктической фауной и являются здесь более обычными формами, чем в Средиземном море. Кроме того, нерестилища всех видов тресковых находятся в пределах занимаемых ими областей, между тем как все средиземноморские тресковые, за исключением указанных выше двух подвидов, мечут икру не только в Средиземном море, но, главным образом, в Атлантическом океане. Наиболее существенный элемент средиземноморско-атлантической фауны — родство с индо-вестпацифической — фауне тресковых чужд. Некоторые указания на небольшую обособленность тресковых здесь все же имеются: ареалы распространения некоторых видов (Molva dipterygia elongata, Phycis phycis, Gadiculus argenteus argenteus) в известной степени совпадают с границами распространения средиземноморско-атлантической фауны и, кроме того, южная граница некоторых видов (Gadus morhua morhua, Melanogrammus aeglefinus, Pollachius virens, Molva molva, Molva dipterygia dipterygia и др.), встречающихся в северной части Атлантического океана, совпадает довольно близко с северной границей средиземноморско-атлантической фауны Ekman'a.
Третья группа тресковых должна быть отнесена к тихоокеанско-бореальной подобласти Ортманна, граница которой с арктическо-циркумполярной подобластью по тресковым в Беринговом море проходит приблизительно от мыса Чукотского к м. Ном на Аляске. К северу от этой границы заходят треска (Gadus morhua macrocephalus) и вахня (Eleginus gracilis), к югу — сайка (Boreogadus saida). Южная граница распространения этой группы тресковых близко совпадает с границами Ортманна: до средней Калифорнии у берегов Сев. Америки и средней Японии в Тихом океане. К югу от этой границы — в Желтое море и Калифорнийский залив — проникают лишь единичные виды тресковых.
Характерной особенностью этой подобласти является то, что при значительной холодноводности некоторых ее морей фауна тресковых носит здесь отчасти бореальный характер. Представитель арктического рода Eleginus — Eleginus gracilis является здесь более теплолюбивым видом, встречаясь даже в Желтом море. По сравнению с атлантическо-бореальной подобластью фауна тресковых тихоокеанско-бореальной подобласти значительно беднее.
В южном полушарии фауна тресковых обеднена еще более: здесь встречаются пять близких к бореальным видов, или, возможно, даже только подвидов от них из родов Gaidropsarus и Merluccius и по одному виду родов Urophycis и Micromesistius.
Таким образом, наиболее богата фауна тресковых в атлантическо-бореальной подобласти, имеющей наибольшее количество и эндемичных родов. Здесь встречаются наиболее крупные по размерам и наиболее многочисленные по количеству особей виды тресковых. Следующей по количеству эндемичных родов и видов является арктическо-циркумполярная подобласть. Тихоокеанско-бореальная подобласть менее богата тресковыми; эндемичный род здесь только один (Theragra). Антарктическая (умеренная) область не имеет ни одного эндемичного рода.
С зоогеографическими областями Ekman'a (1935) такое распределение тресковых согласуется менее прежде всего потому, что выделять в особую средиземноморско-атлантическую область тресковых, встречающихся в Средиземном море, мало оснований. Кроме того, тресковых Берингова и Охотского морей с большим правом следует отнести к тихоокеанско-бореальной области, а не к арктической. Наконец, однородную и бедную фауну тресковых южного полушария правильнее считать как единую, не разделяя на южноафриканскую, южноавстралийско-новозеландскую и перуанско-северочилийскую области.
Почти для всех групп морских животных, в частности и для рыб, многими авторами (Günther, 1880 : 176; Шмидт, 1904 : 394; Берг, 1918 : 1835; Broch, 1935 : 42 и др.) указывалось, что фауна северных частей Тихого океана, имея большое сходство с фауной северных частей Атлантического океана, значительно богаче ее видами, родами и даже семействами. Фауна северных частей Атлантического океана, а также и фауна арктических морей, в основном является обедненной фауной северных частей Тихого океана и ведет свое начало, главным образом, от этой последней.
Среди рыб почти единственным исключением из этого общего правила является семейство тресковых, которое, наоборот, большим количеством видов и родов представлено в северных частях Атлантического океана. Причины этого своеобразного распространения тресковых, стоящего в противоречии с распространением остальных рыб и обратившего на себя внимание одновременно с обнаружением сходства фаун северных частей Атлантического и Тихого океанов и значительно большего богатства последней (Günther, 1880 : 180; Шмидт, 1904 : 402), до сего времени все же остаются не выясненными. После выделения из семейства Gadidae представителей семейства Moridae, которые, как глубоководные рыбы, являются космополитами, это преимущественно атлантическое распространение тресковых становится еще более резко выраженным (см. табл. 1 и 2 и фиг. 8).
Численное преобладание видов и родов тресковых в северных частях Атлантического океана, наличие здесь ряда эндемичных родов (Brosme, Raniceps, Phycis, Urophycis, Molva и др.), отсутствие самобытности тихоокеанской фауны тресковых и явная генетическая зависимость от атлантической с несомненностью говорят о происхождении тихоокеанских тресковых от атлантических, — мнение, к которому в последние годы приходят многие исследователи (Scheuring, 1930 : 197; Зенкевич, 1933 : 21; Шмит, 1936 : 175). Очевидно современная фауна тресковых Атлантического и Тихого океанов и арктических морей имеет иное происхождение, чем остальная ихтиофауна этих бассейнов.
По мнению Л. А. Зенкевича, богатство видов и родов тресковых в Атлантическом океане есть явление, сложившееся в самом Атлантическом океане и произошедшее уже «после имевшего место разъединения двух океанов в конце плиоцена» (l. с. : 21),— мнение, стоящее в противоречии с палеонтологическими данными, так как остатки тресковых найдены значительно ранее — в олигоцене и миоцене, а отолиты — начиная с верхнего палеоцена. К этому необходимо добавить, что и эволюция тресковых не происходила такими быстрыми темпами, так как уже в олиго- и миоценовых отложениях западной Европы и Кавказа тресковые, в особенности из подсемейства Lotinae, представлены теми же родами, что и современные; многие из этих родов представлены несколькими видами, а в ледниковых глинах Скандинавского п-ова найден скелет небольшой трески, который идентичен со скелетом современной Boreogadus saida. Некоторые авторы (Шмит, 1936 : 175) считают, что основным бассейном, в котором развивались в третичное время тресковые, были моря, находившиеся в это время на месте северной части Атлантического океана и далее к северу.
При решении вопроса об истории происхождения фауны тресковых, помимо их современного географического распространения, необходимо принять во внимание: 1) явную и достаточно резко выраженную холодолюбивость их при одновременной евритермности всего семейства в целом, 2) время и места нахождения их в ископаемом состоянии и 3) геологическую историю мест нахождения ископаемых остатков тресковых.
На преимущественно холодолюбивый характер тресковых обращали внимание почти все авторы (Günther, 1880 : 177: Зенкевич, 1933 : 21; S. Ekman, 1935 : 235), отмечавшие их своеобразное распространение. Действительно, и географическое распространение (преимущественно в довольно высоких широтах северного полушария) и, что самое существенное, время нереста, приходящееся у громадного большинства видов на холодное время года, говорят, несомненно, о холодолюбивости тресковых. Условия размножения, вообще говоря, как указывает Appellöf (1912 : 303), могут служить косвенным указанием на происхождение, причем размножение в холодное время года указывает на арктическое происхождение. У теплолюбивых и южнее распространенных рыб время нереста приходится на самое теплое время года. Scomber scombrus (L.), приблизительно в тех же широтах, что и треска (у берегов Норвегии), нерестится в июне — июле (Mcintosh a. Masterman, 1897 : 160), Mullus barbatus L. и Mullus surmuletus L. у берегов Великобритании мечут икру в июне — июле (Meek, 1916 : 304). В более южных широтах нерест этих рыб происходит раньше. Время нереста у камбал, среди которых есть группы как бореальных и даже почти арктических, так и тропических рыб, различно в зависимости от распространения: тропические камбалы мечут икру в теплое время года, нерест камбал с более северным распространением, приходится на более холодное время года; но и среди них лишь некоторые, встречающиеся в наиболее высоких широтах, из подсемейства Pleuronectinae нерестятся в самое холодное время года. Внутри самого семейства тресковых виды, обладающие самым южным распространением, размножаются в более теплое время года, чем виды, распространенные в более северных широтах (J. Schmidt, 1909 : 155).
В ископаемом состоянии представители семейства тресковых в виде несомненных остатков — отпечатков скелетов — известны из средней Европы, Кавказа и немногие из Калифорнии (См. главу «Ископаемые тресковые».). Впервые в нижнем олигоцене появляется Nemopteryx, который к концу среднего олигоцена исчезает. В средне- и верхнеолигоценовых отложениях появляются глубоководные представители тресковых — Brosme и Merluccius. Тресковые типа Gadus s. l. впервые появляются в более поздних отложениях, единичные остатки — в среднем миоцене и многочисленные — в верхнем миоцене. Из нижнеплиоценовых отложений известны Phycis и Lota. Судя по нахождениям отолитов, тресковые встречались и в более раннее время — в эоцене и даже в верхнем палеоцене, но что это были за тресковые, сказать, как указывалось выше, трудно, хотя их и относили ко многим родам и даже видам современных тресковых. Все эти ископаемые остатки найдены в отложениях средней Европы и Кавказа. Из области Тихого океана (южная Калифорния) ископаемые тресковые известны только из плиоценовых отложений.
Ко времени, к которому относятся найденные первые ископаемые остатки тресковых — в палеогене и ранее, на месте современного Средиземного моря и южной и средней Европы, как известно, простиралось море Тетис, продолжавшееся далее на запад на месте средней части современного Атлантического океана и на юго-восток, где оно стояло в связи с Индийским океаном. Северная граница Тетиса в Европе тянулась вдоль северной границы Карпат и Альп, которые возвышались в виде плоских островов; на юге граница его проходила приблизительно вдоль северной Африки. Через северную часть современного Атлантического океана простирался мост, соединявший Европу через Исландию и Гренландию с Сев. Америкой (Jhering, 1927 : 35).
Палеогеновая фауна Тетиса была тропической, очень богатой и, по взглядам Ekman'a (1934 : 359), дала начало современной литоральной фауне тропических морей. Палеоценовые, эоценовые и олигоценовые рыбы Тетиса и прилегающих к нему англо-парижского и бельгийского эпиконтинентальных бассейнов были точно так же тропическими (Leriche, 1902 : 47; 1905 : 49; 1910 : 231); современных тресковых здесь не было. Лишь со середины и конца олигоцена в Тетисе появляются современный глубоководный Merluccius, а в северо-восточной части его (Апшеронский п-ов) Brosme; для держащихся на меньших глубинах тресковых типа Gadus s. l. климат, очевидно, был еще слишком теплым. Встречающийся только в олигоценовых отложениях Nemopteryx был, повидимому, немного более теплолюбивым, чем современные тресковые, и жил, вероятно, на глубинах, так как остатки его находят главным образом в глубоководных отложениях областей, занимавшихся Тетисом; лишь Nemopteryx emarginatus найден в более мелководных отложениях Майнцского бассейна.
С середины третичного периода (в миоцене) Тетис значительно сокращается; появляется ряд островов, северо-восточная часть его отделяется и впоследствии значительно опресняется, пролив через Пиренейский п-ов исчезает, уступая место узкому проливу в области р. Гвадалквивира, утрачивается связь с Индийским океаном. Вследствие похолодания климата европейская часть Тетиса теряет свою тропическую фауну.
Таким образом, на протяжении конца третичного периода происходит постепенное похолодание климата, под влиянием чего палеогеновая тропическая фауна Тетиса исчезает и на смену ей приходит холодноводная фауна, ведущая свое начало из северных частей Тихого океана. Проникновение этой фауны в Атлантический океан оказалось возможным впервые, как показал Берг (1918 : 1835), лишь в плиоцене, когда наряду с трансгрессией в области Берингова пролива был более теплый климат. Тресковые, как указывалось выше, появились в тех морях, которые были на месте Атлантического океана и Европы, значительно ранее этого времени. Уже в среднемиоценовых отложениях Кавказа появляется представитель рода Gadus s. l. (Gadus atropatanus), а из верхнемиоценовых отложений известны многие представители этого рода, кроме Кавказа, и из средней Европы. В верхнемиоценовых же отложениях средней Европы и Алжира появляется и Brosme, известный ранее из олигоцена Кавказа. В Тихом океане тресковые появляются лишь в плиоцене (Калифорния), т. е., повидимому, после трансгрессии в области Берингова пролива.
Все изложенные выше факты — современное распространение тресковых, их экология, места и время нахождения их в ископаемом состоянии и третичная история тех морей, в отложениях которых найдены их остатки, — говорят о том, что при верхнетретичном похолодании климата тресковые пришли в моря, бывшие на месте современной средней Европы и Кавказа, из более холодноводных бассейнов — откуда-то с севера. В каких северных бассейнах выработалась фауна тресковых и где они были в раннетретичное время, на основе современных знаний истории этих бассейнов можно только догадываться и высказывать предположения. Все же можно сказать, что фауна тресковых сложилась не в тех эпиконтинентальных морских бассейнах, которые были на месте современной западной Европы (англо-парижский, бельгийский и др.), хотя в их отложениях и найдены отолиты, о принадлежности которых если не к родам, то, безусловно, к семейству тресковых утверждать можно, так как климат, флора (Köppen-Wegener, 1924 : 104) и фауна этих мест, как водная (Leriche, 1902 : 47; 1905 : 49; 1906 : 125; 1910 : 231; White, 1931 : 36), так и наземная, были тропическими. Очевидно, в раннетретичное время тресковые держались в более холодноводном бассейне и к тропической фауне примешивались лишь наиболее глубоководные из них; это происходило в местах, находившихся, повидимому, в сфере действия какого-либо идущего из этого бассейна холодного течения в придонных слоях воды, где вообще держатся тресковые. Таким бассейном мог быть Полярный и прилегающие к нему Скандский и другие бассейны, но к сожалению, третичная история Полярного бассейна оставалась до последнего времени не изученной, хотя он и изображается на всех (46—62) палеогеографических картах Arldt'a третичного, мелового, юрского и и триассового периодов. Древность Полярного бассейна в сущности подтверждалась только тетраэдрической теорией строения земли, согласно которой вокруг Северного полюса всегда было море (Arldt, 1919 : 1196). По новейшим исследованиям Д. Г. Панова (1946), формирование Полярного бассейна и установление в нем морского режима относится к мезозою с самого начала третичного периода началось углубление и формирование современной впадины.
Существенным подтверждением высказанных соображений о происхождении тресковых может служить распространение некоторых высших ракообразных. Так, например, помимо значительного количества видов из разных групп водных животных (см. об этом Ekman, 1935 : 245), есть несколько родов Amphipoda с почти или исключительно высокоарктическим распространением — «высокоарктические автохтонные формы», как их называет Гурьянова (Gurjanova, 1935 : 566). К числу их принадлежит род Onisimus с 11 чисто арктическими видами, из которых в северную часть Атлантического океана спускаются только три; в северной части дальневосточных морей представители этого рода отсутствуют (Гурьянова, 1932 : 159). Чисто арктическими являются все три вида рода Acanthostepheia (Гурьянова, 1929 : 288; 1932 : 159). Из шести видов рода Pseudolibrotus четыре являются исключительно арктическими, едва спускающимися ниже Полярного круга в северную часть Атлантического океана и отсутствующими в Тихом океане; два вида встречаются в Каспии (Гурьянова, 1932 : 163). Из Isopoda чисто арктическим является род Mesidothea, из четырех видов которого лишь один (Mesidothea entomon) имеет подвиды в Балтийском, Беринговом, Охотском и Каспийском морях и в некоторых пресноводных водоемах северной Европы и Сев. Америки (Гурьянова, 1932 : 175). Число эндемичных ракообразных для Полярного бассейна крайне велико: среди Amphipoda — 41%, среди Isopoda — 34% (Гурьянова, 1939 : 681).
Распространение перечисленных ракообразных имеет большое сходство с распространением тресковых: лишь незначительная часть их (Mesidothea entomon orientatis), пропорционально значительно меньше части тресковых, встречается в северной части дальневосточных морей, будучи в основном распространена в арктических морях и в северной части Атлантического океана, причем единственный тихоокеанский представитель этих ракообразных ведет начало от арктического, подобно тому как тихоокеанские тресковые ведут начало от атлантических. Но, помимо сходства, в распространении тресковых и этих ракообразных есть и различия: большая часть ракообразных встречается в арктических морях, едва опускаясь в северную часть Атлантического океана и почти отсутствуя в дальневосточных морях, между тем как тресковые более многочисленны и встречаются много южнее и в Атлантическом и в Тихом океанах и, что самое главное, представлены большим многообразием в северной части Атлантического океана, а не в арктических морях.
Причина этого сходства, которое в раннетретичную эпоху, можно полагать, было еще большим, обусловлено, несомненно, тем, что тресковые и перечисленные ракообразные выработались в одном и том же, повидимому, Полярном бассейне. Различие же в их распространении произошло позднее и является следствием верхнетретичного похолодания климата и большей подвижности тресковых. Как указывалось выше, климат и фауна Тетиса и расположенных к северу от него эпиконтинентальных бассейнов были тропическими. Несомненно, климат и в области Полярного бассейна был значительно теплее, чем современный, так как палеогеновая флора Гренландии, Исландии и Шпицбергена была даже несколько более южного типа, чем флора современной Норвегии; здесь найдены два вида берез, липа, сосна и др. (Нееr, цитировано по Köppen-Wegener, 1924 : 106). С похолоданием климата тресковые стали из Полярного бассейна отходить и, найдя для себя наиболее подходящие условия южнее, сконцентрировали здесь все свое многообразие, оставив в арктических морях наиболее холодолюбивые элементы. Упомянутые же ракообразные, как менее подвижные и, несомненно, более холодолюбивые, чем основная масса тресковых, остались в арктических морях. Вследствие более раннего соединения Полярного бассейна с Атлантическим, а не с Тихим океаном, тресковые могли при похолодании отойти в северную часть первого, а не последнего. Этим же, повидимому, объясняется большее проникновение в северную часть Атлантического океана, а не Тихого, и упомянутых выше ракообразных. Как известно, обмен фаунами между Тихим океаном и арктическими морями, а через них и с Атлантическим океаном, стал возможен лишь в плиоцене. Соединение Полярного бассейна с Тетисом началось значительно ранее. С самого начала третичного периода и до конца олигоцена (Архангельский, 1935 : 119; Обручев, 1935 : 9,11) на северо-восток от Тетиса, от современного Кавказа по направлению к устьям Оби и Енисея простирался широкий морской рукав — море, по которому, возможно, и проникали на юг олигоценовые представители родов Brosme и Merluccius, находимые в нижнемайкопских отложениях Кавказа. В эоцене это море было мелким, в олигоцене — довольно глубоким, а в конце его оно распалось на лагуны и озера (Обручев, 1935 : 8—9). Кроме того, мост, соединявший Европу через Великобританию, Исландию и Гренландию с Сев. Америкой, по мнению большинства палеогеографов, существовал не постоянно (см. Arldt, 1919 : 406—423), а временами, повидимому, он был покрыт морем, образующим на нем заливы или проливы, соединявшие Полярный бассейн с Тетисом и прилегавшими к нему морями. Вероятнее всего, что тресковые, отолиты которых известны из нижнетретичных отложений западной Европы, именно таким путем и проникали в эти места. Повидимому, как замечает Davies (1920 : 311), англо-парижско-бельгийский бассейн в палеоцене (ярус Landenien) был подвержен влияниям с севера и с юга (Тетиса), с последнего, повидимому, менее, благодаря чему в нем присходило смещение теплолюбивой и холодолюбивой фаун; поэтому здесь и встречаются отолиты тресковых. Вообще же для концентрации тресковых в высоких широтах в палеогене достаточно было господствовавших тогда климатических условий и лишь частичной изоляции Полярного бассейна. Основные миграции тресковых из Полярного бассейна в средние широты происходили, очевидно, в верхнем миоцене, когда суша между Европой и Сев. Америкой исчезла и климат стал почти современным. В это время, как указывалось выше, в отложениях средней Европы и Кавказа появились многочисленные представители рода Gadus s. l. и значительно большее количество, чем в олигоцене, представителей других родов тресковых.
Основные черты современного распространения тресковых, как и всей фауны и флоры, сложились в начале четвертичного периода, когда под влиянием похолодания и развития мощных оледенений, в высокоарктической области сконцентрировались наиболее холодолюбивые элементы тресковых, а все более теплолюбивые принуждены были уйти южнее. При этом, благодаря периодическим опреснениям и осолонениям, происходившим в это время в Полярном бассейне, из холодолюбивых тресковых, как указывает Зенкевич (1933 : 22), в высокоарктической области выжили лишь те, которые смогли переносить опреснение (роды Arctogadus, Boreogadus, Eleginus). Ряд других холодолюбивых, но не переносящих опреснения тресковых (роды Gadiculus, Brosme и др.), принужден был вместе с более теплолюбивыми уйти в более южные широты, но на глубины.
Для концентрации в разных областях различных экологических групп тресковых, повидимому, необязательна была даже и полная изоляция их друг от друга на это время при помощи сплошного моста между Европой и Сев. Америкой. Для этого достаточно было и одного ледникового похолодания и сопутствующего ему хотя бы небольшого опреснения что возможно и не при полном отделении арктических морей от Атлантического океана. Следует, однако, заметить, что, судя по всему, полного опреснения в Полярном бассейне не было, так как иначе здесь не могли бы сохраниться как перечисленные тресковые, размножающиеся только в морской воде, так и упомянутые ракообразные; (см. об этом также у Гурьяновой, 1939 : 679). Переход налима к пресноводному образу жизни происходил, повидимому, в течение долгого периода времени и в разных местах его распространения. Хотя в ископаемом состоянии налим известен из пресноводных или солоноватоводных отложений окрестностей Вены из нижнего плиоцена, но в основном в пресную воду он ушел, повидимому, в ледниковое время и из Полярного бассейна. В пользу этого предположения говорит его современное циркумполярное распространение и отсутствие в неогеновых (плиоценовых) отложениях Сибири (Штылько, 1934 : 74).
В Тихий океан тресковые могли проникнуть, как указывалось выше (см. об этом Берг, 1918 : 1838; 1934 : 69), в плиоцене и затем в послеледниковую трансгрессию, когда было время с более теплым климатом, чем современный (Берг, 1911 : 73; Smith, 1919 : 167). Несомненные остатки тресковых известны из плиоценовых отложений Калифорнии. Повидимому, в плиоцене появилась в Тихом океане Theragra, обособившаяся до степени самостоятельного рода. Когда проникли в Тихий океан остальные тресковые, сказать еще труднее. Относительно этого существуют две точки зрения. По одной из них (Шмит, 1936 : 175), треска и навага в Тихий океан проникли в плиоцене, по другой (Шмидт, 1936 : 136) — перед или во время ледникового похолодания. В пользу первого предположения как будто бы говорит их современное распространение в Тихом океане далеко на юг — до Желтого моря включительно, куда они могли уйти, казалось бы, только под влиянием какого-либо похолодания. Однако это не могло произойти потому, что Желтое море вследствие поднятия суши в начале четвертичного периода само представляло сушу, а Японское море — замкнутый бассейн. Ни в этом замкнутом бассейне, ни в прилегающих к нему частях Тихого океана тресковых в это время быть не могло, вследствие значительно более теплого, чем современный, климата, о чем говорит нахождение тропической фауны (моллюски и кораллы) в бухте Токио в номскую фазу (Криштофович, 1932 : 293). В Желтое и Японское моря треска и навага могли проникнуть, попав в Тихий океан даже в послеледниковое время, так как известно, что, например, треска, принадлежащая, правда, к другой, более мигрирующей форме, может в очень короткий срок при изменении термических условий появиться там, где она раньше не была известна; так, в последние годы, в связи с общим потеплением, треска была обнаружена в Карском море (Берг, 1935 : 73). Таким образом, более вероятно, что треска и навага проникли в Тихий океан в послеледниковое время. Беломорская треска (Gadus morhua maris-albi), поселившаяся в Белом море в послеледниковое время (Дерюгин, 1932 : 443), является, подобно тихоокеанской, точно так же подвидом. Это тем более вероятно еще и потому, что навага и та форма трески, от которой произошла тихоокеанская треска, довольно холодолюбивы. Повидимому, одновременно с треской и навагой проникли в Тихий океан Microgadus proximus и Merluccius productus, имеющие очень близкие виды в Атлантическом океане. С другой стороны, большая теплолюбивость их, чем трески и наваги, говорит о том, что расселение их могло произойти и в другое время, а у Merluccius и иным путем (см. ниже). Значительно позднее, повидимому, по окончании теплого послеледникового времени, появилась в Беринговом море сайка (Boreogadus saida), держащаяся в самых северных частях его; причиной медленного распространения ее южнее, в частности в Охотское море, где она нашла бы для себя подходящие условия, является, повидимому, отсутствие южнее мыса Лопатки вдоль восточного берега Камчатки и у Алеутских о-вов пловучих льдов, с которыми обычно теснейшим образом связана эта рыба. Наиболее холодолюбивый из тресковых род Arctogadus держится в самых холодных из арктических морей.
Современное распространение тихоокеанских тресковых и близких к ним систематически арктических и атлантических видов и подвидов дает указание и на те пути, по которым они проникли в Тихий океан. Распространение Eleginus navaga вдоль северных берегов Европы и Сибири и Eleginus gracilis в дальневосточных морях непосредственным образом указывает на проникновение последней в Тихий океан вдоль берегов Азии. Microgadus proximus (близкий к ней вид Microgadus tomcod встречается у атлантических берегов Сев. Америки), несомненно, прошла в Тихий океан кругом Сев. Америки. Этим же путем прошла в Тихий океан и Gadus morhua macrocephalus, что подтверждается не столько количеством позвонков (Шмидт, 1936 : 64), сколько ее близостью, как краниологической, так и экологической, с Gadus morhua ogac, встречающейся у берегов Гренландии. Gadus morhua ogac и Gadus morhua macrocephalus, характеризующиеся широкоголовостью и, кроме того, экологически тем, что они не совершают подобно узкоголовой Gadus morhua morhua отдаленных миграций, в доледниковое время представляли, повидимому, одну форму, которая занимала один общий обширный ареал. С исчезновением суши между Сев. Америкой и Европой сплошной ареал этой прибрежной формы оказался прерванным образовавшимися на месте суши глубинами и, таким образом, в северных частях у обоих берегов вновь образовавшегося Атлантического океана оказались две формы; первоначально изолированные географически, они затем приобрели и морфологические отличия. Форма, изолированная у северных берегов Европы, повидимому, играла роль в образовании Gadus morhua maris-albi. Форма, державшаяся у берегов Сев. Америки, проникнув в Тихий океан, дала начало Gadus morhua macrocephalus, которая, широко расселившись здесь к югу, образовала более теплолюбивые, чем исходная форма, расы. Узкоголовая форма трески — Gadus morhua morhua, распространенная южнее и более теплолюбивая, не проникла в Тихий океан вместе с широкоголовой треской и навагой, вероятно, потому, что климат в это время у северной оконечности Сев. Америки оказался для нее слишком суровым. Вряд ли таким же путем проник в Тихий океан и Merluccius productus; для него, как для глубоководной рыбы, наиболее вероятен другой путь — от берегов Южн. Америки, в пользу чего отчасти говорит отсутствие его у азиатских берегов, а главное — современное биполярное распространение. Boreogadus saida, встречающаяся в арктических морях у берегов и Азии, и Сев. Америки, держится и в северных частях Берингова моря у обоих берегов. Таким образом, судя по современному распространению, расселение тресковых в Тихий океан происходило, повидимому, главным образом вдоль берегов Сев. Америки, т. е. совпадает с путями расселения большинства представителей других групп водных животных, мигрировавших преимущественно в обратном направлении из северных частей Тихого океана в северную часть Атлантического.
Расселение тресковых в южное полушарие, где они представлены близкими, если не идентичными видами или подвидами родов Gaidropsarus (Gaidropsarus capensis, Gaidropsarus novae-zealandiae) и Merluccius (Merluccius capensis, Merluccius hubbsi, Merluccius australis, Merluccius gayi) и единичными видами родов Micromesistius (Micromesistius australis) и Urophycis (Urophycis brasiliensis) у южной оконечности Африки и Южн. Америки и у Новой Зеландии, могло произойти во время ледникового похолодания, а также отчасти может происходить даже и в настоящее время. Как известно, в тропической области поверхностный слой воды с максимальной температурой воды в широтном направлении наиболее узок, а придонный слой с минимальной температурой наиболее широк в Атлантическом океане у западных берегов Африки (Schott, 1926 : 134, tabl. IX, XIV и XV) и в Тихом океане у западных берегов Америки (Schott, 1935 : 150, 181, tabl. XX—XXIII и XXXI—XXXII). Благодаря этому переход от тропической фауны к умеренной здесь наименее резок (Ekman, 1935 : 88) и вследствие этого здесь же имеются наилучшие условия для обмена фаунами между северным и южным полушариями. Так, например, отдельные представители биполярно распространенного сем. Triglidae (Световидов, 1936 : 8) иногда были находимы в тропической области у западных берегов Африки (Regan, 1915 : 129). В тропической области в Тихом океане, у западных берегов Америки, на глубинах, как указывалось выше, найдены и некоторые тресковые (Merluccius gayi). Таким образом, придонные слои воды в тропической области не представляют непреодолимой преграды для расселения на юг более теплолюбивых (некоторые виды рода Gaidropsarus) и глубоководных (роды Merluccius и Micromesistius) тресковых в указанных местах и в настоящее время, но в основном для большинства из них это расселение произошло во время ледникового похолодания. Вообще же в южное полушарие проникли виды из наиболее теплолюбивых и с более южным распространением родов (Gaidropsarus, Urophycis) и из живущих на больших глубинах и ведущих в значительной мере батипелагический образ жизни (Merluccius, Micromesistius).
Экологические особенности и тесно связанное с этим распространение семейств Gadiformes по климатическим поясам были такими же, как и современные, с самого начала третичного периода. Представители семейства Bregmacerotidae были тропическими в эоцене (Bregmaceros filamentosus, эоцен Ирана), примерно такими же оставались они в миоцене (Bregmaceros albyi, верхний миоцен Алжира и Сицилии); в тропической области держатся они и в настоящее время. Миоценовое похолодание климата влечет за собой приток в средние широты северного полушария более холодноводных, ранее к роду Physiculus и найденным в верхнеолигоценовых отложениях Новой Зеландии (Frost, 1933 : 129), семейство Moridae имело широкое распространение точно так же с самого начала третичного периода.
БИОЛОГИЯ
(Составлено по данным систематической части и работам Hjort'a (1909), Damas'a (1909), J. Schmidt'a (1909) и некоторых других, цитируемых в тексте, авторов.)
Все Gadiformes, за исключением одного вида, — морские рыбы; в особенности относится это к семействам Moridae и Bregmacerotidae, представители которых, как правило, не встречаются даже в опресненных предустьевых пространствах рек. Что касается семейства Gadidae, то среди представителей его имеется один вид — налим с тремя подвидами (Lota lota lota, Lota lota leptura, Lota lota maculosa), живущий и размножающийся в пресной воде. Ряд видов семейства Gadidae, будучи в основном морскими рыбами, заходит в сильно опресненную и даже совсем пресную воду, причем некоторые поднимаются в сфере влияния приливов и отливов на значительные расстояния в реки. К числу этих видов относятся Eleginus navaga, Eleginus gracilis, Microgadus tomcod и, возможно, Microgadus proximus, Boreogadus saida, Arctogadus borisovi и некоторые другие. Из числа исключительно морских видов семейства некоторые (например, Gadus morhua и в особенности некоторые ее формы) способны переносить несколько пониженные солености даже в период размножения, а иногда случайно могут заходить и в совершенно пресную воду (3 экземпляра трески пойманы в р. Луге в 1 км от устья). Ряд представителей семейства (Molva molva, Molva dipterygia dipterygia, Molva dipterygia elongata, Gadiculus argenteus argenteus, Gadiculus argenteus thori, Micromesistius poutassou и некоторые другие) живет и размножается в морской воде, солености не ниже нормальной океанической.
По отношению к температуре каждое из семейств отряда имеет свои существенные особенности. Наиболее разнообразно в этом отношении по своему составу семейство Gadidae. В основном это семейство является довольно холодолюбивым, ряд видов его живет и, что самое существенное, размножается при температурах, близких к 0° и даже ниже, таковы: Arctogadus borisovi, Boreogadus saida, Boreogadus agilis, Eleginus navaga, в меньшей степени Eleginus gracilis и некоторые другие. С другой стороны, ряд видов требует для своего существования значительно более высоких температур. Наиболее теплолюбивы из Gadidae: Trisopterus luscus, Trisopterus minutus minutus, Trisopterus minutus capelanus, Molva dipterygia elongata и некоторые другие. Представители семейства Bregmacerotidae, в противоположность семейству Gadidae, более теплолюбивы, так как живут в тропических частях всех океанов, где встречаются как на глубинах, так и в поверхностных слоях воды. Виды семейства Moridae являются рыбами довольно холодолюбивыми и, приспособившись к обитанию в холодных придонных глубинных слоях воды, живут на больших глубинах у границы или за пределами континентальной ступени.
По вертикальному распределению каждое из семейств Gadiformes имеет точно так же свои особенности. Семейство Gadidae наиболее разнообразно по своему составу и в этом отношении. Почти все виды этого семейства в той или иной мере связаны с дном, за исключением Micromesistius poutassou, отчасти Merluccius и некоторых других, живущих над большими глубинами и ведущих пелагический образ жизни на некоторой, иногда значительной глубине от поверхности или в самых поверхностных слоях. Одни из видов тресковых живут у самого берега или на незначительной глубине, как, например, Raniceps raninus, Eleginus navaga, Eleginus gracilis, Microgadus tomcod, Microgadus proximus и, возможно, Arctogadus borisovi, или среди пловучих льдов, как Boreogadus saida. Ряд других видов семейства Gadidae, как, например, Gadus morhua morhua, Melanogrammus aeglefinus и многие другие, живут на значительных глубинах — до 200 м и выше. Наконец, некоторые из видов семейства живут на больших глубинах, некоторые до 1000 м. К числу наиболее глубоководных видов относятся Brosme brosme, Molva dipterygia dipterygia, Molva dipterygia elongata, Gadiculus argenteus argenteus, Gadiculus argenteus thori и др., причем некоторые из них на больших глубинах держатся, возможно, не у самого дна, а в промежуточных слоях. Все представители семейства Moridae, как правило, живут на больших глубинах в несколько сот метров и даже свыше 1000 м и в прибрежной области обычно не встречаются. Виды семейства Bregmacerotidae ведут пелагический образ жизни. Экология и биология представителей семейств Moridae и Bregmacerotidae вообще изучены слабо, и сведения относительно них очень ограничены.
Почти все Gadiformes, насколько это известно, совершают миграции. Совершенно не изучены в этом отношении лишь семейства Moridae и Bregmacerotidae, что же касается семейства Gadidae, то миграции представителей его, точно так же как и другие вопросы биологии и экологии их, изучены значительно полнее. Некоторые из тресковых совершают небольшие по протяжению миграции, в зависимости от изменения температуры от берегов на несколько большие глубины и обратно или в пределах сравнительно ограниченной части своего ареала. К числу рыб, живущих более или менее оседло и совершающих лишь местные ограниченные миграции, относятся виды рода Gaidropsarus, Enchelyopus cimbrius и некоторые другие. Несколько более подвижными, но совершающими миграции, повидимому, только в ограниченном участке своего ареала, являются Eleginus navaga, Eleginus gracilis, Microgadus tomcod и, вероятно, Microgadus proximus. Boreagadus saida активно больших миграций не совершает, но вместе с дрейфующими льдами переносится, вероятно, на значительные расстояния. Особенно же отдаленные миграции совершают океаническая форма трески Gadus morhua morhua и пикша — Melanogrammus aeglefinus, которые от берегов Норвегии уходят летом почти к берегам Новой Земли, совершая в течение короткого времени путь протяжением в 1500—2000 км. Такие же отдаленные передвижения совершает треска и между Гренландией и Исландией.
Миграции тресковых, как и всех рыб, теснейшим образом связаны о питанием и размножением и в значительной степени зависят от морских течений и сезонных изменений температуры. Треска, миграции которой наиболее изучены, весной, отощав после икрометания, с нерестилищ у Лофотенских островов направляется вдоль струй Североатлантического течения на север в погоне за пищей или в поисках ее. Это весеннее движение голодной трески на север теснейшим образом связано с повышением температуры воды. С наступлением осени и с понижением температуры воды, лофотенская треска постепенно покидает восточные районы Баренцева моря и зимой лишь частично удерживается в западных его районах. С целью питания и в самой тесной связи с изменениями температуры совершает свои почти такие же отдаленные миграции и пикша (Melanogrammus aeglefinus), точно так же как и сайда (Pollachius virens), Micromesistius poutassou и некоторые другие, совершающие миграции несколько меньшего протяжения и не такие массовые, как предыдущие два вида. Под влиянием сезонных изменений температуры и для целей питания совершают свои передвижения и те тресковые, которые мигрируют лишь с глубин к берегам и обратно.
Каждый из видов семейства вне периода икрометания живет на более обширном ареале и при значительно более широких пределах колебаний температуры, солености и глубин, чем в этот период. Во время икрометания половозрелые тресковые собираются на довольно постоянных и известных почти для каждого вида местах, где каждый вид мечет икру при определенных для него, более узких, чем в обычное время, пределах колебаний температуры, солености и глубины. Экологические особенности каждого вида тресковых в период нереста более характерны, чем экология их вне этого периода, и на этом основании некоторыми авторами (Hjort, 1909 : 58; Damas, 1909 : 223; J. Schmidt, 1909 : 178) предложено деление тресковых на несколько биологических групп.
По условиям, местам и времени нереста существенно различаются иногда близкие между собой виды, чему может служить наиболее ярким примером род Molva. Так, Molva molva мечет икру при температуре 6—10°С и солености 35.2‰ на глубине в среднем около 200 м, Molva dipterygia dipterygia — при температуре 5—8°С и солености 35.2‰ на глубине в среднем около 1000 м, Molva dipterygia elongata — при температуре 8—9°С и солености 35.5‰ на глубине в среднем около 1000 м. В связи с этими особенностями размножения каждого вида, нерестилища их находятся в разных местах: у Molva molva ближе к берегам, у обоих подвидов Molva dipterygia на глубинах, причем у Molva dipterygia dipterygia нерестилища расположены к северу от Ирландии, а у Molva dipterygia elongata — к югу. Такие «викарирующие» виды и формы среди тресковых имеются не только по местам, но и по времени нереста.
Главное время нереста тресковых — вторая половина зимы и начало весны, у немногих нерест происходит летом, у многих же видов в небольших размерах может происходить почти круглый год. Время нереста у одного и того же вида в значительной степени зависит от географической широты — в южных местах ареала нерест наступает ранее, чем в более северных.
Плодовитость тресковых крайне велика: крупная Molva molva имеет до 60 млн. икринок (Fulton, 1891: 243), Gadus morhua имеет 4—6, иногда до 9 млн. икринок, причем икра у нее созревает не вся сразу и выметывается, повидимому, не одновременно, так как в зрелом яичнике различают несколько стадий развития икринок. Количество икринок зависит от размеров рыбы: крупные виды тресковых имеют большее количество икринок, мелкие — значительно меньшее; навага, например, имеет лишь от 6 до 90 тыс. икринок.
Икра, личинки и мальки почти всех тресковых ведут пелагический или у некоторых (Micromesistius, Gadiculus) батипелагический образ жизни, причем мальки многих видов (Gadus morhua, Melanogrammus aeglefinus, Eleginus navaga, Eleginus gracilis, Odontogadus merlangus и некоторых других) скрываются под колоколом медуз (Cyanea), а мальки, уже ведя придонный образ жизни (Urophycis chuss), — между створками моллюсков (Pecten magellanicus). Некоторые виды тресковых (например Eleginus navaga) имеют икру демерсальную, у Lota lota икра откладывается на дно или находится во взвешенном состоянии у самого дна. Инкубационный период продолжается от недели до месяца, у арктических видов — значительно долее, чем у бореальных.
При развитии икринки в малька различают (Расс, 1933 : 12) несколько стадий: 1) икринки, 2) личинки, характеризующейся наличием желточного мешка, 3) неоформившегося малька, длящейся от момента полной резорбции желточного мешка до появления лучей в непарных плавниках, 4) оформившегося малька, имеющего лучи в непарных плавниках, но без чешуйчатого покрова и 5) молоди или сеголетка — вполне сформировавшейся рыбки с чешуйчатым покровом.
В течение пелагического периода жизни икра, личинки и мальки течениями разносятся от мест нереста иногда на далекие расстояния. У трески, мечущей икру у Лофотенских островов, мальки к концу пелагической стадии переносятся Североатлантическим течением от мест нереста на сотни километров вдоль и от берегов. Икра и мальки трески, мечущей икру у южных и западных берегов Исландии, циркулирующим здесь круговым течением уносятся к северным берегам острова.
Таким образом, еще на стадии икринки, личинки и малька происходит расселение тресковых от мест нереста. Расселение тресковых в пелагической стадии их развития не выходит за пределы течений, циркулирующих в местах нереста. Активные миграции взрослых значительно расширяют пределы распространения каждого вида. Однако размеры ареала каждого вида тресковых обусловливаются не только миграциями и экологией взрослых, но в значительной степени и экологией размножения, происходящего, как указывалось выше, при значительно более узких пределах колебаний основных регулирующих его факторов, чем при которых может жить данный вид вне периода размножения. Во взрослом состоянии многие виды тресковых могут найти подходящие условия для своего существования на очень далеком расстоянии от мест нереста, но условий для размножения здесь не встретить, и они бывают принуждены возвращаться для этого на значительно меньшие по своим размерам, сравнительно с общим ареалом, места и после икрометания опять отправляться в отдаленные миграции. Примером этому может служить Micromesistius poatassou, во взрослом состоянии проникающая в западную часть Баренцева моря, но для размножения принужденная возвращаться на сравнительно небольшую площадь, тянущуюся узкой полосой вдоль 1000-метровой изобаты от Исландии к Бискайскому заливу. Из этого же примера видно, что расширение ареала вида от его центра, за который следует считать места нереста, происходит вдоль течений. Расширение ареала при изменении внешних условий обычно сопровождается расширением в этом же направлении и нерестилищ. Треска, которая во второй половине прошлого столетия у берегов Гренландии в промысловом количестве не встречалась, в последние годы, в связи с общим потеплением климата и смещением к северу зон жизни, стала ловиться в большом количестве и держится здесь круглый год. В настоящее время установлено, что у западных берегов Гренландии происходит и нерест тресковых. Сайда (Pollachius virens), которая в течение долгого времени не встречалась у берегов Гренландии, появившись в самые последние годы, повидимому, начала здесь и размножаться.
По достижении определенных для каждого вида размеров, колеблющихся в видовом отношении от нескольких до 15 см, мальки или сеголетки постепенно опускаются на дно и, передвигаясь уже активно, приближаются к берегам. Продолжительность пелагического периода жизни молоди для каждого вида различна и колеблется от нескольких недель до нескольких месяцев; у трески, например, он длится в среднем около трех месяцев, а в северных широтах дольше. Наиболее продолжителен этот период у глубоководных видов, хотя это правило и не является общим. Пелагический период молоди у одного и того же вида более продолжителен в северных районах размножения, чем в южных.
Первые годы жизни молодь почти всех видов тресковых проводит у берегов на небольших глубинах. Глубина и удаленность мест обитания молоди от берегов для каждого вида различна и почти не стоит в какой либо связи с глубиной мест нереста. В течение нескольких первых лет придонной жизни молодь совершает лишь ограниченные миграции под влиянием сезонных изменений температуры и в поисках пищи. По мере подрастания молоди миграции удлиняются, и ко времени наступления половой зрелости протяженность их принимает нормальные для данного вида размеры.
Время наступления половой зрелости, точно так же как и рост, у разных видов тресковых неодинаково. Наиболее быстро достигают половой зрелости мелкие виды — Eleginus navaga, Boreogadus saida, которые начинают метать икру по достижении 2—4 лет. Крупные виды тресковых — Pollachius virens, Melanogrammus aeglefinus, Gadus morhua — растут значительно быстрее, но половой зрелости достигают в более позднем возрасте. Pollachius virens начинает впервые метать икру в возрасте 5—6 лет, Melanogrammus aeglefinus — в возрасте 4—5 лет, различные расы Gadus morhua — в возрасте от 4 до 15 лет.
Молодь тресковых питается мелкими беспозвоночными животными. Пища взрослых тресковых крайне разнообразна. Наиболее мелкие виды (Eleginus navaga, Boreogadus saida) питаются мелкими планктонными и бентоническими ракообразными, реже молодью рыб. Крупные виды тресковых (Gadus morhua, Melanogrammus aeglefinus, Pollachius virens) питаются иглокожими, моллюсками, крупными ракообразными и рыбой. Основной пищей некоторых видов (Gadus morhua, Pollachius virens и др.) является рыба.
ХОЗЯЙСТВЕННОЕ ЗНАЧЕНИЕ
(Приводимые здесь цифровые данные взяты из Bull. Statistique des Peches Marit., XXII (1933), 1935 — и XXIV (1934) 1936, Pacific Fisherman (1936) и Canadian Fisheries Manual 1931. Данные об уловах СССР приведены по данным работ Н. А. Маслова (1939, 1944), которые не могли быть использованы здесь полнее по причинам, указанным в предисловии. Уловы минтая указаны по данным Е. К. Суворова.)
Из всех семейств отряда хозяйственное значение имеют представители лишь семейства тресковых; представители остальных семейств для промысла значения не имеют. Тресковые по их роли в мировом промысле стоят по улову на первом месте после сельдевых. Годовой улов всех видов тресковых за последние годы достигает в среднем свыше 17 500 тыс. ц. В эту цифру не входят уловы тресковых в южном полушарии и ряд видов, ловимых в небольших количествах и не учитываемых статистикой. Нашим траловым промыслом в 1934—1938 гг. вылавливалось от 772 до 2132 тыс. ц.
Наибольшее значение из всех видов тресковых имеет треска, уловы которой в Атлантическом океане у берегов Европы, Гренландии и Ньюфаундленда в последние годы превышают 10 млн. ц (около 10.7 млн. ц в 1933 г. и около 10.4 млн. ц в 1934 г. без уловов СССР). Кроме того, в Тихом океане ежегодно в последние годы вылавливалось 1.5—2 млн. ц трески. Таким образом, треска составляет в среднем около 60—70% общего улова тресковых.
На втором месте по улову стоит пикша, которой ежегодно добывается около 2 млн. ц (около 2.13 млн. ц в 1933 г. и около 1.8 млн. ц в 1934 г.). Весьма существенное значение имеет сайда, годовой улов которой превышает 1 млн. ц (около 1.11 млн. ц в 1933 г. и около 1.2 млн. ц в 1934 г.).
Исключительно тихоокеанский вид — минтай (Theragra chalcogramma), запасы которого используются недостаточно, вылавливается все же в довольно значительных количествах: в Японии его добывали свыше 900 тыс. ц.
Довольно значительную роль в улове тресковых играет мерланг (Odontogadus merlangus merlangus); его ежегодно вылавливается около 700 тыс. ц (около 730 тыс. ц в 1933 г. и свыше 715 тыс. ц в 1934 г.). Наконец, Merluccius merluccius и Molva molva, и в играющие, подобно мерлангу, никакой роли в нашем промысле, в улове западноевропейских стран имеют довольно существенное значение: Merluccius merluccius вылавливается ежегодно свыше 400 тыс. ц (свыше 445 тыс. ц в 1933 г. и около 407 тыс. ц в 1934 г.), Molva molva — свыше 300 тыс. ц (свыше 300 тыс. ц в 1933 г. и около 350 тыс. ц в 1934 г.).
Остальные виды тресковых существенного промыслового значения не имеют и статистикой мирового промысла не учитываются.
Промысел тресковых играл существенную роль в экономике стран северо-западной Европы уже на самых ранних этапах истории. Известно, что треска являлась объектом промысла даже первобытного человека. Вначале промысел производился вблизи берегов в местах концентрации трески в определенное время года. Траловый промысел возник в западной Европе в конце XIX века и уже в начале настоящего столетия был сильно развит. Русский прибрежный промысел трески в Баренцевом море возник в XV веке. Единственным орудием лова трескового прибрежного промысла почти до последнего времени являлся ярус. Поддев, применявшийся на западном Мурмане, на восточном был почти не известен. Тресковые сети стали у нас употребляться лишь после Октябрьской революции. Прибрежный промысел производится весной и летом, в период подхода трески ближе к берегам. Возможность тралового лова трески и пикши в наших водах была установлена еще исследованиями Мурманской научно-промысловой экспедиции, однако лов этот русскими рыбопромышленниками не применялся. Развит был исключительно кустарный, прибрежный наживочный лов трески. Результаты исследований Мурманской научно-промысловой экспедиции были использованы иностранцами, в первую очередь англичанами и немцами (Книпович, 1914 : 35; Маслов, 1944 : 18,19). Русского тралового промысла до Октябрьской революции в Баренцовом море не было, если не считать несколько отдельных опытов его. Советский траловый промысел возник по окончании гражданской войны, и в 1919—1924 гг. число тральщиков достигает 11, ловящих преимущественно на Канинской банке и на банках вблизи берегов Мурмана. В 1925 г., с перенесением траловой базы из Архангельска в Мурманск, советские тральщики ловят уже круглый год, количество их увеличилось до 17, а к 1933 г. до 60, и район промысла распространился на все промысловые районы Баренцова моря. Треска является основным объектом советского тралового промысла (46—78% тралового улова в 1922—1933 гг.). Английские и немецкие тральщики в Баренцовом море в этот период трески ловили в несколько меньшем количестве, чем пикши. С 1925 г. уловы советских тральщиков превышают уловы английских тральщиков в Баренцовом море, исключая уловы последних на Медвежинско-Шпицбергенской банке; с 1937 г. уловы советских тральщиков превышают английские уловы в Баренцовом море вместе с уловами и на этой банке. Уловы немецких тральщиков в Баренцовом море вместе с уловами на Медвежинско-Шпицбергенской банке становятся меньше советских, начиная с 1932 г. В дальневосточных морях наш траловый лов начался в 1929 г. Траловый флот СССР в период времени с 1934 по 1938 г. вылавливал в Баренцовом море от 772 (1934 г.) до 2132 (1937 г.) тыс. ц. Главная масса этого улова (от 87.2 до 95.0%) приходится на треску и пикшу. В нашем промысле, как и в мировом, на первом месте по улову стоит треска, которая в Баренцовом море с 1930 по 1938 г. составляла от 47. 5 до 85.1% общего улова тралового флота. Общий улов трески в период времени с 1934 по 1938 г. колебался от 558 (1934 г.) до 1750 (1937 г.) тыс. ц. На втором месте в наших уловах тресковых рыб стоит пикша, которая за период времени с 1930 по 1938 г. составляла от 6.3 до 37% общего улова тралового промысла. Общий улов пикши за время с 1934 по 1938 г. составлял от 84 (1935 г.) до 770 (1938 г.) тыс. ц (Маслов, 1939). В небольшом количестве вылавливается в наших водах сайда. Минтай, вылавливаемый в довольно большом количестве в Японии, нашим промыслом почти не используется:
Несколько большее промысловое значение имеют оба вида наваги. Уловы наваги с 1936 по 1941 г. колебались от 11.6 (1920 г.) до 24.0 (1930 г.) тыс. ц. Дальневосточной наваги (вахни) вылавливалось свыше 16 тыс. ц. Уловы налима за эти же годы колебались от 16.7 (1941 г.) до 18.76 (1937 г.) тыс. ц.
К тресковым, почти совершенно не используемым промыслом, но встречающимся в значительных количествах, относится сайка (Boreogadus saida), играющая существенную роль в питании морских млекопитающих.
Из побочных продуктов трескового промысла следует в первую очередь указать на вытапливаемый из печени жир, имеющий благодаря большому содержанию в нем витаминов широкое применение в медицине. Тресковые головы идут на изготовление рыбной муки. С крупных рыб снимается и выделывается кожа.
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Отряд GADIFORMES (ANACANTINI ex parte)
Nervi olfactorii короткие, lobi olfactorii расположены около носовых капсул (за исключением Raniceps, у которого nervi olfactorii несколько удлинены и lobi olfactorii вследствие этого расположены между носовыми капсулами и передним мозгом), соединяясь с передним мозгом при помощи длинных tractus olfactorii. Nervi и tractus olfactorii расположены в особом канале — продолжении полости черепа над septum interorbitale и отделены, таким образом, от зрительных нервов. Орбито-ростральная часть черепа длиннее или равна (у некоторых Moridae) задней. Basisphenoideum и orbitosphenoideum отсутствуют. Миодома нет. Opisthoticum исключительно сильно развито, пронизано отверстием для n. glossopharyngeus, задним концом частично окаймляет отверстие n. vagus, снизу соприкасается с basioccipitale, отделяя prooticum от occipitale laterale. Supraoccipitale хорошо развито, с продольным гребнем на всем протяжении или лишь над задней частью, соприкасается с frontalia, разделяя правое и левое parietalia. Плечевой пояс прикреплен к черепу, mesocoracoideum нет, лопаточное отверстие между scapula и coracoideum, грудных radialia 4—13. Ossa pelvis позади ключичного симфизиса, свободно прикреплены к нему связкой. Позвонки многочисленные, первые 2 позвонка без парапофизов, ребер и epipleuralia, 2—4 следующих тоже без парапофизов, но с сидячими ребрами и с epipleuralia; все остальные туловищные позвонки с хорошо развитыми парапофизами, к которым прикреплены ребра и epipleuralia, за исключением Merluccius, у которого первые 5—6 позвонков устроены как у всех, следующий один с зачаточными парапофизами, ребрами и epipleuralia и все остальные туловищные позвонки с сильно расширенными парапофизами, но без ребер и с epipleuralia только на 3—4 первых позвонках. Остистый отросток 1-го позвонка плотно прикреплен к гребню supraoccipitale. 4 жабры, позади четвертой щель, псевдобранхий нет, жаберные перепонки свободны или узко прикреплены к межжаберному промежутку; 6—8 лучей жаберной перепонки. Все плавники без колючих лучей, 1—3 спинных, 1—2 анальных плавника, хвостовой плавник отделен от спинного и анального плавников, реже частично или совершенно (у Muraenolepidae) слит с ними, дифицеркальный (Точнее псевдокавдальный (см. выше).), брюшные плавники впереди грудных. Тело и частично голова покрыты мелкой циклоидной чешуей. На подбородке обычно имеется усик. Плавательный пузырь не соединяется с кишечником, иногда рудиментарен. Пилорические придатки многочисленные, реже единичные или отсутствуют.
Отряд более резко очерченный после выделения семейства Macruridae, состоит из семейств Muraenolepidae, Moridae, Bregmacerotidae, Gadidae.
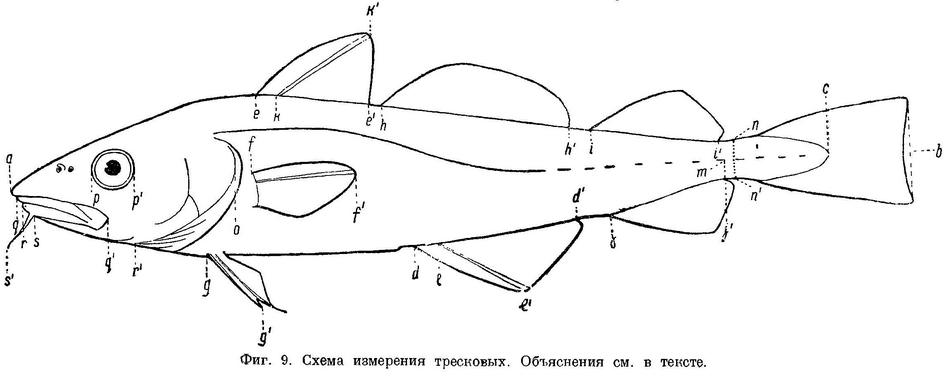
Измерения представителей отряда Gadiformes производились по общепринятой схеме, предложенной В. С. Михиным для трески, причем брались не все измерения, а лишь следующие (фиг. 9): а—b — длина всего тела; а—с — длина тела до конца чешуйчатого покрова; а—d — антеанальное расстояние; а—е — антедорсальное расстояние ; f—f' — длина грудного плавника; g—g' — длина брюшного плавника; е—е' — длина основания 1-го спинного плавника; h—h' — длина основания 2-го спинного плавника; i—i' — длина основания 3-го спинного плавника; d—d' — длина основания 1-го анального плавника; j—j' — длина основания 2-го анального плавника; k—k' — высота 1-го спинного плавника; l—l' — высота 1-го анального плавника; m—с — длина хвостового стебля; а—о — длина головы; а—р — длина рыла; р—р' — продольный диаметр глаза; q—q' — длина верхней челюсти; r—r' — длина нижней челюсти; s—s' — длина усика. Кроме того, существенное значение имеют расстояния между первым и вторым (е'—h), вторым итретьим (h'—i) спинными плавниками и обоими анальными (d'-j).
Высота тела — важный, но крайне ненадежный у тресковых признак, зависящий в сильной мере от степени наполнения желудка — не измерялась.
Более независима от степени наполнения желудка высота тела за основанием брюшных плавников, как указывает Н. В. Европейцева. Почти в равной мере зависят от степени наполнения желудка и зрелости половых продуктов расстояния от грудного и брюшного плавников до анального. Эти измерения здесь точно так же исключены. У видов с менее сложно устроенными плавниками измерения несколько упрощаются.
Ввиду того, что часто происходят недоразумения вследствие различия в измерениях, здесь необходимо дать пояснения о некоторых из них. Длина тела измерялась от вершины рыла (если нижняя челюсть вдается вперед, то от вершины ее) до линии, соединяющей концы лопастей хвостового плавника в расправленном состоянии. Эта длина считается основной и, если говорится о длине тела, то понимается именно эта длина. Грудной плавник измерялся от основания 1-го луча до вершины плавника. За длину брюшного плавника принято расстояние от основания 1-го луча до конца его. В некоторых случаях важно измерять, кроме того, длину брюшного плавника до конца наиболее удлиненного (2-го) луча, но за длину брюшного плавника всегда считается длина до конца 1-го луча, и в тех случаях, когда особо не оговорено, именно эта длина и подразумевается. Высота 1-го спинного и 1-го анального плавников измерялась по наибольшему лучу их, за исключением тех случаев, когда этот луч в спинном плавнике (у некоторых видов Phycis и Urophycis) сильно вытянут в виде нити. В этих случаях за высоту плавника приняты размеры наибольшего, но не вытянутого в виде нити луча, а удлиненный луч измерялся особо. За длину хвостового стебля принято расстояние от заднего конца последнего спинного или анального плавника до конца чешуйчатого покрова (конец чешуйчатого покрова у тресковых виден очень ясно). За высоту хвостового стебля принята наименьшая высота тела или (у Lotinae) высота его у конца последнего спинного и анального плавников. Длина головы измерялась от вершины рыла (если нижняя челюсть выдается вперед, то от вершин ее) до шипообразного конца operculum. Перепонка, окаймляющая жаберную крышку, в расчет не принималась. За ширину межглазничного промежутка принята ширина черепа в межглазничной области, причем кожные покровы не измерялись. Н. В. Европейцева рекомендует измерять, кроме того, длину хвостовой части тела от вертикали заднего конца 1-го анального плавника до конца чешуйчатого покрова.
Все измерения частей тела выражены в процентах к длине тела (до конца лопастей хвостового плавника), а частей головы — в процентах к длине головы. Более правильно и точно было бы выражать измерения в процентах к длине тела до конца чешуйчатого покрова, как это делает для налима М. И. Маркун, но так как всеми принято относить измерения ко всей длине тела, то нарушать это общепринятое правило не следует.
При просчете количества лучей во всех непарных плавниках, за исключением 1-го спинного, перепонку у основания начала и конца каждого из плавников необходимо подрезать, так как иначе несколько лучей, скрытых в перепонке и кожных покровах, остаются непросчитанными. Жаберные тычинки просчитывались в наружном ряду 1-й жаберной дуги, причем тычинки просчитывались все, включая и зачаточные. Позвонки считались все, включая и последний, к которому непосредственно присоединяются средние лучи хвостового плавника.
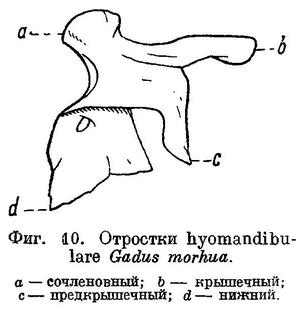
Объяснение названий частей черепа, упоминаемых при его описании, см. на фиг. 5 вводной части. Отростки hyoinandibulare названы здесь следующим образом (фиг. 10): сочленовный — отросток, которым эта кость причленяется к черепу; крышечный — отросток, к которому причленяется operculum; предкрышечный отросток, к которому присоединяется praeoperculum, и нижний — отросток, к которому присоединяются снизу symplecticum и stylohyale. Поры, ведущие в каналы системы боковой линии, расположенные на голове и имеющие важное систематическое значение, в диагнозах перечислены для can. praeoperculo-mandibularis, can. infraorbitalis, commissura и can. supraorbitalis. Количество пор в portio supratemporalis и portio cephalica lineae lateralis постоянно, и поэтому эти поры не перечисляются.
В настоящее время, при наличии большого числа работ с детальным описанием черепа некоторых видов тресковых, нет надобности останавливаться на общем описании черепа Gadidae. Поэтому здесь обращено внимание, главным образом, лишь на те особенности в строении черепа, которые имеют значение для системы или различения родов и видов тресковых. С таким же расчетом выбраны и немногие, кратко описываемые здесь кости висцерального скелета, жаберной крышки и поясов конечностей, рисунки которых могут оказать помощь, кроме того, и при изучении питания рыб. Следует, однако, иметь в виду, что почти все эти кости висцерального скелета вариируют в довольно широких пределах. Рисунки позвоночников могут оказать существенную помощь при изучении ископаемых рыб, причем особое значение имеют остистые и поперечные отростки и гемальные дуги, по размерам и форме которых значительно отличаются виды и роды тресковых. Ребра и epipleuralia, слабо прикрепленные к позвонкам и при удалении мышц обычно отваливающиеся, на этих рисунках не изображены.
Остеология некоторых видов (Melanogrammus aeglefinus, Eleginus navaga, Eleginus gracilis, Boreogadus saida и, в особенности, Gadus morhua morhua) изучена по нескольким десяткам скелетов. Ряд видов (Gaidropsarus mediterraneus, Raniceps raninus, Phycis phycis, Urophycis regius, Urophycis brasiliensis, Lota lota lota, Molva molva, Molva dipterygia elongata, Merluccius merluccius, Trisopterus minutus capelanus, Odontogadus merlangus euxinus, Gadus morhua morhua n. hiemalis, Gadus morhua maris-albi, Gadus morhua macrocephalus) изучен по 2, 3, 4 и более скелетам. Остальные, более редко встречающиеся виды, изучены по одному скелету.
Количество исследованных экземпляров указывается в скобках после описания каждого вида; таких указаний нет лишь в тех случаях, если исследовано более 10 рыб.

ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА СЕМЕЙСТВ
1 (2). Хвостовой плавник слит со спинным и анальным. Грудных radialia не менее 10. (Подотряд Muraenolepidoidei). 1. Muraenolepidae
2 (1). Хвостовой плавник отделен от спинного и анального плавников, реже частично слит. Грудных radialia 4—6. (Подотряд Gadoidei).
3 (4). Плавательный пузырь через фонтанелли в occipitalia lateralia сзади черепа соприкасается с ушными капсулами. Обнаружить это очень легко, приподняв жаберную крышку и сделав над жабрами продольный надрез вдоль основания черепа. При этом у Moridae виден рожкообразный отросток плавательного пузыря, а если приподнять его, то и фонтанелль. Продолжив надрез вперед под глазом, можно далее установить, костная или перепончатая нижняя часть канала над межглазничной перегородкой. Neurocranium спереди почти закрыт, канал обонятельных нервов и частично межглазничная перегородка костные. Головка vomer часто без зубов. Глубоководные рыбы. 2. Moridae
4 (3). Фонтанеллей езади черепа в occipitalia lateralia нет, плавательный пузырь не соприкасается с ушными капсулами. Neurocranium широко открыт спереди, межглазничная перегородка и канал обонятельных нервов снизу перепончатые. Головка vomer всегда с зубами. Рыбы обычно умеренных глубин.
5 (6). Первый спинной плавник на затылке и состоит лишь из одного тонкого длинного луча. Орбито-ростральная часть черепа равна задней. Канал обонятельных нервов широкий, по ширине почти равен neurocranium. 3. Bregmacerotidae
6 (5). Первый спинной плавник позади головы, обычно нормального строения, реже состоит из одного луча. Орбито-ростральная часть черепа значительно длиннее задней. Канал обонятельных нервов узкий. 4. Gadidae
Рыбы пресных вод СССР и сопредельных стран. Л.С.Берг 1948 г.
VI. Отряд GADIFORMES. — Трескообразные
Anacanthini Regan, Ann. Mag. Nat. Hist. (7), XI, 1903, p. 459 (частью). — Gadiformes Берг, Система рыб, 1940, стр. 283; Световидов, Трескообразные, 1948, стр. 62.
Закрытопузырные. Колючек в плавниках нет. Брюшные плавники впереди грудных. Чешуя циклоидная. В костях нет костных клеток. Обонятельные нервы расположены не в глазничной полости по соседству с межглазничной перегородкой (как обычно у большинства Teleostei), а помещаются в канале, который лежит над межглазничной перегородкой и сообщается с черепной полостью. Обонятельные лопасти в непосредственном соседстве с носовыми капсулами; таким образом мозг продолжается далеко вперед. Opisthoticum очень велико. Нет миодома, орбитосфеноида и базисфеноида; нет мезокоракоида. Первые позвонки без парапофизов. Межмышечных косточек нет (верхние ребра имеются). Главным образом морские рыбы. 4 семейства. Из них пресной воде свойствен налим, и иногда попадаются другие виды.