(Gnathostomata Zittel, 1879) Инфратип Гнатостоматы, Челюстноротые, Infraphylum Gnathostomata Zittel, 1879
Материал подготовил Фактистов И. Е., 2020 г. Сайт www.fishbiosystem.ru
Гнатостоматы, Челюстноротые (Gnathostomata) — инфратип (суперкласс, группа) позвоночных животных (Vertebrata). Включает около 60000 видов (99% всех позвоночных). Подразделяется на надклассы рыб и четвероногих.
Название инфратипа Gnathostomata происходит от древнегреческих слов γναθος (gnathos) — «челюсть» и στομα (stoma) — «рот», что соответствует русскому переводу — челюстноротые.
Как и следует из названия группы, её представители характеризуются наличием противопоставленных челюстей. Кроме этого, челюстноротые отличаются от других хордовых наличием зубов, парных конечностей, третьим (горизонтальным) полукружным каналом внутреннего уха. На клеточном уровне следует отметить наличие у челюстноротых миелинового покрытия нейронов и адаптивной иммунной системы, которая применяет рекомбинацию V(D)J-цепочек антител для распознавания антигенов.
В классической систематике челюстноротые делятся на два надкласса: рыб (Pisces) и четвероногих (Tetrapoda). Поскольку четвероногие произошли от рыб, последние являются парафилетической группой. Поэтому рыбы признаются естественным объединением и допустимым таксоном лишь в рамках эволюционной таксономии, но не кладистики.
Отдельные подгруппы рыб также получают разную трактовку со стороны эволюционной таксономии и кладистики. Именно, костные рыбы (Osteichthyes) и лопастепёрые рыбы (Sarcopterygii) в эволюционной таксономии рассматриваются как парафилетические таксоны (не включающие происшедших от них четвероногих). В кладистике названия этих двух таксонов обозначают уже клады, получаемые добавлением к ним всех членов надкласса Tetrapoda.
Переходя к общей характеристике челюстноротых, заметим, что их название чётко соответствует важному эволюционному приобретению этих животных по сравнению с бесчелюстными: их рот вооружён подвижными челюстями. Во внутреннем ухе имеется уже три полукружных канала, а не два, как у бесчелюстных. Челюстноротые способны к энергичным передвижениям, к активному захватыванию найденной пищи.
Считается, что челюсти произошли от передних опорных дуг жабр, которые приобрели новую роль, будучи модифицированными для перекачивания воды через жабры, более эффективно открывая и закрывая рот — это как механизм насоса. Рот мог тогда увеличиваться и расширяться, давая возможность захватывать более крупную добычу. Этот механизм закрывания и открывания со временем становился все сильнее и сильнее, превращая дуги жабр в настоящие челюсти.
Более новые исследования показывают, что ветвь пластинокожих рыб или плакодерм (Placodermi), скорее всего, была предком современных челюстноротых. У окаменелости плакодермы по имени Entelognathus возрастом 419 миллионов лет был костный скелет и анатомические детали, связанные с хрящевой и костной рыбой, демонстрируя, что отсутствие костного скелета у Chondrichthyes является производной чертой. Окаменелости примитивных костистых рыб, таких как Guiyu oneiros и Psarolepis, которые жили одновременно с Entelognathus и имели тазовые пояса больше схожи с плакодермами, чем с другими костными рыбами, показывают, что они были родственниками, а не прямыми предками сохранившихся челюстноротых. Находки окаменелостей молодых плакодерм, у которых были настоящие зубы, которые росли на поверхности челюстной кости и не имели корней, что делало невозможным их замену или повторное отрастание по мере того, как они ломались или изнашивались по мере взросления, доказывает, что общий предок всех Гнатостомат имел зубы, и их происхождение было связано с эволюцией челюстей или вскоре после неё.
Поздние ордовикские окаменелости того, что было идентифицировано как чешуйки акантоидов или «акулоподобных рыб», могут означать первое появление Gnathostomata в летописи окаменелостей. Однозначно неоспоримо, что окаменелости челюстноротых, в основном примитивных acanthodians, начинают появляться в начале силура, и умножатся в начале девона.
Группа Гнатостоматы, Челюстноротые (Gnathostomata) традиционно является суперклассом, разбитым на три группы верхнего уровня: Chondrichthyes, или хрящевые рыбы; Плакодерми (Placodermi), вымершая клада бронированных рыб; и Teleostomi, включающая знакомые классы костных рыб, птиц, млекопитающих, рептилий и земноводных. Некоторые системы классификации используют термин Amphirhina. Это сестринская группа бесчелюстным (Agnatha).
Рыбы открытого океана. Н.В.Парин 1998 г.
Группа животных, попросту называемых в повседневной жизни рыбами, с точки зрения зоолога-систематика, объединяет 4 разных класса, различающиеся между собой не менее чем пресмыкающиеся, птицы и млекопитающие. Два из них — миксины (Myxini) и непарноноздревые (Monorhina) вместе с группой давно вымерших панцирных рыб относятся к особой ветви бесчелюстных подтипа позвоночных; два других класса — хрящевые рыбы (Chondrichthyes) и костные рыбы (Osteichthyes) — принадлежат к ветви челюстноротых того же подтипа.
Ихтиопланктон открытого океана включает не только ранние стадии развития океанических рыб (естественно, исключая щележаберных — акулы и скаты с самого начала жизни принадлежат к нектону), но и личинок и мальков множества шельфо-неритических видов, которые попадают в эпипелагиаль в результате пассивного выноса. В тропических водах его характеризует эфемерность существования — ведь развитие икры продолжается, как правило, не более 2 суток (правда, у макрелещуковых и летучих рыб — 2 недели, у вогмеровых рыб — до 2 месяцев), а личинки находятся в составе планктона в среднем около 8 недель. В более холодноводных районах эти сроки удлиняются, и длительность предмалькового периода жизни достигает у большинства видов 4—6 месяцев. Сильно растянуто личиночное развитие и у некоторых тепловодных форм, в частности у многих камбаловых и угреобразных рыб (личинки европейского угря, например, проводят в ихтиопланктоне до 3 лет), и это сильно способствует их дальнему разносу и широкому pacnpoстранению.
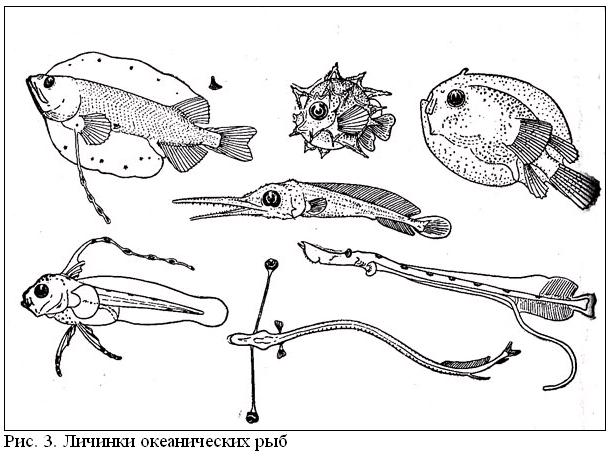
Икринки и личинки имеют приспособления к планктонному существованию, способствующие парению в толще воды: разного рода выросты и придатки на теле, удлиненные лучи плавников, а также жировые включения и газовые пузырьки (рисунок 3). Как правило, они сильно отличаются по всему своему облику от взрослых особей и в ряде групп претерпевают резкий метаморфоз (изменение строения) в ходе индивидуального развития. Поэтому установить принадлежность раннего малька, а тем более личинки (не говоря уже об икринке) к тому или иному виду, роду и даже семейству во многих случаях совсем непросто.
Форма тела нектонных рыб соответствует их активному образу жизни. Наиболее быстрым и подвижным хищникам — тунцам, акулам и др. — свойственна возникшая конвергентным путем торпедообразная форма тела. У некоторых рыб (меч-рыба, марлины) оно снабжено; спереди еще особым рострумом-обтекателем, который, как предполагает В. В. Овчинников, турбулизирует набегающий поток и, подобно выступу в подводной части носа современных быстроходных судов, сильно уменьшает лобовое сопротивление. Важную роль в обеспечении возможности быстрого движения имеют и хвостовые кили, увеличивающие поперечную жесткость хвостового стебля и выполняющие роль горизонтальных стабилизаторов. Для нектоирых рыб довольно обычно также симметрично сжатое с боков тело, в той или иной степени вытянутое в длину, или стреловидное тело с отставленными кзади спинным и анальным плавниками. В открытом океане минимальные размеры нектонных рыб могут, по-видимому, находиться в пределах от 15 до 20—30 см.
Летучие рыбы, сайра и макрелещука, а также приповерхностные светящиеся анчоусы принадлежат к числу рыб, привлекаемых источниками искусственного освещения: реагируя ночью на судовые огни, летучие рыбы, например, нередко залетают на палубу судов, а в отдельных случаях даже в открытые иллюминаторы кают. Такая особенность поведения важна как для научного изучения этих рыб, так и для их промысла. Наблюдения на световых станциях, часто включаемых в программы экспедиционных рейсов, позволяют узнать много интересного.
Тихая темная ночь... Судно медленно дрейфует, развернувшись лагом к чуть заметному ветерку... В таинственно мерцающей голубой линзе воды, границы которой постепенно переходят в кромешную черноту, появляются то зигзагом проскакивающие освещенную зону сайры, сверкающие, как рыболовные блесны, то менее скоростные, но очень юркие светящиеся анчоусы, то медленно дрейфующие по поверхности с расправленными крыльями мальки летучих рыб... Взрослые летучки подплывают к свету поодиночке или стайками, а иногда и подлетают над водой, с силой ударяясь головой о борт судна... Появление кальмаров и хищных рыб — змеиных макрелей и корифен — всегда неожиданно: их стремительные броски в световом пятне, нередко заканчивающиеся захватом добычи, напоминают выпады разящего клинка или сверкание молнии... Крупные акулы (у поверхности наиболее обычна длиннокрылая), напротив, пересекают зону света медленно и чинно...
Иной раз, впрочем, приходится часами простаивать, облокотившись о планширь и ожидая появления очередной рыбы с готовым к броску закидным сачком — сеткой на обруче, к которому прикреплен длинный линь (сказать бы попросту «веревка», но этого слова в морском лексиконе нет). Ловля таким сачком — настоящее искусство, в котором истинным асом был на «Витязе» Григорий Касьянович Фисунов - старый моряк, долго занимавший пост техника-лаборанта при ихтиологической лаборатории. Ему удавались фантастические по дальности и точности броски.
На световых станциях можно увидеть почти всех обитателей верхнего 20-метрового слоя эпипелагиали. Действительно, список океанических рыб, показывающихся более или менее регулярно у самой поверхности, можно пополнить совсем немногими видами. В их числе нельзя не упомянуть некоторых акул, рыб-лун, копьерылов и тунцов.
Атлас-определитель рыб: Книга для учащихся. Н.А.Мягков 1994 г.
ВНЕШНЕЕ СТРОЕНИЕ РЫБ
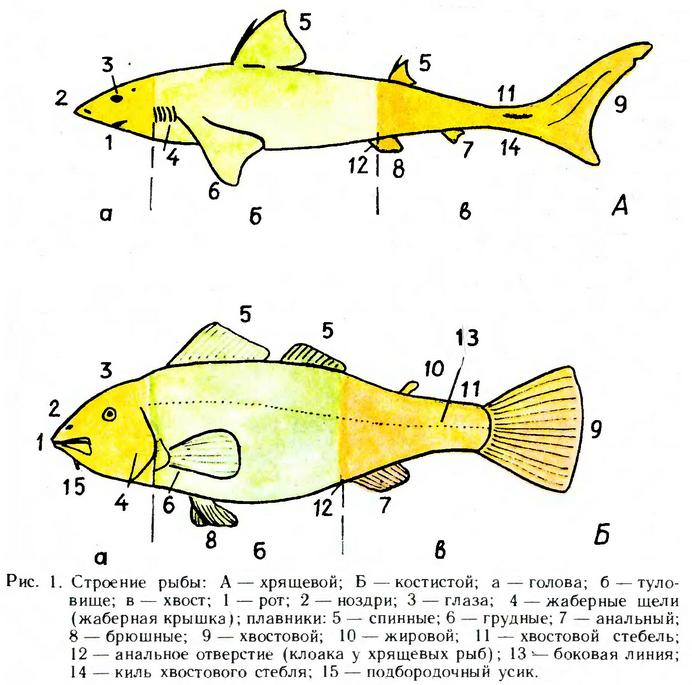
Рыбы и рыбообразные имеют тело, разделяемое на три отдела: голову, туловище и хвост. Голова заканчивается у костных рыб на уровне заднего края жаберной крышки, у круглоротых и хрящевых рыб — на уровне первого жаберного отверстия (щели). Туловище у всех рыб заканчивается на уровне анального отверстия (рис. 1), иногда его называют телом. Хвост состоит из хвостового стебля и хвостового плавника, который у некоторых видов может отсутствовать (редуцирован) или быть представленным длинной нитью (хвостовая нить).
Рыбы имеют парные (соответствующие поясам конечностей наземных позвоночных животных) и непарные плавники (круглоротые — лишь последние). К парным плавникам относят грудные и брюшные плавники, к непарным — хвостовой, спинные (один — три) и один или два анальных плавника; у ряда видов есть непарные хвостовые плавнички — над и под хвостовым стеблем (скумбрия) и жировой плавник — позади спинного (лосось). Часть брюшных плавников у самцов хрящевых рыб и анального у некоторых костных (для которых характерно внутреннее оплодотворение) в процессе эволюции видоизменились в особый совокупительный орган. У разных видов рыб плавники видоизменились в своеобразные присоски (бычки), «удочки» (удильщики) и т. д.
Плавники лучеперых рыб имеют скелет, состоящий из колючих (жестких) и ветвистых (мягких) лучей. Колючие лучи могут иметь вид мощных шипов (например, у сомовых, карповых и др.); у некоторых акул и скатов на теле и перед спинными плавниками есть мощные колючки и шипы — это модифицировавшиеся в процессе эволюции плакоидные чешуи. По наличию и характеру лучей в плавниках большинства костных рыб составляется плавниковая формула, которая широко используется при их описании и определении. В этой формуле латинскими буквами приводится сокращенное обозначение плавника: А — плавник анальный (от лат. pinna analis), Р — плавник грудной (p. pectoralis), V — плавник брюшной (p. ventralis) и D1, D2,... — плавники спинные (p. dorsalis); римскими цифрами даны числа колючих, а арабскими — мягких лучей.
Хвостовой плавник у рыб может иметь самую разнообразную форму (рис. 4): если обе его лопасти, верхняя и нижняя, равны по размерам и форме, то он называется гомоцеркальным; если одна из них больше другой — гетероцеркальным. Если длиннее верхняя лопасть, плавник эпибатный (акулы), если нижняя — гипобатный (чехонь, летучие рыбы). На хвостовом стебле у ряда рыб может быть развит костный или кожистый киль; у некоторых развиваются отдельные крупные чешуи или щитки.
Форма тела у рыб сильно различается и связана с условиями, в которых обитают особи данного вида. Рыбы, живущие в толще воды, имеют, как правило, торпедовидную или стреловидную форму, которая позволяет им развивать большие скорости со сравнительно малыми затратами энергии. Донные рыбы чаще всего имеют уплощенную (бычки, морские лисички) или даже совсем плоскую форму тела (камбалы, скаты). Виды, обитающие среди водных растений, камней и коряг у дна, обычно имеют форму тела, сильно сжатую с боков (лещи, караси). Это, очевидно, обеспечивает им лучшую маневренность. Есть и целый ряд своеобразных форм тела у рыб (рис. 2), но в наших водах подавляющее большинство видов рыб по форме тела укладывается в названные группы.
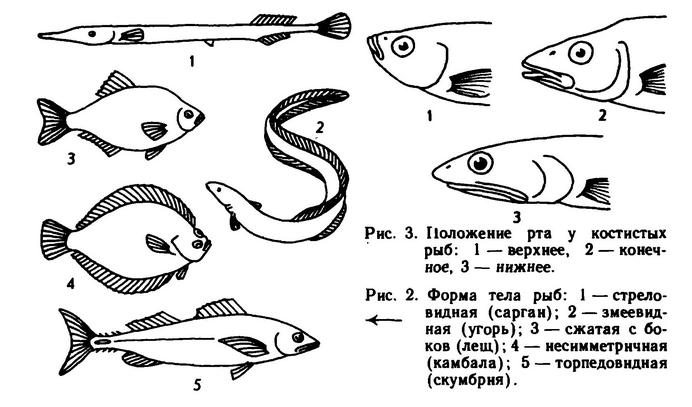
На голове рыб расположен рот, который может быть различным по размеру и положению. По последней характеристике, также связанной с экологией (типом питания), рыб разделяют на виды с нижним, верхним, конечным положениями рта; есть и промежуточные варианты. Для рыб приповерхностных вод характерно верхнее положение рта (чехони, верховки), что позволяет им подбирать добычу, упавшую на поверхность воды. Для видов-хищников и других обитателей толщи воды характерно конечное положение рта (окуни, судаки), а для обитателей придонной зоны и дна водоема — нижнее (лещи). Наиболее характерный нижний рот имеют многие акулы и скаты (рис. 3). У круглоротых, не имеющих настоящих челюстей, рот как таковой отсутствует; он заменен ротовой воронкой, вооруженной роговыми зубами.
Из органов чувств у рыб надо прежде всего назвать обоняние, представленное на голове отверстиями обонятельных капсул (есть у всех рыб). На голове расположены глаза, у некоторых пещерных и глубоководных рыб глаза могут быть частично или полностью редуцированы. Вокруг рта у многих видов рыб есть усики различной длины (от одного-двух до нескольких десятков), в большинстве своем они играют роль органа осязания. По голове проходят сейсмосенсорные каналы, входящие в систему органов чувств боковой линии рыб, продолжающейся на туловище и реже на хвостовом стебле. У многих видов боковая линия хорошо видна как одна или несколько цепочек чешуек с мелкими отверстиями. На голове хрящевых рыб хорошо видны отверстия особых органов чувств (ампул Лоренцини и пузырьков Сабо), улавливающих малейшие изменения электропроводности среды и т. д. Они также входят в систему органов чувств боковой линии. Подобные электрочувствительные органы-ампулы известны на сегодня у некоторых костных рыб, например осетровых, некоторых сомовых и большой группы тропических сильно- и слабоэлектрических рыб.
У рыб достаточно хорошо развиты и другие органы чувств, хотя обитание в водной среде и наложило на них свой отпечаток. У рыб нет внешних органов слуха (слуховых отверстий или ушных раковин), но хорошо развитое внутреннее ухо позволяет им слышать звуки. Некоторые рыбы могут и сами производить разные звуки, так что поговорка: «Нем как рыба» — мало соответствует действительности. Рыбы хорошо распознают запахи и наличие вкусовых веществ в воде; у многих видов вкусовые почки расположены не только в ротовой полости и на губах, но и на различных усиках и кожных выростах вокруг рта.
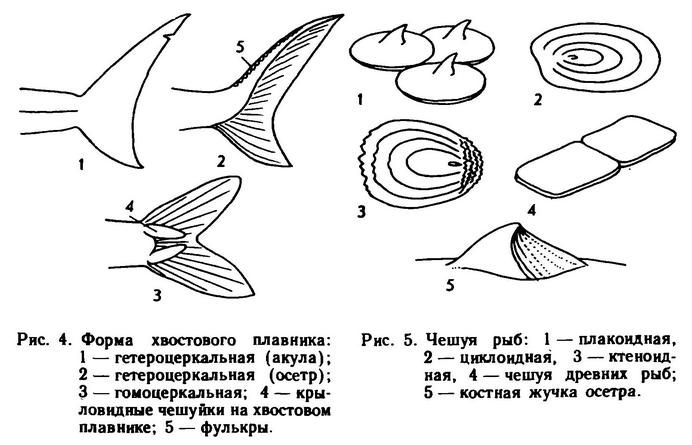
Тело рыб может быть голым, покрытым слизью или покрытым чешуей четырех типов: плакоидной (акулообразные), ганоидной (двоякодышащие тропические виды), циклоидной (сельди и др.) и ктеноидной (окуни и др.). Существуют различные модификации чешуи и защитные костные образования на теле рыб (рис. 5) — жучки осетровых, костные щитки некоторых сомовых и т. д. Чешуя на теле рыб может располагаться различным образом (сплошным покровом, участками и т. д.), а также быть различной по форме и размерам.
Окраска рыб может быть самой разнообразной. Особенно ярко раскрашены тропические морские и пресноводные рыбы.
Почти во всех случаях окраска рыб играет либо маскирующую (от хищников), либо сигнализирующую (у стайных видов), либо предупреждающую (у ядовитых видов) роль. Часто окраска рыб изменяется в зависимости от сезона, условий обитания и физиологического состояния; наиболее ярко многие виды рыб окрашены в период размножения. Существует понятие брачная окраска (брачный наряд) рыб. Все богатство окраски рыб определяется развитием и функционированием особых образований — меланофоров, лежащих в коже рыб.
В период размножения у некоторых рыб, кроме окраски тела, появляются и другие «элементы» брачного наряда; в частности, на чешуе многих карповых (например, плотвы) появляются «жемчужные» бугорки.
На жаберных крышках и других костях черепа многих костных рыб (морские окуни, скорпены, керчаки и др.) развиты мощные шипы и колючки, играющие защитную роль. Часто их форма, число и положение используются при определении видовой принадлежности рыб.
По местообитанию и отношению к солености воды всех рыб можно разделить на три группы: морские (живут при солености, близкой к океанической, равной 35 промиллям), пресноводные (не переносят осолонения) и солоноватоводные, встречающиеся как в приустьевых участках моря, так и в низовьях рек. Последние виды близки к полупроходным рыбам, нагуливающимся в солоноватоводных дельтах, губах и лиманах, а нерестящимся в реках и пойменных озерах. Ряд видов, обычно живущих в морской или пресной воде, может легко переходить в новых условиях к нетипичной для себя «воде». Так, ряд морских игл распространился в водохранилищах наших южных рек.
Отдельную группу образуют проходные рыбы, нагуливающиеся и созревающие в море, а на нерест приходящие в реки или, наоборот, совершающие нерестовые миграции из рек в моря. Чаще всего рыбы возвращаются в те водоемы, где они сами появились на свет (это явление носит название хоминга — инстинкта дома). Пока еще неизвестны механизмы, позволяющие проходным рыбам находить безошибочно свою родную речку или озеро. Рыб, идущих в реки из моря на нерест, называют анадромными, а их переход — анадромными нерестовыми миграциями. К этим рыбам относятся многие ценнейшие в промысловом отношении осетровые и лососевые рыбы. Есть у них и немногочисленные антиподы, живущие в реках, а на нерест уходящие в море. Таких рыб и их миграции называют катадромными. Среди нашей фауны катадромные путешествия совершает речной угорь, живущий и созревающий в реках и озерах, а для продолжения рода уходящий в сердце Атлантического океана, в Саргассово море.
Крайне различаются рыбы и по характеру питания. Принято выделять четыре большие группы (хотя это деление во многом условное): планктофаги, питающиеся мелкими планктонными животными—дафниями, циклопами и др. (зоопланктофаги) или водорослями (фитопланктофаги); бентофаги, поедающие организмы бентоса, обитателей дна, прежде всего различных беспозвоночных (червей, моллюсков, личинок насекомых и др.); хищники, нападающие на более мелких рыб; паразиты (в наших водах — круглоротые).
Характер питания рыб может сильно меняться с возрастом: часто молодь является планктофагом или бентофагом, а с возрастом переходит к хищничеству. Так, например, белуга после выклева из икринки питается бентосом, а затем все более и более тяготеет к хищничеству. Наши судак и окунь в самом юном возрасте — типичные планктофаги, но довольно скоро начинают нападать на молодь других рыб и даже на особей своего вида (каннибализм). Личинки миног питаются детритом и органическими веществами ила, а взрослые формы большинства из них паразитируют на рыбах.
НАДКЛАСС ЧЕЛЮСТНОРОТНЫЕ [GNATHOSTOMATA]
Позвоночные черепные животные с хрящевым или костным скелетом, двумя поясами конечностей (парные плавники), которые в той или иной степени могут быть редуцированы у некоторых видов. Характерно наличие хорошо развитых челюстей и у большинства видов — зубов на них. Надкласс объединяет ряд современных и вымерших классов водных и наземных позвоночных животных.
Редкие и исчезающие животные. Рыбы Под редакцией академика В.Е.Соколова 1994 г.
Рыбы — наиболее крупная группа позвоночных животных. Она насчитывает свыше 20 тыс. видов, т. е. около половины обитающих на земном шаре позвоночных. При этом «инвентаризация» ихтиофауны еще далеко не закончена, ежегодно публикуются десятки описаний новых видов, обнаруживаются новые роды и даже семейства.
Рыбы также и наиболее древняя группа позвоночных. Они появились на Земле свыше 400 млн. лет назад и за столь длительный отрезок эволюции освоили практически все известные типы водоемов — океаны, моря, реки, ручьи, озера, болота и др. Результатом этого явилось чрезвычайное многообразие жизненных форм рыб, их образа жизни и приспособлений. Рыбы обитают как в пресных водоемах, так и в соленых, вплоть до солености 60—80‰; в холодных водах, при температурах, близких к точке замерзания (—2°С), и в термальных источниках, при +52°С; у самой поверхности и в океанических впадинах глубиной свыше 8 тыс. м; в быстрых горных потоках и в прудах со стоячей водой; как в прозрачных, так и в чрезвычайно мутных водах, несущих огромное количество взвешенных веществ; в высокогорных озерах и подземных пещерах.
Рыбы обладают чрезвычайным разнообразием формы тела, от змеевидной и лентовидной до шаровидной и плоской. Размеры взрослых особей составляют от нескольких миллиметров до 15—20 м и более. Они не только плавают в толще воды, но и ползают по дну, иногда вылезая на сушу, прыгают по корням деревьев и даже летают над поверхностью воды. Одни рыбы совершают очень протяженные миграции на тысячи километров, другие всю жизнь живут в норках на литорали; ведут стайный, групповой и одиночный образ жизни. Чрезвычайно разнообразны способы размножения у рыб — от откладки икры до живорождения. Одни рыбы выметывают до 300 млн. икринок, у других происходит рождение всего одного или нескольких детенышей. Для «обучения» молоди некоторые виды образуют семейные группы.
Рыбы не только видят, слышат и обоняют, но с помощью органов боковой линии ощущают колебания воды. Они имеют электрические органы и электрорецепторы, способны осуществлять электроориентацию и электрокоммуникацию.
Как показывают палеонтологические исследования, современная фауна рыб сформировалась в основном уже в конце миоцена, т. е. приблизительно около 9—10 млн. лет назад. С конца плиоцена (около 4 млн. лет назад) ведет свое начало большинство обитающих на Земле видов рыб, которые существуют и до настоящего времени.
Четвертичный период по сравнению с другими периодами в истории Земли относительно короток и занимает всего около 2 млн. лет, но на протяжении его произошли существенные изменения в ихтиофауне различных водоемов, обусловленные как климатическими, так и антропогенными факторами. Даже на протяжении последнего временного отрезка этого периода — голоцена — наблюдались колебания климата, охватывающие значительные территории Евразии и Северной Америки, что отражалось и на составе фауны рыб.
В раннем голоцене (7700—12 000 лет назад) во всем Северном полушарии господствовал влажный и холодный климат. Граница тайги простиралась до арктических областей, на многие сотни километров севернее, чем теперь. Уровень Мирового океана быстро поднимался, реки были полноводными, и многие речные бассейны соединялись между собой системами озер или заболоценных водоемов. По данным палеоихтиологических исследований, в этот период в большинстве рек европейской части нашей страны широкое распространение имели лососевые и сиговые рыбы. В Сибири по численности преобладали такие холодноводные виды, как таймень, сиг, налим. Даже в реки Крыма в этот период заходили многие проходные и полупроходные рыбы, такие, как кумжа, рыбец, вырезуб; постоянно обитали в них и крупные пресноводные хищники — сом и судак.
В среднем голоцене (2500—7700 лет назад) на смену холодному и влажному климату приходит теплый атлантический и суббореальный. Ареалы многих теплолюбивых видов продвигаются далеко на север. В бассейне Белого моря (р. Онега) в это время водились такие виды, как красноперка, жерех, синец, сом. В Ладожском озере обитал бёрш, характерный для фауны Черноморско-Азовского и Каспийского бассейнов; встречались здесь также очень крупные сомы. Реки Крыма сильно усыхают, и из состава их фауны исчезают кумжа, вырезуб, сом и судак. Заметное потепление климата наблюдалось и на Дальнем Востоке, о чем, к примеру, свидетельствует обилие сазана в р. Шилке, а также появление в прибрежных водах залива Петра Великого большого количества японской скумбрии и ряда других теплолюбивых видов.
С окончанием среднего голоцена снова наступает похолодание, что отражается на составе ихтиофауны многих водоемов. Так, в р. Онеге исчезают вышеперечисленные теплолюбивые рыбы, а в Ладожском озере больше не встречается бёрш. По всей видимости, уже с этого времени в водоемах северо-запада резко сокращается численность сома и судака.
Четвертичный период ознаменовался и тем, что в начале его появился человек, вся последующая деятельность которого оказывала всё возрастающее влияние на природу. Поначалу антропогенное воздействие было еще ничтожным, но уже в неолите охота и рыболовство стали важнейшими занятиями древнего человека, появились достаточно совершенные орудия лова (сети, крючковые снасти, остроги), с возрастанием численности населения увеличивался список добываемых видов и общий вылов рыбы. Так, если в палеолитических стоянках человека, исследованных археологами на территории нашей страны, обнаружены костные остатки только 17 видов, то в эпоху неолита объектами промысла являлось уже 54 вида.
Наиболее ощутимо воздействие антропогенных факторов на ихтиофауну проявилось в последние полтора-два столетия. К числу основных факторов, под влиянием которых за столь короткий исторический отрезок времени произошли серьезные и часто необратимые изменения в водных экосистемах (количественного взаимоотношения видов в водоемах, структуры популяций и ряда биологических показателей отдельных видов, значительные изменения ареалов), следует отнести постоянно возрастающую интенсивность промысла, нарушение естественного гидрологического режима рек и озер, загрязнение водоемов отходами промышленности, а затем и крупномасштабного сельскохозяйственного производства, зарегулирование стока рек плотинами ГЭС и другими гидротехническими сооружениями.
Иллюстрированная энциклопедия рыб. Ст.Франк 1983 г.
Наиболее распространенная форма тела рыб торпедообразная или сигарообразная, сужающаяся к концу тела. Некоторые рыбы имеют угревидную форму тела, либо плоскую, сжатую с боков или в спинно-брюшном (дорзовентральном) направлении, иногда же почти круглую. Форма тела является следствием приспособления к определенному образу жизни: например, рыбы, живущие в толще воды, имеют округлое и вытянутое тело, а обитающие у дна, характерны уплощенной формой тела и т. д.
Тело рыбы состоит из головы, туловища и хвоста. Голова у отдельных видов рыб имеет весьма различающуюся форму. Рот по своему строению и назначению имеет в основном три типа: конечный, верхний, нижний. Положение рта тесно связано со способом питания. Рот окаймлен губами, у некоторых рыб очень толстыми, служащими для захвата пищи и в том числе для соскребывания водорослей с поверхности растений и камней. У некоторых сомовых рот имеет форму диска, служащего для присасывания к поверхности. У некоторых видов рыб прежде всего вокруг рта расположены усики, служащие органами осязания и для определения вкуса пищи. Глаза расположены чаще всего по сторонам головы, но иногда - на ее верхней части. У большинства рыб они достаточно велики, причем у молоди относительно большие, чем у зрелых особей. Глазные веки хорошо развиты только у акул. Некоторые виды пещерных рыб вторично слепы. В передней части головы перед глазами находится рыло, переходящее сзади в жаберные крышки. Ноздри парные, большей частью имеют форму слепо заканчивающихся ямок, иногда они трубовидно вытянуты. Туловище состоит из спинной и брюшной частей. Хвост обычно снабжен сильной мускулатурой. Хвостовой стебель - наименование хвостовой части тела между концом анального плавника и основанием хвостового плавника.
Плавники рыб на первых стадиях развития образуются в виде плавниковой каемки, которая позже распадается на отдельные костные плавниковые лучи, соединенные перепонкой или свободные. В передней части плавника обычно имеется 1 - 3 неразветвленных твердых луча, часто превращенных в шипы или иглы. Последующие лучи вилообразно ветвятся и называются мягкими. Плавники обычно подразделяются на парные и непарные. К парным относятся плавники грудные (им соответствуют передние конечности высших позвоночных) и брюшные (соответствующие задним конечностям). К непарным плавникам относятся спинной, анальный и хвостовой плавники, причем спинных и анальных плавников у некоторых видов рыб может быть несколько. Парные плавники часто имеют весьма различающееся строение. У некоторых лабиринтовых рыб, например, брюшные плавники длинные, нитевидные, преобразованные в органы осязания. У ползающих рыб грудные плавники служат для передвижения по влажной земле и вне воды. Некоторые виды рыб имеют огромные грудные плавники, приспособленные к парящему полету над водной поверхностью. У самцов акул и скатов брюшные плавники особым способом преобразованы в органы спаривания, так называемые mixipterygia. Органы спаривания у самцов живородящих рыб также возникли из перерожденного анального плавника и называются gonopodium. Так называемый жировой плавник имеется у лососевых, харациновых и некоторых сомовых рыб. Этот плавник различных размеров находится между спинным и хвостовым плавниками и не имеет плавниковых лучей.
Кожа рыб многослойна, с большим числом слизистых клеток. Выделяемая слизь обеспечивает легкое скольжение в воде и защищает кожу от инфекций. У большинства рыб тело покрыто чешуей. У акул это так называемые кожные плакоидные чешуйки с одним зубом на поверхности, покрытым эмалью. Другим типом чешуи является ганоидная чешуя в виде ромбических пластинок, свойственная осетровым, поверхность которых покрыта блестящим ганоином. У большинства рыб и в том числе карповых рыб преобладает циклоидная чешуя, представляющая собой гладкие костяные пластины овальной формы. Весьма сходна с ней ктеноидная чешуя, свободная задняя часть которой покрыта мелкими зубцами и шипами. Такая чешуя встречается у окуней. Два последних типа чешуи размещаются на теле рыб рядами, и, налегая друг на друга, покрывают его, как черепица крышу. У многих рыб тело голое, без чешуи, покрытое только толстой кожей (например, у сомовых).
Окраска рыб обусловлена разноокрашенными пигментными клетками, расположенными на границе эпидермиса и дермы. Клетки, названные эридоцитами, находятся глубоко в дерме. Они заполнены кристалликами гуанина, которые отражают падающие световые лучи и вызывают блестящую серебристо-белую окраску рыб. У некоторых рыб наблюдается так называемый ксанторизм. В их коже недостает всех видов пигментных клеток, кроме клеток с желтым и оранжевым пигментом. Глаза у этих рыб всегда черные. Если же в коже совсем нет пигмента, наблюдается так называемый альбинизм. Глаза у этих рыб обычно красные. Окраска в сочетании с формой тела имеет для рыб, в первую очередь, защитное значение. Примером так называемой мимикрии, доведенной до совершенства, может служить австралийский конек-тряпичник (Phyllopteryx eques), кожный покров которого представляет лентообразные кожистые выросты с коричневыми и красными полосами. По своей форме и окраске такие рыбы неотличимы от сплетения морских водорослей, среди которых они обитают. Окраска рыб характерна для вида, однако она меняется в зависимости от окружающей среды, в связи с возрастом рыбы, полом, физиологическим состоянием и даже настроением. Часто наблюдаются существенные различия в дневной и ночной окраске.
После общего внешнего знакомства с рыбами присмотримся к основным чертам внутреннего строения их тела. Основной опорой всего тела является скелет, костяк головы и позвоночный столб, состоящий из позвонков, число которых редко превышает 100, и то только у рыб угревидного строения тела. Мускулатуру полости тела поддерживают ребра. Скелет головы у рыб и рыбообразных находится на разной ступени развития. У круглоротых он хрящеобразный и не имеет челюстей. Наиболее сложное строение головы у костистых рыб, где скелет головы состоит из большого количества костей. Большинство рыб имеет зубы, расположенные на челюстях или на других костях рта. Там, где зубов нет, например, у карповых, на видоизмененной шестой жаберной дужке находятся глоточные зубы, которые служат для перетирания твердой пищи, например, раковин моллюсков и т. п.
Под жаберными крышками расположены жаберные дуги с жаберными лепестками и тычинками. Функция этого органа - дыхание и удержание пиши в ротовой полости. Внутренняя сторона жаберных дуг покрыта многочисленными жаберными тычинками; в зависимости от вида рыб эти тычинки по своему характеру и численности весьма различны. Жаберные тычинки образуют своего рода фильтр, при помощи которого рыбы отфильтровывают мелкую живую пищу (планктон) из воды, или удерживают ее во рту. У хищных рыб жаберные тычинки имеют форму бугорков с щитами, а пищевод может растягиваться и тем самым он приспособлен к поглощению значительного объема добычи.
Кроме жаберного дыхания у некоторых рыб имеются и своеобразные легкие, или дополнительные органы, позволяющие использовать для дыхания атмосферный кислород. Например, у лабиринтовых рыб верхняя часть первой жаберной дуги превратилась в небольшую полость, где в многочисленные тонкие кровеносные сосуды проникает атмосферный кислород, поступающий сюда вместе с воздухом, периодически заглатываемым рыбой ртом. Такой дыхательный аппарат называется лабиринтом. Иным типом является вспомогательное кишечное дыхание некоторых сомов и вьюнов, которые заглатывают пузырьки воздуха.
Плавательный пузырь - орган гидростатический. Эмбрионально он образуется выпячиванием спинной части стенки пищевода, тогда как легкие образуются выпячиванием брюшной стороны. Настоящие легкие имеются только у двоякодышащих рыб. Связь пищевода с плавательным пузырем у некоторых рыб сохраняется и во взрослом состоянии (например, у карповых). У других рыб (например, окуневых) плавательный пузырь отделен от пищевода, а наполнение газом и выравнивание давления с окружающей средой регулируется специальной железой, пронизанной сетью кровеносных капилляров, находящейся в стенке плавательного пузыря. У некоторых рыб плавательный пузырь вообще отсутствует. Часто плавательный пузырь тесно связан со слуховыми и гидростатическими органами. Это или прямая связь слухового аппарата с плавательным пузырем, (например, у трески), или с помощью так называемого Веберова аппарата, т. е. нескольких косточек в первой камере плавательного пузыря, имеющегося прежде всего у карповых, сомов, харацин и электрических угрей.
Рыбы - позвоночные существа с изменяющейся температурой тела (пойкилотермные) в зависимости от температуры окружающей среды, т. е. воды, в которой они живут. Центральный орган кровообращения у рыб - сердце - расположено в передней нижней части тела. Кровеносная система замкнутая и состоит и артерий, вен и капилляров. В сердце поступает лишенная кислорода кровь, которая затем идет к жабрам, а оттуда кровь уже обогащенная кислородом поступает к различным органам тела.
Нервная система у рыб достаточно совершенна. Ее образуют головной мозг, спинной мозг и большое число нервов. Большую часть головного мозга занимают средний мозг и мозжечок, но его величина относительно невелика.
Важным органом ориентации у рыб является так называемая боковая линия. Это в сущности каналик, идущий по бокам тела и пронизывающий ряд чешуек. В нем находятся чувствительные клетки, реагирующие на движение и состояние окружающей среды. Боковая линия часто бывает неполной по бокам, а на голове она нередко разделяется на ряд ветвей. Эта линия у рыб представляет своеобразный «радар». Она позволяет определить физические изменения водной среды и способствует устойчивости положения рыбы в воде, помогая сохранять ориентацию и в мутной воде, и при потере зрения.
Большинство рыб мечет икру, оплодотворение которой происходит вне тела. Только у живородящих рыб оплодотворение внутреннее. Акулы откладывают одно или несколько яиц, заключенных в роговую капсулу Обыкновенно эти довольно крупные капсулы имеют четырехугольную форму с нитевидными выростами, которыми они прикрепляются к водорослям или камням на дне. У некоторых представителей семейства Isuridae они достигают 22 см и являются самыми крупными из известных яиц позвоночных. Многие акулы живородящие. Рыбы обычно раздельнополы. Однако и тут исключение подтверждает правило. У некоторых костистых рыб двуполость, гермафродитизм - явление обычное; например, у морских окуней семейства Serranidae.
В зависимости от отношения к среде все рыбы делятся в отдельные периоды своего жизненного цикла на морских, пресноводных и проходных рыб. К последним относятся рыбы, живущие как в пресной, так и в морской воде. Кроме того, различают рыб пелагических, живущих в толще воды, а также придонных, живущих у дна. Для пресноводных рыб важным фактором является проточность воды. Различают рыб реофильных, живущих в течении рек, и лимнофильных, обитающих в стоячей воде, в озерах, омутах, прудах. Некоторые рыбы меняют среду обитания, и их очень трудно причислить к одной из приведенных категорий. Проходные рыбы в основном принадлежат к двум типам:
1. рыбы, живущие море и только в период нереста идущие в реки (анадромная миграция - например, лосось);
2. рыбы, проводящие большую часть жизни в реках, а на время нереста уходящие в море (так называемая катадромная миграция, например, угорь).
К числу важнейших физических факторов, оказывающих влияние на расселение рыб, относятся температура и соленость воды. Морскими течениями рыбы и особенно их икра могут разноситься на значительные расстояния; вместе с тем, морские течения могут явиться непреодолимым препятствием - барьером для распространения некоторых видов рыб. Для некоторых рыб свойственен так называемый коменсализм; рыба-прилипало сопровождает акулу, питается остатками ее пищи, присасывается к ней и таким образом транспортируется акулой на большие расстояния, Интересны часто встречающиеся отношения симбиотического характера между видами семейства Pomacentridae и морскими анемонами (актиниями). Эти рыбы скрываются в актиниях и живут с ними в симбиозе, в то время как рыб других видов актинии оглушают и пожирают. Паразитизм у высших рыб встречается редко. Например, сомик из рода Vandellia паразитирует в жаберной полости крупных сомоз Sorubim, либо Platystoma. У него узкое длинное тело, которое даст ему возможность проникнуть между жаберными дугами хозяина. Присосавшись к жабрам хозяина, он питается его кровью. В некоторых областях Бразилии этот сомик оказывается опаснейшей рыбой. Он проникает в мочевые пути купающихся людей. Там он закрепляется с помощью шипов и, таким образом, не имея возможности двигаться, вызывает воспаление, которое кончается отравлением крови и смертью. Некоторые авторы объясняют это тем, что он привлекается хемотоксически, т. е. запахом мочи. Скорее это будет реакция тигмотоксическая, т. е. он стремится плыть против течения мочи, подобно тому, как он проникает в жабры сомов, привлекаемый выбрасываемой при дыхании водой.
Половой диморфизм - частое явление у рыб. Чаще всего он выражается появлением на голове и на теле самцов (преимущественно период нереста) так называемой нерестовой сыпи - рогообразных кожных бугорков, у карпообразных рыб. Самцы большинства рыб отличаются от самок формой и размерами плавников, длиной усиков, расстоянием между глаз, наличием горба за головой и рядом других признаков.
Надкласс Челюстноротые (Gnathustomata)
Вторым надклассом являются челюстноротые (Gnathostomata). Эта систематическая единица включает в себя 4 класса рыб - акулообразных (Elasmobranchii), слитночерпных или цельноголовых (Holocephali), двоякодышащих (Dipnoi) и, наконец, класс высших рыб - Теleostomi, к которому принадлежит большая часть ныне живущих рыб.
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
НАДКЛАСС ЧЕЛЮСТНОРОТЫЕ (GNATHOSTOMATA)
Челюстноротые — преобладающая на
нашей планете группа позвоночных
животных. Они составляют 99,8% всего
числа видов этого типа животных, заселяя
воды и сушу. В водной среде они
представлены двумя классами рыб (хрящевые
и костные рыбы), а на суше — четырьмя
классами наземных животных
(земноводные, пресмыкающиеся, птицы и
млекопитающие).
Важнейшие черты строения челюстноротых позвоночных: наличие челюстного
аппарата в виде хватающих челюстей,
развившихся из жаберных дуг; парные
конечности в виде парных плавников,
пятипалых (исходно) конечностей или крыльев;
наличие костной или хрящевой ткани в
скелете; парные ноздри; три полукружных
канала во внутреннем ухе.
Самыми примитивными челюстноротыми
являются рыбы — хрящевые и костные.
Рыбы — это постоянно живущие в воде
позвоночные животные, дышащие жабрами
и передвигающиеся с помощью плавников.
Сто лет назад всех живущих в воде
похожих на рыб животных считали рыбами
и ученые объединяли их в один класс
«Рыбы». Исследования показали, однако,
что, например, ланцетник — это не рыба,
а не имеющее черепа, бесчерепное, очень
примитивное животное, близкое по
строению к предкам всех позвоночных вообще,
но имеющее черты строения и
беспозвоночных, и низших хордовых. Точно так
же миноги и миксины не рыбы, а
рыбообразные круглоротые животные, близкие
к предкам всех черепных позвоночных,
не имевшим ни парных конечностей, ни
хватающих челюстей. В истории развития
типа позвоночных животных настоящим
рыбам предшествовали рыбообразные
животные — Бесчерепные (Acrania),
Бесчелюстные, или Круглоротые (Agnatha), и
Панцирные (Placodermi).
Сотни миллионов лет назад рыбы были
представлены множеством групп и видов,
вымерших задолго до нашей эры. Сейчас
бесчерепные представлены ланцетниками
(класс Ланцетники — Amphioxi),
бесчелюстные — только миногами и миксинами (класс Круглоротые — Cyclostomata),
а панцирные рыбы полностью вымерли,
хотя в далекие силурийский и девонский
периоды, за 420—350 миллионов лет до
нашей эры, они были преобладающими
группами позвоночных животных.
Ныне живущих настоящих рыб, к
которым не причисляют сейчас ни
ланцетников, ни круглоротых, разделяют на
несколько классов, различия между
которыми не меньше, чем между классами
наземных позвоночных — земноводных,
пресмыкающихся, птиц и млекопитающих.
В этой книге мы различаем два класса
настоящих рыб — класс Хрящевые рыбы
(Chondrichthyes) и класс Костные рыбы
(Osteichthyes).
Рыбы пресных вод СССР и сопредельных стран. Л.С.Берг 1948 г.
Ветвь GNATHOSTOMATA. — Челюстные
Есть челюсти. Полукружных каналов три. Жаберный скелет у взрослых, а если у взрослых его нет, то в эмбриональном состоянии, типичного строения.
Надкласс PISCES. — Рыбы
Водные челюстные, обладающие жабрами во взрослом состоянии. Парные конечности, если они есть, никогда не бывают типа пятипалой конечности. Непарные плавники поддерживаются специальным скелетом. Есть только внутреннее ухо.
Класс Elasmobranchii. Внутренний скелет хрящевой, но вещество хряща может пропитываться известью. Кожных костей нет ни на голове, ни на челюстях, ни на плечевом поясе. Череп гиостилического или амфистилического типа. 5—7 пар жаберных дуг и жаберных щелей. — Акулы и скаты.
Класс Holocephali. Как Elasmobranchii, но череп голостилический (palatoquadratum слито с черепом). 4 пары жаберных щелей, прикрытых с каждой стороны кожной жаберной крышкой. Есть pharyngohyale. — Химера и др.
Класс Dipnoi. Внутренний скелет более или менее окостеневший. Кожные кости есть на черепе, на челюстях и на плечевом поясе. Наружное жаберное отверстие одно, прикрытое костной жаберной крышкой. Парные плавники с членистой серединной осью. Palatoquadratum слито с черепом. Плавательный пузырь превращен в легкое (или в легкие). Есть внутренние ноздри.
Класс Teleostomi. Внутренний скелет более или менее окостеневший. Кожные кости есть на черепе, на челюстях и на плечевом поясе. Наружное жаберное отверстие одно, прикрытое жаберной крышкой. Парные плавники не имеют серединной членистой оси. Palatoquadratum не слито с черепом.
Этот класс, заключающий громадное большинство ныне живущих рыб, разделяется на два подкласса: Crossopterygii и Actinopterygii. К числу Crossopterygii (кистеперые) относятся преимущественно вымершие рыбы. До недавнего времени к кистеперым причисляли и Polypterus. Но это недоразумение: у Polypterus нет основного признака, кистеперых — серединной членистой оси в парных плавниках. В 1939 г. у берегов южной Африки открыта настоящая кистеперая рыба, Latimeria, из группы Coelacanthi.
Actinopterygii раньше делились на три группы: Chondrostei, Holostei и Teleostei. К Chondrostei из ныне живущих относятся семейства Polypteridae, Acipenseridae и Polyodontidae, к Holostei — роды Amia и Lepidosteus, к Teleostei — все остальные Actinopterygii. Но если принять во внимание ископаемые формы, то можно убедиться, что Chondrostei постепенно переходят в Holostei, а эти — в Teleostei.