(Vertebrata J-B. Lamarck, 1801) Подтип Вертебраты, Позвоночные, Subphylum Vertebrata J-B. Lamarck, 1801
Материал подготовил Фактистов И. Е., 2020 г. Сайт www.fishbiosystem.ru
Позвоночные (лат. Vertebrata) — подтип хордовых животных. Доминирующая (наряду с насекомыми) на земле и в воздушной среде группа животных. Обилие и разнообразие видов позвоночных делают их важнейшим фактором экологической среды.
Всех остальных животных объединяют в несистематическую группу беспозвоночные. Использование этого термина оправдывается лишь той важной ролью, которую играют позвоночные в современном животном мире, так как в остальном группа беспозвоночных объединяет организмы, зачастую не имеющие практически ничего общего.
Позвоночные — высший подтип хордовых. По сравнению с низшими хордовыми — бесчерепными и оболочниками (3 класса: Appendicularia — Аппендикулярии, Ascidiacea — Асцидии, Thaliacea — Сальпы) — они характеризуются значительно более высоким уровнем организации, что наглядно выражено как в их строении, так и в физиологических отправлениях. Если низшие хордовые ведут сидячий (прикреплённый) или малоподвижный образ жизни, то поведение позвоночных несравненно более сложно. Среди них нет видов с сидячим образом жизни; они перемещаются в широких пределах, активно разыскивая и захватывая пищу, спасаясь от преследования врагов, разыскивая для размножения особей другого пола. Активному образу жизни представителей подтипа отвечает высокий уровень развития их нервной системы.
Позвоночные появились не позднее ордовика, а в юре существовали уже представители всех известных ныне их классов. Общее число современных видов около 40 тыс.
О начальных этапах эволюции позвоночных известно мало. И первые их формы, и их непосредственные предки почти наверняка были мягкотелыми организмами, похожими на современных ланцетников и практически неспособными сохраняться в ископаемом состоянии. Такая возможность возникла только после появления костей, чешуй и зубов. По всей видимости, предпосылки для фоссилизации костей сложились значительно позже первоначальной радиации позвоночных.
В 2014 году в силурийских отложениях древностью не менее 423 млн лет найдена Megamastax amblyodus — костная рыба длиной до метра; на 2014 год она считается первым позвоночным хищником, специализирующимся на поедании других позвоночных.
Филогения позвоночных во многом не ясна. В связи с этим в их составе обычно выделяют 7—13 и даже больше классов позвоночных. При этом земноводные, пресмыкающиеся, птицы и млекопитающие — общепризнанные классы, и различия обычно заключаются в количестве и составе остальных классов. Ниже перечислены 13 классов, в том числе пять вымерших.
Классы позвоночных:
• Круглоротые (Cyclostomata): миксины и миноги. В современной систематике часто разделяются на два отдельных класса: Myxini и Petromyzontida (Cephalaspidomorphi) соответственно.
• † Конодонты (Conodonta). По строению тела близки к круглоротым, однако имели более сложное строение ротовой полости.
• Бесчелюстные (Agnatha) [парафилетический класс]. Иногда в него включают — в качестве подкласса — и ныне живущих миног, иногда вообще не считают законной таксономической группой, подразделяя на несколько самостоятельных классов. Так, в классификации Дж. С. Нельсона (2006) фигурируют классы Pteraspidomorphа (или Diplorrhina), Anaspida, Thelodonti и Cephalaspidomorphi (или Monorrhina).
• Надкласс Рыбы (Pisces)
• † Плакодермы (Placodermi)
• Хрящевые рыбы (Chondrichthyes): акулы, скаты и химерообразные.
• † Акантоды (Acanthodii)
• Лучепёрые (Actinopterygii)
• Лопастепёрые (Sarcopterygii) [парафилетический класс]
• Надкласс Четвероногие (Tetrapoda)
• Земноводные или Амфибии (Amphibia): лягушки, жабы, тритоны и др.
• Пресмыкающиеся или Рептилии (Reptilia): черепахи, крокодилы, клювоголовые, ящерицы и змеи [парафилетический класс]
• Птицы (Aves)
• † Синапсиды (Synapsida) [парафилетический класс]
• Млекопитающие или звери (Mammalia)
Кроме того, выделяют различные таксономические и нетаксономические группы позвоночных (в алфавитном порядке):
• Амниоты (Amniota) — высшие позвоночные. Группа включает в себя пресмыкающихся, птиц, синапсид и млекопитающих.
• Анамнии (Anamnia) — низшие позвоночные; все классы челюстноротых, не входящие в группу амниот.
• Костные рыбы (Osteichthyes) — высшие рыбы (около 96 % всех видов рыб). Ранее считался классом, в современной систематике обычно трактуется как объединение двух сестринских групп: классов Лучепёрые (Actinopterygii) и Лопастепёрые (Sarcopterygii).
• Инфратип Челюстноротые (Gnathostomata). Клада, включающая надклассы рыб и четвероногих.
Ставриды Мирового океана (род Trachurus). В.В.Некрасов 1994 г.
У многих видов рыб соотношение полов в нерестовой части популяции близко к равному [Никольский, 1961; Правдин, 1966]. И вообще в популяциях многих двуполых организмов соотношение самцов и самок примерно равное [Пианка, 1981]. Для полной точности различают соотношение полов: первичное (пренатальное, или эмбриональное), вторичное - соотношение полов у потомства при рождении (для рыб - постэмбриональная и мальковая фазы) и третичное - соотношение полов у взрослых, размножающихся особей [Геодакян и др., 1967]. Обычно, когда специально не оговаривают, под соотношением полов в популяции имеют в виду третичное соотношение полов. Ч.Дарвин полагал, что соотношение полов, равное 1:1, выгодно группе, так как в пределах каждого пола оно до минимума сокращает конкуренцию за полового партнера [цит. по: Пианка, 1981]. Р.А.Фишер [Fisher, 1930] показал, что для видов, выращивающих свое потомство, наилучшие результаты получаются, если в возрасте половой зрелости достигается соотношение самцов и самок 1:1. Другими авторами было высказано мнение, что соотношение полов 0,5 (доля самцов в популяции) в репродуктивном возрасте максимально облегчает встречу особей противоположного пола и способствует понижению степени инбридинга [Геодакян и др.,1967].
И тем не менее, несмотря на все перечисленные преимущества равного соотношения полов, мы редко встречаем популяции, в которых соотношение полов равно 1:1. Например, по данным Е.К.Суворова (1948), это соотношение следующее. Волжский лещ: самцов 47,0% - самок 53,0%; треска Охотского моря: самцов 41,4-50,5 - самок 58,6-49,5%.
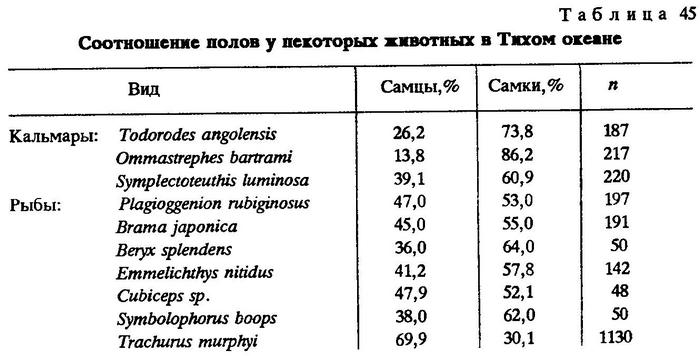
По нашим наблюдениям в 1982 г. в 4-м рейсе НПС "Возрождение", в Тихом океане за пределами 200-мильных экономических зон соотношение полов у большинства различных животных было таким, что, как и в примере Е.К.Суворова (1948), в популяциях преобладали самки. Лишь в популяции перуанской ставриды значительно преобладали самцы (табл.45).
Разумеется, у всех перечисленных видов не всегда и не везде такое же соотношение полов, которое мы обнаружили у популяций, обитающих в открытых водах юго-западной части Тихого океана. Вероятно, у других популяций этих видов в других районах соотношение полов будет несколько иным. Известны виды рыб, у которых соотношение полов в различных популяциях сильно различается. Так, например, у обыкновенного окуня (Perca fluviatilis ) в разных озерах соотношение самцов к самкам варьирует от 1:1 до 1:9 [Никольский, 1961]. В популяциях серебряного карася (Carassius auratus), обитающих в водоемах Китая и Японии, соотношение полов равно 1:1, но в бассейне Амура - 1:3, а на Урале, Северном Кавказе и в европейской части России существуют популяции без самцов. Г.В.Никольский (1965) высказал мысль, что, очевидно, бессамцовость здесь служит показателем хорошего состояния популяции, поскольку по мере ухудшения условий жизни в популяциях этого карася появляются самцы. Любопытно, что аналогичная картина наблюдается и у различных видов и популяций низших ракообразных, у которых есть все переходы от чистого партеногенеза до популяций с соотношением полов 1:1. При этом обнаруживается, что чем стабильнее условия среды, тем меньше в популяции самцов [Геодакян, Кособутский, 1969а]. А в практике разведения артемии для нужд рыбоводства уже сложилось такое положение, что появление самцов в садках является индикатором ухудшения условий содержания культуры.
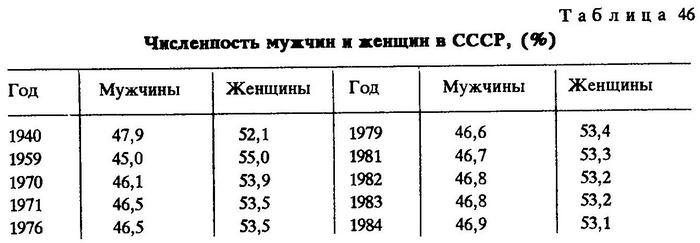
Результаты наблюдений и экспериментов по изучению относительной смертности полов у видов птиц, бабочек, млекопитающих, растений не оставляют сомнений, что повышенной смертностью у многих из этих видов отличаются самцы [Светлов, Светлова, 1950; Левин, 1951; Лэк, 1957; Урланис, 1965]. Подобная закономерность наблюдается даже у человека. Так, например, численность мужчин в СССР (табл.46) была несколько меньше, чем женщин [СССР в цифрах, 1984].
Коротко говоря, хотя в биологии не существует понятия "доказательство", как, скажем, в математике с ее заранее принятыми постулатами, собрано множество фактических данных, свидетельствующих в пользу того, что повышенная смертность особей мужского пола - явление универсальное. Вряд ли это случайно. По-видимому, среди многих факторов, связанных с дифференциацией полов, повышенная смертность мужского пола как общебиологическое явление тесно связана с эволюционными преимуществами раздельнополости. И вообще следует отметить, что неясен эволюционный смысл не только высокой смертности мужского пола, но и, собственно, самого явления дифференциации на два пола. Какие преимущества связаны с ним?
Все приводимые преимущества полового размножения (возможность избежать инбридинг, обеспечить генетическое разнообразие и замещение вредных мутаций и т.д.) сводятся только к комбинации и поэтому целиком относятся к процессу скрещивания, а не дифференциации. Получается, что большинство существующих теорий легко объясняют преимущества раздельнополого размножения, предполагающего и скрещивание и дифференциацию, перед бесполым, где отсутствует и скрещивание и дифференциация. В то же время они не в состоянии объяснить эволюционную прогрессивность раздельнополого размножения (со скрещиванием и дифференциацией) по сравнению с гермафродитизмом (с наличием скрещивания, но без дифференциации). Более того, если придерживаться существующего объяснения, приходится признать гермафродитный способ размножения эволюционно более выгодным, чем раздельнополый, так как генетическое разнообразие, обеспечиваемое гермафродитными способами, при одинаковой численности популяций примерно в два раза превосходит возможности раздельнополого способа. Действительно, если сравнить комбинаторные возможности гермафродитной и раздельнополой популяций одинаковой численности N, то в первом случае мы получим N(N-l)/2 = NN/2, а во втором N/2 х N/2 = NN/4 возможных вариантов скрещивания. Однако все прогрессивные в эволюционном смысле формы (млекопитающие, птицы, насекомые и др.) размножаются раздельнополым способом. Очевидно, разделение полов дает виду какие-то дополнительные преимущества. Какие же это преимущества?
В.А.Геодакяном (1965, 1974) была предложена концепция дифференциации полов, рассматривающая ее как специализацию по двум главным альтернативным аспектам эволюционного процесса - сохранения (консервативный) и изменения (оперативный) генетической информации популяции в ее взаимоотношениях со средой. Можно сказать, что дифференциация полов - это специализация по двум основным формам естественного отбора: стабилизирующего и движущего. Женский пол больше реализует тенденцию стабилизирующего отбора, а мужской пол - движущего. Иными словами, поток экологической информации к популяции передает больше мужской пол, а поток генетической информации (от поколения к поколению) - женский. Разделение эволюционирующей системы (популяции) на две подсистемы, одна из которых теснее связана со средой, поэтому более изменчива и подвижна (мужской пол), а другая более изолирована от среды, в результате чего более устойчива и стабильна (женский пол), повышает эволюционную устойчивость системы в целом. Концепция рассматривает повышенную смертность мужского пола как выгодную для популяции форму информационного контакта со средой, как плату за новую информацию, получаемую популяцией от среды, о происшедших в ней изменениях. Информационный контакт популяции со средой осуществляется через элиминацию вредным фактором среды части особей популяции.
Такая трактовка позволила по-новому осмыслить основные характеристики раздельнополой популяции (дисперсию полов, половой диморфизм и соотношение полов) и вскрыть некоторые связи их с эволюционными параметрами вида и среды.
Во-первых, согласно изложенной концепции, фенотипическая дисперсия мужского пола должна быть больше, чем женского. Более широкая фенотипическая дисперсия мужского пола обеспечивает более тесный информационный контакт его со средой и проявляется в виде повышенной смертности особей мужского пола. Такая форма связи со средой (ценой высокой смертности особей мужского пола) выгодна для популяции в целом. Она позволяет за новую информацию об изменениях, происшедших в среде, платить элиминацией мужских особей, а так как в панмиктной популяции каждый самец может оплодотворить много самок, то высокая смертность самцов не отражается на численности потомства.
Во-вторых, половой диморфизм по признаку (разница между средними значениями признака) удается связать с эволюционной тенденцией изменения этого признака. Такая связь вытекает из авангардной роли мужского пола, которого эволюционные изменения затрагивают в первую очередь.
И, наконец, вторичное соотношение полов рассматривалось ранее как константа, характерная для данного вида. С точки зрения нового подхода соотношение полов в популяции - переменная величина, тесно связанная с условиями среды и определяющая эволюционную пластичность вида. Очевидно, соотношение полов в популяции отражает специфику взаимосвязей популяции данного вида со средой. Оно не остается постоянным и в различных частях ареала вида. Половая структура популяции вида - такое же его приспособительное свойство, как и все другие признаки и свойства вида [Никольский, 1965]. Именно поэтому в стабильной эволюционной ситуации "вековой" оптимум соотношения полов устанавливается на относительно низком уровне (меньше самцов), а некоторые виды или популяции вида даже полностью утрачивают самцов.
В связи с этим большой интерес вызывает обнаруженное нами значительное преобладание по численности самцов у перуанской ставриды в юго-западной части Тихого океана (см. табл.45).
Известно, что распространение большинства видов рода Trachurus ограничено неритическими зонами, пределы которых они не покидают на протяжении всего жизненного цикла. Лишь несколько видов этого рода, среди них и перуанская ставрида, встречаются не только на шельфе, но и на расстоянии сотен и даже тысяч миль от побережья.
Таким образом, если концепция В.А.Геодакяна верна и в стабильной эволюционной ситуации "вековой" оптимум соотношения полов представлен преобладанием самок, то мы должны сделать вывод, что популяция перуанской ставриды в юго-западной части Тихого океана обитает в лабильных условиях. Однако, согласно концепции В.А.Геодакяна, при ухудшении условий среды в популяции должны гибнуть в большем количестве самцы, и в связи с этим возникает вопрос: почему самцов в популяции, несмотря на это, больше?
Чтобы ответить на этот вопрос, важно знать механизм изменения соотношения полов, в какую сторону и при каких условиях происходит изменение половой структуры.
Прежде всего следует обратить внимание на преобладание самцов в популяциях перуанской ставриды, обитающих не только в океанической эпипелагиали юго-западной части Тихого океана, но и у обитающих на шельфе. Наши наблюдения показали преобладание самцов в океанических популяциях перуанской ставриды, что имеет место в течение нескольких поколений и в различных районах.
Конечно, преобладание самцов в скоплении перуанской ставриды в юго-западной части Тихого океана, на периферии ареала вида, можно объяснить, по-видимому, следствием миграции от Американского континента в сторону Новой Зеландии. Известно, что в период миграций иногда замечается переформирование соотношения полов в скоплениях рыб, так как самцы активнее и подвижнее самок [Шмидт, 1936; Асланова, 1952]. Однако такого объяснения явно недостаточно, когда мы знакомимся с соотношением полов скоплений ставриды в южном подрайоне юго-восточной части Тихого океана, где в течение всего года последовательно встречаются нерестующиеся, нагульные и преднерестовые особи ставриды. К тому же мы располагаем данными из шельфовых участков ареала этого вида, полученными в 1973 г. в 12-м рейсе НПС "Академик Книпович" в водах Чили. Эти данные говорят о том, что в то время на шельфе в популяции перуанской ставриды тоже преобладали самцы. Соотношение полов составляло 51,7% : 48,3% (n = 1716).
Наконец, чтобы убедиться в том, не является ли преобладание самцов в популяциях во всем роде Trachurus обычным, мы привлекли для сравнения наши материалы по другим видам этого рода, собранные в разные годы в Индийском и Атлантическом океанах (табл. 47).
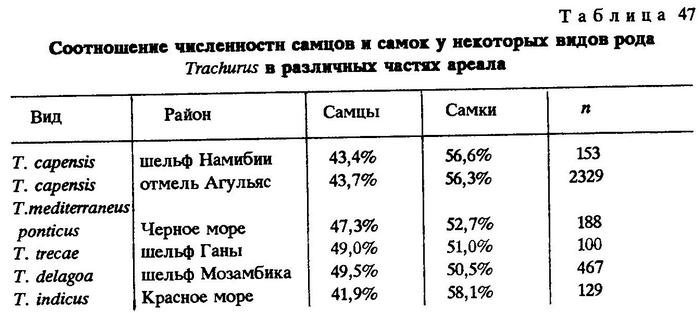
Как видно, у большинства видов рода Trachurus в популяциях преобладают самки. Итак, объяснение высокого уровня соотношения полов у океанических популяций перуанской ставриды только миграциями представляется недостаточным и неубедительным. Возможно, это является следствием пока нам неизвестных суровых условий среды обитания вида. Преобладание самцов в популяции перуанской ставриды, обитающей на шельфе, очевидно, свидетельствует об изменчивой среде ее обитания, что, по-видимому, вынуждает ставрид осваивать новые биотопы в открытом океане. Известно, что перуанская ставрида, как и калифорнийская, выходит далеко за границу континентального шельфа, на расстояние многих сотен миль от него [Roedel, Fitch, 1952; Ahlstrom, 1956; Алеев, 1957а]. Н.В.Парин (1968), рассматривая в сравнительном аспекте состав ихтиофауны открытой и прибрежной эпипелагиали, пришел к выводу, что основная масса рыб, слагающих ихтиофауну открытой эпипелагиали, принадлежит к филогенетическим молодым группам. Очевидно, это справедливо и для перуанской ставриды.
Скопления перуанской ставриды, относительно недавно вышедшие в открытую эпипелагиаль, обладают высокой пластичностью, что дает возможность освоить новые биотопы, отвечая еще более высоким уровням соотношения полов, т.е. большим количеством самцов в популяции.
Те же тенденции наблюдаются и при более кратковременных изменениях среды в пределах одного поколения. В экстремальных условиях среды третичное соотношение полов падает, т.е. снижается количество самцов в популяции.
По-видимому, для объяснения описанной ситуации нужен иной подход.
Известен метод исследования изменений полового состава популяций рыб путем анализа размерно-половых соотношений [Замахаев, 1959]. Размерно-половыми соотношениями называют соотношения долей самцов и самок, приходящихся на каждую размерную группу вариационного ряда длин рыб. Применение этого метода исследований для видов рыб, у которых отсутствует половой диморфизм, удачно еще и тем, что позволяет одновременно анализировать изменение соотношения полов и в связи с возрастом, так как рост есть функция возраста. Суть этого метода заключается в том, что при большей продолжительности жизни особей одного пола, например, самок, среди крупных рыб обнаруживается значительное их преобладание.
Выделяют три типа размерно-половых соотношений рыб. Первый тип - равная численность особей обоих полов во всех размерных группах; второй тип - увеличение доли самок от незначительного в размерных группах мелких рыб до ста процентов в размерных группах наиболее крупных рыб; третий тип - обратный второму, уменьшение доли самок до нуля.
Конечно, эти три типа размерно-половых соотношений носят несколько условный характер. Известны случаи, когда в одних популяциях какого-либо вида имеет место соотношение полов первого типа, а в других популяциях того же вида - ближе ко второму. Однако несомненно, что распределение особей разного пола по размерным и возрастным группам у рыб является приспособительным свойством вида и отражает специфику взаимосвязей каждой его популяции со средой. На изменение условий жизни популяция закономерно отвечает изменением половой структуры [Замахаев, 1959; Никольский, 1965].
Считают, что у рыб первого типа во всех случаях гарантируется успешное оплодотворение икры, а у рыб второго типа преобладание самок среди крупных рыб обеспечивает большую плодовитость стада, поскольку крупные рыбы откладывают больше икры. Но если иметь в виду, что в панмикстной полигамной популяции каждый самец может оплодотворить много самок, придется признать, что второй тип соотношения полов более выгодный, чем первый, и останется только гадать, что мешает видам первого типа изменить соотношение полов на второй тип. Также считают, что соотношение полов третьего типа обычно бывает у тех видов рыб, самцы которых охраняют икру, хотя непонятно, почему в этом случае самцы обязательно должны быть крупнее самок. Биологическое значение размерно-половых соотношений у третьего типа рыб, не охраняющих икру, но у которых самцы преобладают среди рыб более крупных размеров, многие авторы затрудняются объяснить [Замахаев, 1959]. К тому же неясным остается, каким образом реагируют популяции соотношением полов на изменение условий среды: в каком случае популяция отвечает увеличением относительной численности самцов, а в каком - ее уменьшением.
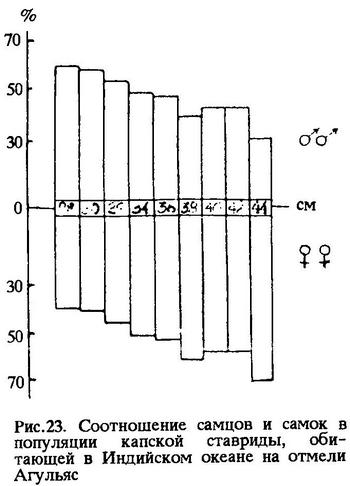
Интересно отметить, что у других видов рода Trachurus, обитающих на шельфе, например, у Trachurus mediterraneus ponticus, Trachurus trecae, Trachurus delagoa, Trachurus capensis размерно-половое соотношение второго типа. Наиболее наглядно это видно на примере популяции капской ставриды Trachurus capensis, обитающей на отмели Агульяс (рис.23) (n = 2329 экз.). Если при длине тела капской ставриды 38-42 см в возрасте 7-10 лет самцы составляли лишь 41,6-41,4%, а среди более крупных рыб длиной 42-44 см в возрасте 12 лет и более лет самцов было всего 28,6%, то среди рыб длиной 26-30 см в возрасте 3-4 года доля самцов составила 59,2-58,0%.
He только у ставрид, включая перуанскую, к концу жизни особей наблюдается увеличение количества самок. Так, например, соотношение полов у морской камбалы Pleuronectes platessa в Северном море следующее: среди молоди длиной 6-7 см на 100 самок приходится 150 самцов; чем крупнее камбала, тем меньше попадается самцов, а среди рыб длиной 67-70 см встречаются только самки. То же наблюдается и у других камбал [Суворов, 1948; Шатуновский, 1963]. Вообще второй тип соотношения полов характерен для очень многих видов рыб [Никольский, 1965]. Да и не только рыб. Д. Б. Гамильтон [цит. по Геодакяну, 1974] приводит обзор дифференциальной смертности полов для 70 видов, включая такие различные формы, как немотоды, моллюски, ракообразные, насекомые, паукообразные, птицы, рептилии, рыбы, млекопитающие. Согласно этим данным, у 62 видов средняя длительность жизни самцов меньше, чем у самок; у большинства остальных нет разницы между полами, и лишь в отдельных случаях длительность жизни самок короче, чем самцов. Второй тип соотношения полов характерен и для растений [Геодакян, Кособутский, 1969а,б].
Закономерности изменения соотношения полов в популяции с возрастом в основном сводятся к следующему. Во-первых, отмечается избыточное зарождение самцов. Во-вторых, у многих видов рождается больше особей мужского пола. В-третьих, в огромном разнообразии видов, как среди животных, включая человека, так и среди растений, наблюдается повышенная смертность особей мужского пола в результате воздействия вредных факторов среды [Геодакян, 1974]. Если бы не было селективного воздействия элиминирующих факторов среды на особей разного пола, то третичное соотношение полов не отличалось бы от вторичного. Точно так же вторичное соотношение полов образуемся из первичного под действием антенатальной смертности. Связующим же звеном между третичным соотношением полов (взрослых особей) и вторичным (при рождении), т.е. вероятностью рождения потомков того или другого пола у каждой отдельной пары особей, является, по-видимому, автономная отрицательная обратная связь, регулирующая соотношение полов при нарушениях его оптимума. Накопилось много научных сообщений о том, что такая связь действительно существует [Геодакян, 1974]. У разных видов организмов конкретный механизм отрицательной обратной связи может реализовываться по-разному, только принцип единый для всех. Так, например, у пчел из неоплодотворенных яиц развиваются самцы, а из оплодотворенных - самки. У некоторых растений связующим звеном является количество пыльцы, попадающей на рыльце цветка: много пыльцы - развиваются растения женского пола, мало пыльцы - развиваются растения мужского пола. Возможны и другие механизмы отрицательной обратной связи.
И вот на фоне изложенного неожиданным противоречием открывается перед нами картина размерно-полового соотношения в океанических популяциях перуанской ставриды к концу жизни в сторону увеличения доли самцов. Чем объяснить это нетипичное явление, когда при третичном соотношении полов с возрастом увеличивается относительная численность самцов? Почему самок с возрастом гибнет больше, чем самцов, и как это увязывается с изложенной выше концепцией В.А.Геодакяна, согласно которой все должно быть наоборот?
Рассмотрим схему действия элиминирующего фактора среды на популяцию. Любому вредному фактору среды соответствует определенное свойство организмов: низким температурам - морозостойкость, высоким - теплоустойчивость, длительному голоданию - способность к жиронакоплению и т.д. Все признаки организмов прямо или косвенно связаны с факторами внешней среды. Если изобразить кривой нормального распределения смертность особей популяции в зависимости от того или иного элиминирующего фактора, то мы тем самым получим кривую фенотипического распределения популяции по соответствующему признаку. Следовательно, чем больше дисперсия кривой смертности, тем больше дисперсия кривой фенотипического распределения и тем чувствительнее популяция к данному вредному фактору среды. И если сравнить две вариационные кривые смертности особей мужского и женского пола в популяции, то совершенно очевидно, что кривые будут различаться по любому элиминирующему признаку величиной дисперсии, т.е. степенью разнообразия по данному признаку и средними значениями вредного фактора среды или сопряженных с ними признаков, в результате чего гибнет ровно половина популяции. Дисперсия у особей мужского пола шире, а у особей женского пола больший эксцесс вариационной кривой. И разница эта есть не что иное, как половой диморфизм, иначе бы третичное соотношение полов не отличалось от вторичного. Таким образом, если первые жертвы элиминирующего фактора мужские особи, то это означает, что фенотипическая дисперсия у мужского пола больше, чем у женского.
Значит, более тесная связь особей мужского пола со средой обеспечивается большей величиной их фенотипической дисперсии по сравнению с особями женского пола. Типичной иллюстрацией к этому служит размерно-половое соотношение в популяции капской ставриды, обитающей на отмели Агульяс (рис.23). В этом случае кривые смертности имеют вид, изображенный на рис.24. По оси абсцисс - устойчивость к вредному фактору среды; по оси ординат - количество особей с данной устойчивостью. На рисунке видно, что кривая смертности женских особей имеет положительный эксцесс, а мужских - отрицательный. И это понятно, потому что у мужских особей разнообразных отклонений от моды популяции больше, чем у женских, а за счет этого образуется зона дискомфорта (Зд). Зона дискомфорта тем шире, чем популяция пластичнее. На рис.24а показано, что вредный фактор среды (Ф - фронт вредного фактора) элиминирует малую часть популяции (заштриховано на рисунке), состоящую преимущественно из самцов. Именно поэтому к концу жизни поколения в популяции капской ставриды остается больше самок. Такая форма связи со средой ценой высокой смертности самцов, как уже отмечалось выше, выгодна для популяции в целом.
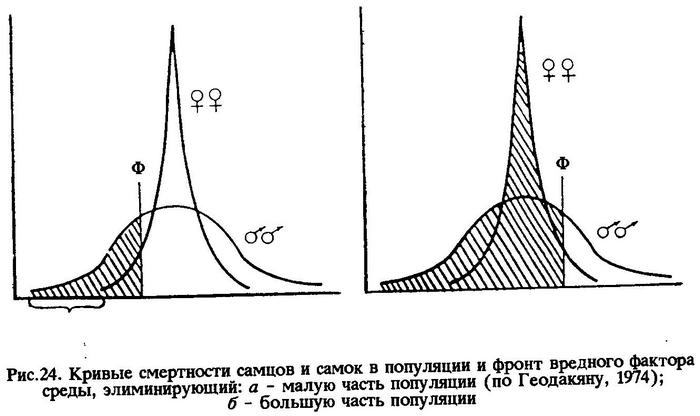
В случае с океаническими популяциями перуанской ставриды, в которых к концу жизненного цикла поколения остается больше самцов, мы обязаны были бы предположить, что, при прочих равных условиях, фенотипическая дисперсия больше у особей женского пола. И это явилось бы редчайшим исключением в природе. Но исключения здесь нет.
Принимая во внимание то, что популяция на изменение условий среды закономерно отвечает изменением половой структуры, мы вправе считать, что условия обитания популяций перуанской ставриды нестабильные. Нестабильные условия среды и обусловливают численное преобладание самцов. На океанические популяции перуанской ставриды вредный фактор (или факторы) среды, вероятно, действует постоянно и элиминирует большую часть популяции (см.рис.24б). Но так как фенотипическая дисперсия у самцов выше, чем у самок, то имеется широкая зона дискомфорта не только в левой части вариационной кривой, но и в правой. И, стало быть, при более мощном воздействии вредного фактора в правой части остается больше самцов. Именно поэтому к концу жизненного цикла поколения перуанской ставриды, обитающей в океанической части ареала, остается больше самцов.
В природе имеется много примеров, подтверждающих правильность нашего умозрительного заключения. Так, например, по устному сообщению И.Н.Горкина, было установлено, что в чистых реках Приморского края соотношение полов у гольца Nemacheilus barbatulus toni составляет 49% : 51%, т.е. самцов чуть меньше. В реках, в водах которых присутствуют соли тяжелых металлов (цинка, меди, свинца, кадмия). соотношение полов сдвигается в сторону увеличения количества самок - 41% : 59%. В тех реках, в которых концентрация солей тяжелых металлов высокая, при общем значительном снижении численности гольца, резко увеличивается количество самцов - 69% : 40%.
В случае с океаническими популяциями перуанской ставриды, в которых к концу жизненного цикла поколения остается больше самцов, мы обязаны были бы предположить, что, при прочих равных условиях, фенотипическая дисперсия больше у особей женского пола. И это явилось бы редчайшим исключением в природе. Но исключения здесь нет.
Принимая во внимание то, что популяция на изменение условий среды закономерно отвечает изменением половой структуры, мы вправе считать, что условия обитания популяций перуанской ставриды нестабильные. Нестабильные условия среды и обусловливают численное преобладание самцов. На океанические популяции перуанской ставриды вредный фактор (или факторы) среды, вероятно, действует постоянно и элиминирует большую часть популяции (см.рис.24б). Но так как фенотипическая дисперсия у самцов выше, чем у самок, то имеется широкая зона дискомфорта не только в левой части вариационной кривой, но и в правой. И, стало быть, при более мощном воздействии вредного фактора в правой части остается больше самцов. Именно поэтому к концу жизненного цикла поколения перуанской ставриды, обитающей в океанической части ареала, остается больше самцов.
В природе имеется много примеров, подтверждающих правильность нашего умозрительного заключения. Так, например, по устному сообщению И.Н.Горкина, было установлено, что в чистых реках Приморского края соотношение полов у гольца Nemacheilus barbatulus toni составляет 49% : 51%, т.е. самцов чуть меньше. В реках, в водах которых присутствуют соли тяжелых металлов (цинка, меди, свинца, кадмия). соотношение полов сдвигается в сторону увеличения количества самок - 41% : 59%. В тех реках, в которых концентрация солей тяжелых металлов высокая, при общем значительном снижении численности гольца, резко увеличивается количество самцов - 69% : 40%.
Иллюстрированная энциклопедия рыб. Ст.Франк 1983 г.
Рыбы и рыбообразные животные относятся к подтипу ЧЕРЕПНЫХ (Craniata), являясь наиболее примитивными представителями этой группы, и именно они понимаются под словом «рыба» в широком его понимании. В настоящее время к хордовым относится более 40 тысяч видов, причем рыбы составляют приблизительно половину, т. е. около 20 тысяч видов. О возникновении хордовых имеется много различных теорий. Упомянем сейчас некоторые из них. Некоторые исследователи ведут происхождение хордовых от МОРСКИХ ЧЕРВЕЙ (Annelida), прежде всего, от их свободно плавающих личинок, так называемых трохофор. Многие авторы считают, что хордовые произошли от другого класса червей, а именно от НЕМАТОД (Nematoda) или от НЕМЕРТИН (Nemertini). Мак Фарлан ведет хордовых от пресноводных немертин и обращает внимание на некоторые их общие признаки с бесчерепными, круглоротыми и, наконец, с рыбами. Другие авторы указывают на их связь с членистоногими. Ряд теоретических предположений пришлось пересмотреть в связи с установлением того, что планктонные ЩЕТИНОЧЕЛЮСТНЫЕ ЧЕРВИ (Chaetognatha), ИГЛОКОЖИЕ (Echinodermata) и РЫБЫ (Pisces) принадлежат в своем развитии к живым существам эмбрионально вторичноротым (Deuterostomii).
Дальнейшие исследования установили ряд общих признаков между названными выше группами. Северцев же, обращая внимание на некоторые анатомические признаки, создал подробную схему развития хордовых, ведя их от гипотетических ПРИМИТИВНЫХ ХОРДОВЫХ (Acrania primitiva), из которых возникают бесчерепные (далее не развившиеся), а также предки всех остальных хордовых, которых он называет Procraniata. От этих последних возникли две ветви, из которых одна заканчивается тупиком - рыбообразными круглоротыми (миноги, миксины), а вторая развилась далее, до сегодняшних ступеней развития собственно рыб, а именно акул (Elasmobranchii), ДВОЯКОДЫШАЩИХ (Dipnoi), кистеперых (Crossopterygii), ХРЯЩЕВЫХ ГАНОИДОВ (Chondrostei) и КОСТИСТЫХ (Teleostei).
Кроме костистых рыб, все остальные группы уже достигли наиболее высокого уровня своего развития в отдаленном прошлом; свидетельством тому является богатство форм, которые у этих групп находят палеонтологи в различных геологических эпохах. Наоборот, костистые рыбы представляют собой прогрессирующую, развивающуюся ветвь хордовых. С рыбообразными - круглоротыми палеонтологи не встречаются ни в одном геологическом периоде. Самыми примитивными представителями предков рыб считаются семейства Cephalaspidae, Pteraspidae, Соеlоlерidae и Drepanaspidae, жившие уже в палеозойскую эру, в силурийском и девонском периодах и достигавшие 20 - 60 см в длину. У них большая часть головы, груди, а иногда и всего тела, была покрыта прочным панцирем, уже содержащим костные клетки. Дальнейшая эволюция этих форм в геологические периоды трудно поддается изучению, так как отсутствует ряд промежуточных звеньев, связывающих отдельные группы.
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
ТИП ПОЗВОНОЧНЫЕ (VERTEBRATA)
ОБЩИЙ ОЧЕРК
Позвоночные характеризуются
наличием внутреннего осевого скелета,
двусторонней (двубоковой) симметрией тела,
характерным расположением основных
систем органов, двумя парами конечностей,
двуслойным строением покровов,
замкнутой кровеносной системой, сложно
устроенной — трубчатой — нервной системой,
высоким развитием органов чувств и
другими признаками.
Для типа позвоночных особенно
характерен внутренний скелет, основой которого
является прочный осевой стержень— хорда
или позвоночник. Эта опора скелета
простирается вдоль всего тела от переднего
до хвостового конца. Хорда — это
упругий стержень из пузырчатых клеток,
заключенных в прочный волокнистый чехол,
сохраняющийся в таком виде у
низших позвоночных — ланцетников — в
течение всей жизни. У нескольких групп
беспозвоночных животных (кишечнодышащие, оболочники) имеются в строении
зачатки хорды, в связи с чем их иногда
объединяют вместе с позвоночными в надтип
хордовых. Хорда имеется у зародышей
всех позвоночных. У большинства
позвоночных хорда во время развития
замещается костными позвонками; остатки ее
сохраняются у взрослых только в виде
упругих межпозвонковых дисков.
Скелет у большинства позвоночных построен
преимущественно из костной ткани,
которая образуется особыми костными
тельцами (клетками — остеобластами), и
растет и перестраивается в течение всей
жизни животного.
Позвоночник образован отдельными
костными позвонками, между которыми
расположены упругие линзовидные
межпозвонковые диски. Такая структура
придает одновременно прочность и упругость
этой основной опоре тела позвоночных
животных. Типичный позвонок состоит из
кольцеобразного или цилиндрообразного
тела позвонка и отходящих от тела
парных отростков — верхних и нижних дуг.
Концы верхних дуг обеих сторон
соединяются друг с другом, образуя
канал, в котором расположен спинной мозг.
Нижние дуги в области туловища
образуют короткие поперечные отростки, с
которыми сочленены ребра. В области хвоста
концы их соединяются внизу, образуя
канал, по которому проходят хвостовая
артерия и вена. Таким образом,
позвоночник не только служит опорой всего тела,
но его отростки и сочленяющиеся с ними
ребра образуют и защитную костную
грудную клетку — футляр для
важнейших систем органов позвоночных.
С передним концом позвоночника у всех
черепных позвоночных сочленена
черепная коробка. Она представляет собой
хрящевой или костный футляр, в средней
части которого помещается головной мозг,
а в боковых частях — капсулы парных
органов чувств: обоняния, зрения и
слуха. С черепной коробкой снизу сочленен
так называемый висцеральный скелет (от
латинского висцера — внутренности).
Он представляет собой исходно ряд дуг,
которые в эволюции позвоночных
преобразовались в скелет челюстей, подъязычную
кость и слуховые косточки.
Позвоночные имеют, как правило, две
пары конечностей — передние и задние, с
их опорными поясами — плечевым и
тазовым, связанными обычно с
позвоночником. Нет конечностей только у низших
позвоночных — ланцетников и круглоротых — и у вторично утративших их
змеевидных животных.
В соответствии со средой обитания
водные позвоночные — бесчерепные
(ланцетники), круглоротые (миноги и миксины)
и рыбы — дышат жабрами. Конечности у
рыб сформированы в виде плавников.
Тело имеет более или менее рыбовидную
или торпедовидную форму, без внешнего
обособления в нем шеи, и основным
органом движения у них служит хвост.
Наземные позвоночные — земноводные,
пресмыкающиеся, птицы и
млекопитающие — дышат легкими (только
развивающиеся в воде личинки земноводных имеют
жабры). В дополнение к голове, туловищу
и хвосту у наземных позвоночных
обособляется шея, благодаря чему голова
приобретает подвижность. Парные
конечности у наземных позвоночных
построены по принципу сложных рычагов, имеют
в основе пятипалое строение и служат у
большинства для передвижения по суше
и в воздухе.
Непарных конечностей,
соответствующих непарным плавникам водных
животных, у наземных позвоночных нет.
Кожа позвоночных — плотный и
прочный покров, обособляющий организм от
внешней среды, состоит из двух слоев:
поверхностного — надкожицы, или
эпидермиса, и нижнего — собственно кожи,
или дермы (от греческих слов эпина и дерма — кожа). Эпидермис у
ланцетников однослойный, у всех остальных
позвоночных — многослойный, причем у
водных — рыбообразных и рыб, а также у
земноводных он снабжен слизистыми
железами; у высших наземных позвоночных
наружный слой эпидермиса ороговевает,
образуя роговые чешуи — у
пресмыкающихся, перья — у птиц, волосы — у
млекопитающих. В эпидермисе высших
позвоночных также развиваются
разнообразные кожные железы: потовые, сальные,
слизистые.
Важнейшие системы внутренних органов
расположены вдоль осевого скелета —
хорды или позвоночника: над хордой —
центральная нервная система, под ней —
спинной ствол .кровеносной системы
(спинная аорта). По обе стороны вдоль
спинной аорты расположены органы
выделения — почки. Ниже этого спинного
комплекса систем органов, внутри туловища,
расположены брюшная и околосердечная
полости тела, содержащие органы
пищеварения п другие системы органов
(«внутренности») и сердце.
Система органов пищеварения
объединяет длинную, усложняющуюся у
высших классов позвоночных
пищеварительную трубку и несколько пищеварительных
желез, соединенных с пищеварительной
трубкой протоками. Пищеварительная
трубка подразделяется на ротовую полость,
глотку, пищевод, желудок, тонкую и
толстую кишки. Основные пищеварительные
железы — печень и поджелудочная железа.
Печень выделяет желчь, которая
способствует перевариванию жиров;
поджелудочная железа выделяет соки,
участвующие в переваривании белков и углеводов.
Участвует в пищеварении также слюна,
выделяемая в ротовую полость слюнными
железами.
Особое значение имеет глотка, из
стенок которой развиваются железы
внутренней секреции — щитовидная и зобная —
и которая всегда бывает связана с
органами дыхания. У водных позвоночных
боковые стенки глотки прободены
жаберными щелями, а у двоякодышащих рыб и
у наземных животных от ее брюшной
стенки отходит дыхательная трубка,
ведущая к легким.
Кровеносная система позвоночных
замкнутая, т. е. кровь циркулирует по сети
сосудов по всему телу, нигде не
застаиваясь в полостях — лакунах.
Центральный орган системы — сердце —
расположен на брюшной стороне. Только у
низших позвоночных— ланцетников — сердце
не сформировано и его функции выполняет
передний отдел брюшного сосуда. У
большинства рыб сердце двухкамерное, у
земноводных и высших позвоночных — трех-
камерное и четырехкамерное. Сердце —
сложно устроенный орган, снабженный
мощной мускулатурой и регулирующими
ток крови клапанами.
Ритмические сокращения мышц сердца
(пульсация) гонят кровь по артериям
к голове, головному мозгу, органам
дыхания, в спинную аорту. Кровь
расходится по утончающимся сосудам по всему
телу, отдавая тканям и органам кислород
(полученный ею при прохождении через
органы дыхания) и питательные вещества.
В почках и в печени она растекается
по мельчайшим сосудам — капиллярам,
через стенки которых протекает обмен
веществ, и вновь собирается в вены,
ведущие ее к сердцу. Это характерные для
позвоночных воротные системы печени и
почек. Кровообращение у высших позвоночных достигло большого
совершенства, обеспечивающего
поддержание собственной
постоянной температуры тела и
повышение энергетического
уровня жизнедеятельности.
Центральная нервная
система имеет вид трубки,
располагающейся обычно в канале,
образованном верхними дугами
позвонков над позвоночником.
В голове она расширяется и
усложняется, образуя головной
мозг, защищенный черепной
коробкой.
Головной мозг чрезвычайно
усложняется у высших
позвоночных, разделяясь на пять
основных отделов — передний,
промежуточный, средний,
продолговатый мозг и мозжечок,
в которых располагаются
основные центры восприятия
внешних и внутренних
раздражений, их переработки и
формирования ответных реакций и
поведения организма. Исходно, в
грубой схеме, передний мозг
содержит обонятельные и
осязательные центры, средний и
промежуточный — зрительные,
мозжечок — двигательные,
продолговатый — вкусовые,
слуховые и сейсмосенсорные
(восприятия движений воды). У
высших позвоночных структура
головного мозга усложняется
изгибами, складками,
многослойным расположением
взаимосвязанных разнородных
нервных клеток, развитием
проводящих путей и связей (комиссур).
Центры восприятий в мозгу
взаимосвязаны, особенно у высших
позвоночных, и поведение животного
определяется переработкой сочетаний
различных восприятий, подчас очень сложной,
вплоть до ассоциативной.
Спинной мозг представляет собой
толстостенную трубку, состоящую из серого
и белого мозгового вещества. Серое
мозговое вещество содержит нервные клетки
и нервные волокна, белое — одетые
белой мякотной оболочкой нервные
волокна.
От головного мозга отходит
от 8 до 12 пар головных
нервов, от спинного отходят метамерно спинномозговые нервы,
число которых соответствует
числу первоначальных
мышечных сегментов животного.
Отростки спинномозговых нервов
разветвляются по всему телу.
Органы чувств у
позвоночных животных достигают очень
большой сложности и
совершенства строения. Слабо развиты
они только у ланцетника. У
позвоночных имеются кожные
органы осязания, ощущений
движений воды (сейсмосенсорные),
вкуса; развиты органы
обоняния; имеются специальные
органы слуха, служащие
одновременно и органами
равновесия, а также специальные
органы зрения. Все эти парные
сложно устроенные органы
помещаются в специальных
впадинах черепных костей.
Органы зрения позвоночных
представлены парными
глазными яблоками. Плотная
наружная оболочка их — склера —
служит внешней опорой,
скелетом глаза. В передней части
глаза склера переходит в
прозрачную роговицу. Внутри
глазного яблока располагается
прозрачный линзовидный
хрусталик. Изнутри склера
выстлана тремя оболочками —
сетчатой (воспринимающая),
сосудистой (питающая) и
пигментной (изолирующая). На
границе с роговицей сосудистая
оболочка переходит в радужину,
диафрагмирующую отверстие зрачка.
Задняя камера глазного яблока заполнена
стекловидным телом, а передняя —
водянистой влагой. Через прозрачные части
глазного яблока (роговица, хрусталик,
водянистая влага и стекловидное тело)
проходят и преломляются лучи света.
Внутренний аппарат органа слуха и
равновесия (внутреннее ухо) представляет
собой сложно устроенный перепончатый
лабиринт, наполненный жидкостью — эндолимфой — и заключенный в костном лабиринте. Лабиринт состоит из трех
полукружных каналов, расположенных во
взаимно перпендикулярных плоскостях,
центрального протока, нижнего мешочка
и полого выроста, спирально
закручивающегося у млекопитающих,— улитки.
Наибольшей сложности и тонкости строения
органы слуха достигают у высших
наземных позвоночных — млекопитающих, у
которых в добавление к внутреннему
аппарату развивается также среднее и
наружное ухо. При единстве основного типа
строения позвоночные разных классов
существенно отличаются друг от друга
строением тела, характером развития, образом
жизни.
Рыбы постоянно живут в воде: от яйца
(икринки) до взрослых животных,
дышащих жабрами. Только двоякодышащие
рыбы наряду с жабрами имеют и легкие,
благодаря которым могут дышать
атмосферным воздухом. У земноводных
развитие яиц и личинок обычно происходит в
воде, причем личинки дышат жабрами;
взрослые животные большей частью живут
на суше, дыша развившимися у них
легкими. У пресмыкающихся, птиц и
млекопитающих вся жизнь проходит на суше
(кроме вторично перешедших к жизни в
воде китов и дельфинов).
Пресмыкающиеся откладывают покрытые защищающей
их от высыхания кожистой скорлупой
яйца, из которых вылупляются уже
сформированные животные. Птицы
откладывают яйца в известковой скорлупе;
большинство птиц заботится о потомстве,
насиживая яйца и выкармливая птенцов.
У млекопитающих все развитие
происходит внутри тела матери, надежно
защищающего зародыш. После рождения
детеныша родители кормят его, защищают и
учат, пока он не подрастет и не станет
самостоятельным.
Скелетная, кровеносная и нервная
системы, а также органы чувств и характер
воспроизводства достигают у позвоночных
животных высших во всем животном мире
форм устройства и дифференцировки,
частично связанных, видимо, с тем, что
многие животные этой группы перешли от
водного к наземному образу жизни.
Позвоночные представляют собой наивысший
этап усложнения строения животных.
Самые древние остатки позвоночных
известны из ордовикских слоев
палеозойской эры.
Позвоночные — самый молодой из
известных типов животных; остатки
других типов животных встречаются в
гораздо более древних слоях. Предками
позвоночных были, по-видимому,
пелагические личинки оболочников, от которых
позвоночные развились неотенически
(путем получения личинками способности
размножения).
Всего насчитывается около 40 тысяч
видов позвоночных, в том числе 20
рыбообразных и рыб, земноводных и
пресмыкающихся — 6, птиц — около 8,6 и
млекопитающих около 4 тысяч видов.
Позвоночные достигли замечательного
разнообразия строения. Среди них есть малютки,
достигающие 1 см длины и 15 мг веса
(филиппинские бычковые рыбы), и
гиганты — 33 м длины и 150 т веса (синие киты).
Позвоночные ведут самый различный
образ жизни, полностью овладев
пространствами нашей планеты, встречаясь
в реках и океане, на поверхности земли и
зарываясь в нее, в нижних слоях
атмосферы. Они приспособились к жизни в
условиях полярных морозов, в сухих
накаленных пустынях, во влажных
тропических лесах и даже в горячих ключах, в
постоянном мраке пещер и на ярком
солнце, при страшном давлении океанских
глубин до 7,8 км и в разреженной атмосфере
высочайших горных вершин. Некоторые
из позвоночных способны улавливать
тончайшие запахи, малейшие изменения
температуры, слабейшие звуки, ничтожные
колебания воды; они одарены необычайной
остротой зрения, позволяющей видеть на
огромном расстоянии.
Некоторые позвоночные обладают
своеобразными звуковыми и
электрическими радарами, ориентируясь с их
помощью в пространстве; у многих из них
есть сложнейшие тепловые, звуковые,
химические, гидродинамические
«приемники» и «антенны». Они достигли
наивысшей для организмов энергетической
эффективности и в особенности
необыкновенного развития нервной системы,
обусловившего достижение наибольшей
высоты психической деятельности.
Рыбы Японского моря и сопредельных частей Охотского и Желтого морей. Часть 1. (Amphioxi — Holocephali). Г.У.Линдберг, М.И.Легеза 1959 г.
Тип Vertebrata — Позвоночные
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА КЛАССОВ РЫБООБРАЗНЫХ И РЫБ
1 (2). Черепа нет. Имеется своеобразная перибранхиальная полость. Кровь бесцветная (Подтип Acrania). I. Amphioxi — Ланцетники.
2 (1). Череп есть. Нет перибранхиальной полости. Кровь красная. (Подтип Craniata).
3 (в). Челюстей нет. Парные плавники отсутствуют. Нет жаберных крышек. 1 непарное носовое (назо-гипофизное) отверстие. Тело червеобразное, голое.
4 (5). Есть спинные плавники (обычно 2, редко 1). Глаза у взрослых нормальные. Ротовой аппарат присасывательного типа. Назо-гипофизная полость не сообщается с глоткой. 7 наружных жаберных отверстий с каждой стороны, расположенных в передней части тела, сразу же позади головы. Рыло без усиков. Носовое отверстие на верхней стороне головы. II. Petromyzones — Миноги.
5 (4). Нет спинного плавника. Глаза дегенерированы, снаружи не различимы; нет ни глазных мышц, ни соответствующих этим мышцам нервов. Ротовой аппарат не присасывательного, а разрушающего типа. Назо-гипофизная полость сообщается с полостью глотки. 1 или от 6 до 15 жаберных отверстий с каждой стороны тела, расположенных на значительном расстоянии от головы. Рыло с усиками. Носовое отверстие на вершине рыла. III. Myxini — Миксины.
6 (3). Есть челюсти. Парные плавники обычно имеются, если же отсутствуют, то внутренние жаберные щели покрыты жаберной крышкой. Носовые отверстия парные.
7 (8). Наружных жаберных отверстий 5—7 с каждой стороны головы; нет жаберной крышки или складки кожи, покрывающей жаберные щели. IV. Elasmobranchii — Акулообразные.
8 (7). Наружных жаберных отверстий только по одному с каждой стороны головы; имеется жаберная крышка или складка кожи, покрывающая жаберные щели.
9 (10). Внутренние жаберные щели (4) покрыты сверху складкой кожи, лишенной костного скелета. У самцов имеются птеригоподии. V. Holocephali — Цельноголовые.
10 (9). Внутренние жаберные щели (4—5) покрыты жаберной крышкой, имеющей специальный костный скелет. Никогда нет птеригоподий. (Обзор Teleostomi будет дан в последующих выпусках.) VI. Teleoslomi — Высшие рыбы.
Подтип Acrania (Cephalochordata, Leptocardii) — Бесчерепные
Хорда продолжается до переднего конца тела, располагаясь здесь впереди центральной нервной системы. Черепа и позвонков нет, парных конечностей и их поясов нет. Хряща в скелете нет. Нет органов слуха и нормальных парных глаз. Головной мозг очень слабо развит. В области головного мозга всего 2 пары нервов. Имеется своеобразная перибранхиальная полость. Органы выделения представлены многочисленными своеобразными нефридиями, расположенными сегментально, без общего выводного протока. Сердце представлено сократимым брюшным сосудом. Кровь бесцветная. Гонады расположены метамерно. Эпидермис из 1 ряда клеток. (Берг, 1940 : 97—98).
Представлен 1 классом — Amphioxi, с 1 отрядом.
I. Класс AMPHIOXI — ЛАНЦЕТНИКИ
Характеристика класса дана при описании подтипа Acrania.
1 отряд (AMFHIOXIFORMES).
Подтип Craniata — Черепные
Хорда продолжается в голову, но доходит самое большее лишь до области infundibulum. У взрослых скелет костный или хрящевой. Есть череп и, как правило, позвонки (кроме Myxini). Обычно есть парные конечности. Есть типичное мускулистое сердце. Кровь красная. Есть парные органы слуха и зрения. Головной мозг хорошо развит. Как правило, не менее 10 пар головных нервов. Перибранхиальной полости нет. Почки не типа нефридий, имеют парные выводные протоки. Гонад не более 1 пары. Эпидермис многослойный. (Берг, 1940 : 98).