(† Ctenacanthiformes Cappetta, 1988) Отряд Ктенакантообразные, Ктенакантиды, Order † Ctenacanthiformes Cappetta, 1988 3 семейства
Синонимы:
Ctenacanthida
Материал подготовил Фактистов И. Е., 2021 г. Сайт www.fishbiosystem.ru
Ктенакантообразные, Ктенакантиды (Ctenacanthiformes) — отряд вымерших хрящевых рыб (Chondrichthyes Huxley, 1880), относящийся к классу пластиножаберных (Elasmobranchii Bonaparte, 1838), похожих на акул. Существовали ктенакантиды 407,6 — 201,3 млн лет назад с начала девона до конца триаса.
Ктенакантообразные — преимущественно хищные рыбы (некоторые — склерофаги с тупыми зубами). Нейрокраний расширен в области орбит и затылка, ростра нет (череп плохо известен у большинства видов). Зубы кладодонтные (с центральной вершиной и симметричными дополнительными зубчиками по бокам) либо пирамидальные; у некоторых — давящие. Зубы закрепляются в челюстях монолитными рядами, сочленяясь друг с другом выступами и впадинами у корня. Парные плавники некрупные, грудной — унисериальный, есть метаптеригиальная ось. Отмечается сходство строения грудного плавника с плавниками современных акул. Два спинных плавника, с крупными шипами. Собственно, по плавниковому шипу группа и была описана, большинство представителей также известно по плавниковым шипам. Плавниковые шипы длинные, слегка изогнутые назад. Длина шипа может достигать 30 см. Поверхность шипов орнаментирована, иногда несёт зубчики, дистальная часть (выступавшая над кожей) покрыта ортодентином (эмалью). Крупные зубчики могут быть на задней поверхности шипа. Передний плавник часто меньше по размеру, его шип более изогнут и направлен назад более полого, радиалий в переднем плавнике может и не быть. В обоих спинных плавниках есть крупные базальные пластинки. Маленький анальный плавник приближен к хвостовому. Хвостовой плавник полулунный, симметричный, верхняя лопасть без радиалий. Чешуя ктенакантного типа.
Ктенакантиды вели разнообразный образ жизни. Виды с типично кладодонтными зубами питались, вероятно, какой-то скользкой добычей (сходная форма зубов у современных песчаных акул). Такой добычей могли быть безраковинные головоногие, конодонты, черви и др. Известны раковины плавающих конулярий, повреждённые зубами кладодонтных рыб. У «Ctenacanthus» costellatus в желудке были остатки мелких рыб. Форма тела выдаёт в ктенакантообразных пелагических хищников. Интересно, что среди них довольно много пресноводных видов. Виды с давящими зубами питались донными и пелагическими моллюсками и ракообразными.
Ктенакантообразные были предками другой важной группы хрящевых рыб — гибодонтообразных. Связь ктенакантообразных с современными акулами не столь очевидна, но самые ранние представители отряда могли быть предками настоящих акул.
Группа ктенакантообразных в целом плохо известна. Типовой род Ктенаканты (Ctenacanthus) был выделен Луи Агассисом в 1835 году на основании крупного плавникового шипа (Ctenacanthus major) из раннего карбона Англии. Не менее 25 видов из девона — перми всех материков были отнесены к этому роду. Кладодонтные зубы, относимые к роду Cladodus, известные из тех же отложений, обычно связывали с ктенакантами, но в последствие выяснилось, что такие зубы характерны для многих палеозойских хрящевых рыб.
Известен отпечаток почти полного скелета небольшой рыбы из раннего карбона Шотландии, обладающей ктенакантовыми шипами («Ctenacanthus» costellatus), но её череп не сохранился. В 1936 году из раннего карбона Шотландии была описана довольно крупная (около 2 метров длиной) рыба гудрихтис (Goodrichthys) с кладодонтным озублением и ктенакантовыми шипами. Впрочем, шипы гудрихтиса глубоко погружены в тело и почти не выступают на поверхность. Тем не менее, их орнамент сходен с орнаментом типового вида ктенакантов. Отпечаток передней части тела ктенакантообразной рыбы известен из девона Огайо (Ctenacanthus clarki). Из поздней перми Германии описаны отпечатки тела некрупного ктенаканта с давящими зубами (Wodnika striatula). Из карбона Небраски известны отпечатки мелкой ктенакантообразной рыбы, относимой к описанному по зубам виду Glikmanius occidentalis.
Классификация отряда не устоялась, по данным сайта Fossilworks выделяют до 3 семейств (не включая устаревшее Tamiobatidae Hay, 1902 и выделенное в самостоятельный отряд Phoebodontidae Williams, 1985) и несколько родов, не относящихся к определённому семейству:
Роды incertae sedis («неопределённого положения» — латинское выражение, указывающее на то, что положение таксона в системе неясно)
• Род Kaibabvenator Hodnett et al., 2012 (1 вид)
• Род Nanoskalme Hodnett et al., 2012 (1 вид)
• Род Neosaivodus Hodnett et al., 2012 (1 вид)
• Род Saivodus Duffin & Ginter, 2006 (1 вид)
Семейство Heslerodidae Maisey, 2010 — небольшие рыбы с тупой мордой и особым строением зубов — боковые зубцы равны центральному, отчего зуб похож на трезубец. Зубы очень мелкие. Челюсти выдаются вперёд по отношению к нейрокранию, очень высокие кзади. Возможно, челюсти могли выдвигаться. Орбиты очень большие. Передний плавник больше заднего, шипы очень глубоко посажены.
• Род Heslerodus Ginter, 2002 (1 вид)
Надсемейство Ctenacanthoidea Zangerl, 1981
Семейство Bandringidae Zangerl, 1969 — очень мелкие (до 20—25 см длиной) рыбы с необычайно длинным уплощённым рылом (внешне напоминают веслоноса или акулу-домового).
• Род Bandringa Zangerl, 1969 — 2 вида, известны из пресноводных отложений позднего карбона Северной Америки.
Семейство Ctenacanthidae Dean, 1909 typus — к семейству относятся практически все представители, известные по отпечаткам скелетов. Десять родов, из девона — триаса всех материков.
• Род Acondylacanthus John & Worthen, 1875 (15 видов)
• Род Amelacanthus Maisey, 1982 (4 вида)
• Род Bythiacanthus St.John & Worthen, 1875 (7 видов)
• Род Cladodus Agassiz, 1843 (49 видов)
• Род Cratoselache Woodward, 1924 (1 вид)
• Род Ctenacanthus Agassiz, 1837 (56 видов)
• Род Eunemacanthus St.John & Worhten, 1883 (4 вида)
• Род Glikmanius Ginter et al., 2005 — 3 вида, род описан по зубам до 43 мм длиной из позднего карбона (Северная Америка и Подмосковье) — средней перми (Северная Америка и Азия). Как уже упоминалось выше, к этому роду принадлежат отпечатки небольших (25 см длиной) рыб из карбона Небраски. Судя по размеру зубов, вид Glikmanius occidentalis из карбона Подмосковья мог достигать более 2 метров в длину.
• Род Glymmatacanthus St.John & Worhten, 1875 (3 вида)
• Род Tamiobatis Eastman, 1897 — 4 вида, известны из позднего девона — раннего карбона северного полушария.
Роды incertae sedis
• Род Asteroptychius Agassiz, 1843 (12 видов)
• Род Pyknotylacanthus Mutter & Rieber, 2005 (2 вида)
• Род Rhombacanthus Williams, 1985 (1 вид)
• Род Wodnika Münster, 1843 (2 вида)
Род (incertae sedis) Кайбабвенаторы — † Kaibabvenator Hodnett, Elliot, Olson & Wittke, 2012 (известен из пермского периода, от 272,5 до 268,0 миллиона лет назад) состоит из одного вида — † Kaibabvenator swiftae Hodnett et al., 2012. [J. P. M. Hodnett, D. K. Elliott, T. J. Olson and J. H. Wittke. 2012. Ctenacanthiform sharks from the Permian Kaibab Formation, northern Arizona. Historical Biology 24:381-395]
† Kaibabvenator swiftae — крупная вымершая ктенакантообразная акула, которая жила в морской среде на территории современной Аризоны в среднепермский период. Kaibabvenator swiftae известна по крупным зазубренным зубам длиной до 30 миллиметров, обнаруженным в Kachina Microsite в нижней части пачки ископаемых гор, в формации Кайбаб (Kaibab Formation) около города Флагстаффа (Flagstaff), штат Аризона, США. Название формации дало названию рода. Видовое название swiftae дано в честь исследовательницы Сандре Свифт (Sandra Swift) за её палеонтологические исследования в Университете Северной Аризоны (Northern Arizona University). Kaibabvenator swiftae, возможно, был высшим морским хищником в древнем море Кайбаб. Оценки его размеров различаются, хотя было высказано предположение, что эта примитивная ктенакантообразная акула могла достигать длины, аналогичной сегодняшней большой белой акуле (Carcharodon carcharias).
Акула Kaibabvenator swiftae, также известная как «Охотник Свифта моря Кайбаб» («Swift's Kaibab hunter»), была большой (около 19,7 футов) акулой с большими зазубренными режущими зубами. Предположительно, это был активный высший хищник по крупной добыче, включая других акул.
Около 260 миллионов лет назад, в пермский период, большая часть Аризоны была покрыта тёплым, относительно мелким морем. Осадки, отложившиеся в этом море, образуют то, что сейчас является формацией Кайбаб, грядой желтоватого известняка, который образует край Гранд-Каньона. Хотя обширная фауна беспозвоночных из Кайбаба уже давно изучается, до недавнего времени очень мало работ было сделано по позвоночным. Первоначальный сбор образцов был произведён местным любителем и выпускником Университета Северной Аризоны Томом Олсоном (Tom Olson) и был расширен студентом университета Ходнеттом (JP Hodnett) и профессором геологии Дэвидом Эллиоттом (David Elliott).
Результатом их работы стала серия статей об акулах Кайбаб, организмах, представленных в основном отдельными зубами и шипами плавников. Первоначальное изучение акул, представленных в коллекциях, предполагает, что здесь может быть представлено до 40 различных таксонов, что делает эту фауну самой разнообразной в своем возрасте в мире. В настоящее время описано девять из этих таксонов, что позволяет идентифицировать ряд акул от больших Kaibabvenator swiftae, которые могли достигать 12–19,7 футов (3,66–5,91 м) в длину и, вероятно, являлись главным хищником в море Кайбаб, до Diablodontus michaeledmundi – небольшие, 2–3-футовые акулы (0,6–0,9 м), похожие на современных куньих акул (hound sharks), которые питаются у дна мелкой рыбой и беспозвоночными.
%201f.jpg) |
%202f.jpg) |
%203f.jpg) |
%204f.jpg) |
%205f.jpg) |
%206f%20teeth.jpg) |
Род (incertae sedis) Наноскальмеи — † Nanoskalme Hodnett, Elliot, Olson & Wittke, 2012 (известен из пермского периода, от 279,5 до 272,5 миллиона лет назад) состоит из одного вида — † Nanoskalme natans Hodnett et al., 2012. [J. P. M. Hodnett, D. K. Elliott, T. J. Olson and J. H. Wittke. 2012. Ctenacanthiform sharks from the Permian Kaibab Formation, northern Arizona. Historical Biology 24:381-395]
Nanoskalme natans входит в список палеозойской жизни Аризоны, который содержит различные доисторические формы жизни, чьи окаменелые останки были зарегистрированы в этом американском штате и имеют возраст от 541 до 252,17 миллиона лет. Окаменелые зубы этой акулы были обнаружены в известняках ископаемых гор формации Кайбаб (Kaibab Formation) около города Флагстаффа (Flagstaff), штат Аризона, США.
Акула Nanoskalme natans, также известная как «лезвие плавающего карлика» («swimming dwarf blade»), имела сильные режущие зубы, похожие на маленькие лезвия, и предназначенные для хватания и резания. Эта акула была маленькой, чуть более трех футов (около 3,2 фута) в длину. Вероятно, это был падальщик и хищник, специализирующийся на мелкой рыбе.
Род (incertae sedis) Неосайводусы — † Neosaivodus Hodnett, Elliot, Olson & Wittke, 2012 (известен из пермского периода, от 272,5 до 268,0 миллиона лет назад) состоит из одного вида — † Neosaivodus flagstaffensis Hodnett et al., 2012. [J. P. M. Hodnett, D. K. Elliott, T. J. Olson and J. H. Wittke. 2012. Ctenacanthiform sharks from the Permian Kaibab Formation, northern Arizona. Historical Biology 24:381-395]
Акула Neosaivodus flagstaffensis, также известная как «новый Saivodus из Флагстаффа» («new Saivodus from Flagstaff», была акулой среднего размера, около 6,6 футов (2 м) с цепкими зубами, которая могла быть в молодости специалистом по поеданию наутилоидов, но во взрослом возрасте — более универсальным хищником. Окаменелые зубы этой акулы были обнаружены в известняках ископаемых гор формации Кайбаб (Kaibab Formation) около города Флагстаффа (Flagstaff), штат Аризона, США.
%201f%20teeth.jpg) |
Род (incertae sedis) Сайводусы — † Saivodus Duffin & Ginter, 2006 (известен из девонского, каменноугольного и пермского периодов, от 360,7 до 272,5 миллиона лет назад) состоит из одного вида — † Saivodus striatus Agassiz 1843. [L. Agassiz. 1843. Recherches Sur Les Poissons Fossiles. Tome III (livr. 15-16). Imprimerie de Petitpierre, Neuchatel 157-390]
Синонимы: Cladodus corrugatus Croneis, 1927; Cladodus curtus Davis, 1883; Cladodus eccentricus St John & Worthen, 1875; Cladodus elongatus Davis, 1883; Cladodus ferox Newberry & Worthen, 1866; Cladodus grandis Newberry & Worthen, 1866; Cladodus hornei Davis, 1883; Cladodus magnificus Tuomey, 1858; Cladodus prototypus Eastman, 1907; Cladodus spinosus Newberry & Worthen, 1866; Cladodus stenopus Newberry & Worthen 1866.
Сайводы жили в то время, когда ктенакантовые акулы были в наибольшем своем разнообразии, ареал обитания которых охватывал весь земной шар. Saivodus striatus — крупная вымершая ктенакантообразная акула, которая была одним из главных хищников поздних морей периода Миссисипи (335—340 миллионов лет назад). Ктенакантовые акулы — дальние родственники современной линии акул, и во время позднего периода Миссисипи они достигли наибольшего разнообразия во всем мире. У этих древних акул были плоские челюсти и большие шипы спинного плавника с уникальным орнаментом по всей длине шипа. Saivodus хорошо известен в летописи окаменелостей отдельных зубов, найденных в Европе и Северной Америке. Некоторые из этих зубов превышали 6 сантиметров в длину от основания зуба, что указывает на то, что эта акула могла достигать размеров тела больше, чем у современных больших белых акул (Carcharodon carcharias), которые могут достигать максимальной длины 20 футов (6 метров).
Saivodus striatus на территории Северной Америки в раннем карбоне обитал в Кентукки, Алабаме и Аризоне.
Самый известный Saivodus был найден на потолке пещеры в Национальном парке Мамонтова пещера (Mammoth Cave National Park), с оценкой длины тела 15—28 футов в длину (4,5—8,4 м). (Мамонтова пещера считается самой длинной в мире — одна только её исследованная часть простирается больше чем на 587 километров.) Вокруг этого экземпляра были обнаружены многочисленные останки. Открытие черепных хрящей, идентифицированных как Saivodus в Национальном парке Мамонтова пещера, является первым в своем роде для этого вида акул. Самый длинный идентифицируемый элемент черепа Saivodus — нижняя челюсть, длина которой составляет около 2,5 футов (75 см). Она выступала из камня, также обнаружены примерно 5 зубов. Большинство последних представляют собой корни зубов, один из которых обнажает коронку и выгибается назад, что позволяет предположить, что это был задний зуб. Эти зубы имеют ширину от 3 до 3,5 сантиметров (1,5 и более дюймов). Левая половина нижней челюсти направлена наружу, это означает, что кусок принадлежит левому элементу. Виден вентральный нижнечелюстной гребень. Этот образец сохраняет элементы хряща. Множество изолированных зубов было собранно из рыхлых отложений со дна пещеры под черепом, и единственный маленький шип, застрявший в отложениях вместе с челюстью, позволяет предположить, что тушу этой акулы поедали несколько более мелких видов акул. Туша Saivodus striatus на морском дне была бы оптимальным источником пищи для других организмов, таких как более мелкие акулы. На рисунке Юлиуса Чотони (см. ниже) изображены животные, которых палеолонтологи обнаружили в этом районе. Ископаемые зубы Bransonella (на переднем плане), Falcatus (в центре, маленькие акулы) и Thrinacodus, напоминающие угря (питающиеся сзади), были обнаружены из рыхлых отложений под черепными хрящами Saivodus в Мамонтовой пещере. Американские учёные считают, что это далеко не последняя находка, ведь когда-то это место буквально кишело хищниками.
Исследователи нашли останки целой стаи акул — 150 особей. По словам палеонтолога Джона-Пола Ходнетта (John-Paul Hodnett), учёные идентифицировали останки до 20 различных видов акул внутри глубокой пещеры, включая голову огромного монстра большого размера, которая частично выступала из стены.
«Мы только поцарапали поверхность, но это уже показывает, что в Мамонтовой пещере есть богатая летопись ископаемых акул» — сказал Ходнетт.
Что интересно, открытие произошло случайно: учёные просто составляли карту Мамонтовой пещеры и наткнулись на нечто, напоминающее акульи зубы. Они отправили фотографию палеонтологу и специалисту по акулам, а он приехал в пещеру. Палеонтолог обнаружил останки головы гигантского монстра, которые выступали из стены. Они были настолько огромными, что учёный даже сначала принял останки за целый скелет. Но оказалось, что это часть челюсти — ещё один сюрприз для исследователя, ведь он нашёл не только зубы, но и хрящ. Это большая редкость, ведь хрящевая ткань мягче костной и обычно не сохраняется, особенно в течение сотен миллионов лет.
По состоянию на 2019 год было идентифицировано не менее 40 различных видов акул и их родственников, в том числе 6 новых видов. Редкая сохранность трёхмерного скелетного хряща, задокументированная в Мамонтовой пещере, позволяет понять анатомию и взаимоотношения этих древних акул.
%201f.jpg) |
%202f.jpg) |
%203f.jpg) |
%204f.jpg) |
%205f.jpg) |
%206f.jpg) |
%207f%20fossil.jpg) |
%208f%20teeth.jpg) |
%209f%20teeth.jpg) |
%2090f%20teeth.jpg) |
%2091f%20teeth.jpg) |
%2092f%20teeth.jpg) |
%2093f%20teeth.jpg) |
Семейство Геслеродиды † Heslerodidae Maisey, 2010, Род Геслеродусы — † Heslerodus Ginter, 2002 (известен из каменноугольного и пермского периодов, от 314,6 до 272,5 миллиона лет назад). [M. Ginter. 2002. Taxonomic notes on “Phoebodus heslerorum” and Symmorium reniforme (Chondrichthyes, Elasmobranchii). Acta Palaeontologica Polonica 47(3):547-555]
Род Геслеродусы († Heslerodus) состоит из одного вида — † Heslerodus divergens Trautschold, 1879. [синонимы — † Phoebodus heslerorum Williams, 1985; † Cladodus divergens Trautschold, 1879]
Таксономический статус Phoebodas heslerorum Williams, 1985. У этого вида есть один или два коротких, частично орнаментированных плавниковых шипа. Из-за этого Уильямс (1985) решил включить свой недавно установленный вид в подсемейство Ctenacanthoidea. Он назвал его Phoebodus (фебодус) из-за предполагаемого сходства зубов между фебодонтами (phoebodont) и Phoebodus heslerorum. В результате этого отнесения к тектенакантоидам (thectenacanthoids) были включены все фебодонты. Однако зубы фебодусов (Phoebodus) значительно отличаются от зубов акулы Уильямса (Вильямса). Таким образом, «Phoebodus heslerorum» может быть ктенакантоидом, но это не фебодонт, и связь между фебодонтами и ктенакантоидами все еще открыта для предположений. Более того, специфический эпитет, heslerorum, по-видимому, является младшим синонимом «Cladodus» divergens Trautschold, 1879. Кладодус, вероятно, является nomen dubium (Zangerl 1973:6; Chorn and Whetstone 1978). [Nomen dubium — «сомнительное название») — означает, что данное название имеет неопределённое применение или по причине невозможности удовлетворительной типификации, или из-за невозможности установить, к какому таксону должен быть отнесён номенклатурный тип данного названия.] Но даже если это действительное имя, зубы Cladodus mirabilis Agassiz, типового вида рода Cladodus, сильно отличаются от зубов акулы Уильямса (личное сообщение К. Даффина, 1998 г.). Таким образом, в любом случае для «Cladodus» divergens необходимо новое родовое имя. Диагноз нового рода Heslerodus, пересмотр записей Heslerodus divergens (= «Phoebodas heslerorum») и описание его зубов представлены ниже.
Род Heslerodus, Ginter 2002; вид Heslerodus divergens (Trautschold, 1879).
Упоминаемые материалы: Зубы, MNA V10441, V10442, V10476, V10493, V10494; три зуба V10489.
Местонахождение и горизонт: MNA LOC 1645, Качина. Нижняя часть пачки ископаемых гор.
Эти маленькие зубы имеют длину от 3 до 4 мм. Срединный бугорок треугольный, суженный. Грубые кристы (гребни) украшают поверхность. Срединный выступ чуть выше базолабиальной впадины и заканчивается на кончике бугорком. Боковые выступы почти равны длине срединного выступа, треугольные и узкие до тонкого. Кристы украшать поверхность боковых бугров, исходя из основание зуба и заканчиваясь на кончике куспид. Кили отсутствуют срединно и латерально. Присутствует единственный небольшой промежуточный бугорок между средним и боковым бугорками. Основание зуба трапециевидной формы с закруглённой лингвальной каймой. Оролингвальный гребень образован двумя близко расположенными закруглёнными дискообразными выступами возле лингвальной дорсальной границы зуба. Многочисленные мелкие отверстия перфорируют заднюю поверхность основания зуба. Базолабиальный гребень формируется
двумя дискообразными выступами, разделёнными глубоким базолабиальным углублением. Основание вентральной поверхности зуба имеет одно большое первичное питательное отверстие с многочисленными дополнительными отверстиями меньшего размера, проникающие в основание зуба (рисунок 3).
Heslerodus divergens известен по остаткам зубов и скелета от верхнего карбона до ранней перми Северной Америки и Евразии (Гинтер, 2002). Пермские зубы Heslerodus divergens известны также из раннепермских отложений в Трехмильном известняке округа Поттаватоми (Threemile Limestone of Pottawatomie County), штат Канзас, США (Schultze 1985, Ginter 2002) и из раннепермских отложений Уральских гор (Иванов 2005) и Печорского моря (Иванов 1999, Гинтер 2002) Российской Арктики. Печорское море — Pechora Sea — акватория в юго-восточной части Баренцева моря, между островами Колгуев и Вайгач. Этот гидроним встречается уже на карте мира Меркатора 1569 года, где относился ко всему Баренцеву морю
%201f%20teeth.jpg) |
%202f%20teeth.jpg) |
%203f%20teeth.jpg) |
 |
 |
Семейство Бандрингиды † Bandringidae Zangerl, 1969, Род Бандринги — † Bandringa Zangerl, 1969 (известен из каменноугольного периода, от 311,45 до 306,95 миллиона лет назад). [R. Zangerl. 1969. Bandringa rayi: A New Ctenacanthoid Shark form the Pennsylvanian Essex Fauna of Illinois. Fieldiana Geology 12:157-169]
Род Бандринги († Bandringa) состоит из двух видов — † Bandringa herdinae Zangerl, 1979 и † Bandringa rayi Zangerl 1969. [R. Zangerl. 1979. New Chondrichthyes from the Mazon Creek fauna (Pennsylvanian) of Illinois. Mazon Creek Fossils 449-500] и [R. Zangerl. 1969. Bandringa rayi: A New Ctenacanthoid Shark form the Pennsylvanian Essex Fauna of Illinois. Fieldiana Geology 12:157-169]
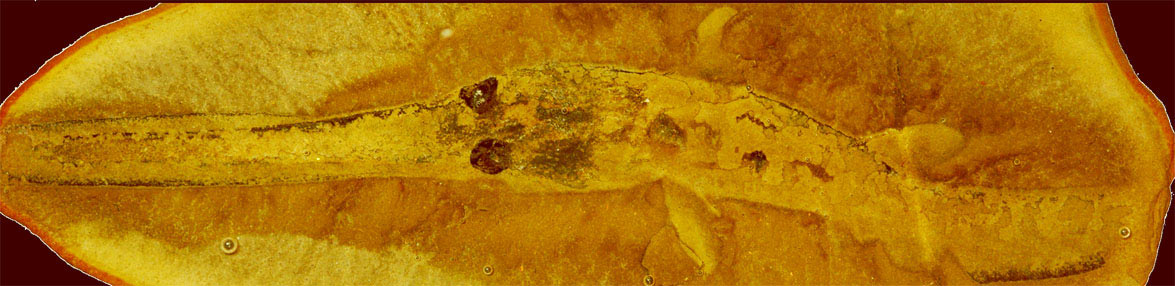
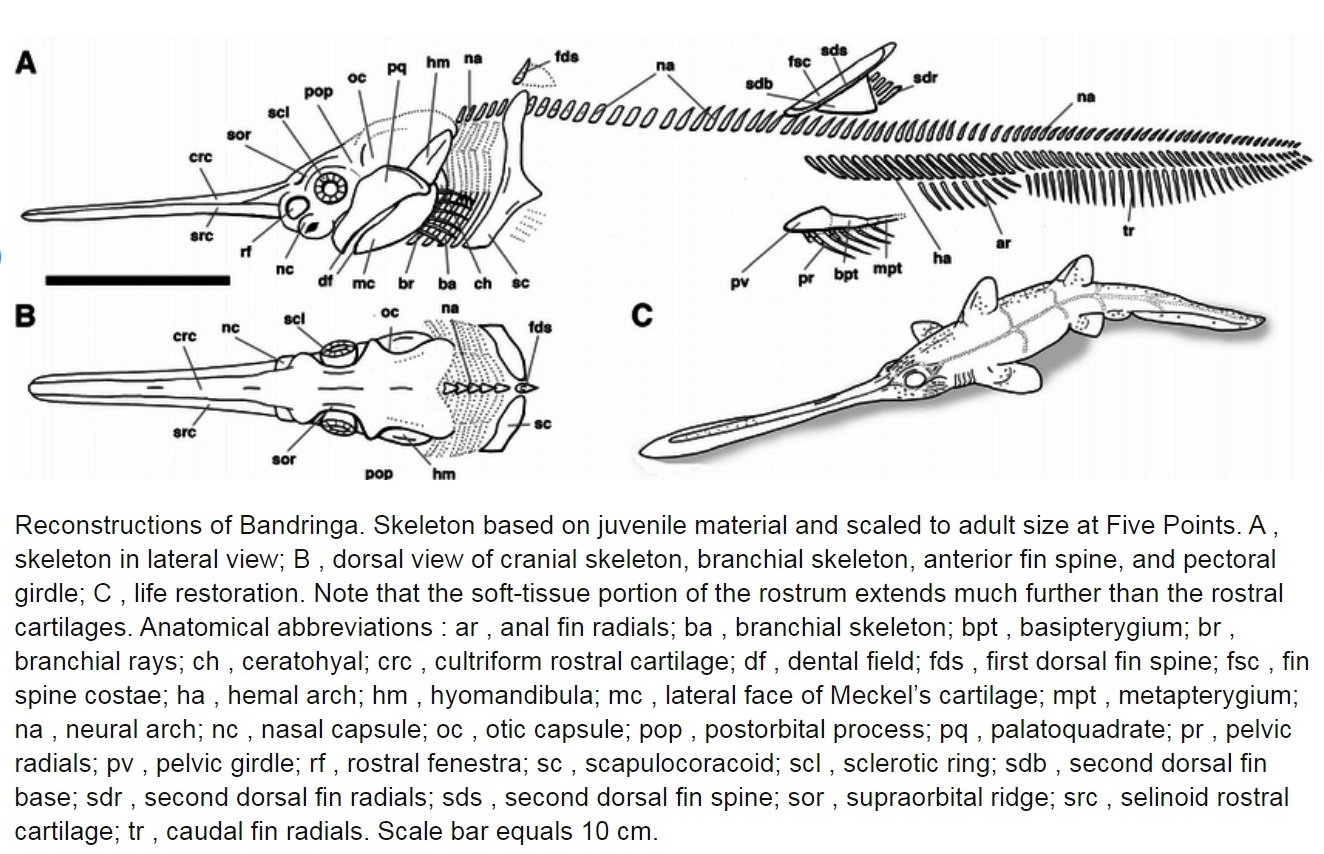
В настоящее время многие систематики выделяют один единственный вид Bandringa rayi, известный из Пенсильванского подпериода карбона. Он известен по особо сохранившимся особям, найденными в Лагерштетте Мазон-Крик (Mazon Creek Lagerstätte) в Иллинойсе (Illinois), который восходит к позднему московскому этапу. Изначально были описаны два вида этого рода, но учёные считают, что различия между ними тафономические по происхождению. (Тафономия — раздел палеонтологии и археологии, изучающий закономерности процессов захоронения и образования местонахождений ископаемых остатков организмов.) Все особи Мазон-Крик, описанные как Bandringa herdinae, похоже, представляют собой молодых особей Bandringa rayi. Это позволяет предположить, что этот район был для них питомником. Такое мнение также подтверждают случаи обнаружения в тех же местах окаменелых яиц, хотя неясно, принадлежат ли они к этому роду. Взрослые окаменелости, приписываемые Bandringa rayi, также были обнаружены в отвалах угольных шахт Five Point около Конесвилля (Conesville), штат Огайо, и Каннелтона (Cannelton), штат Пенсильвания, оба из которых включают ориентировочно формацию Киттанинг (Kittaning Formation) группы Аллегейни (Allegheny Group). Этот вид имеет длинный рострум, который может быть аналогом удлинённых рыл представителей современных семейств Пилоносые акулы, или Акулы-пилоносы (Pristiophoridae) и Пилорылые скаты, или Рыбы-пилы (Pristidae). Похоже, Bandringa rayi питался всасыванием. Законсервированное содержимое кишечника включало конечности членистоногих. По оценкам окаменелостей молодых особей и частично взрослых, размер взрослой особи составляет около 70 см (28 дюймов). Родство Bandringa rayi с другими пластиножаберными рыбами в настоящее время неясно.
По данным палеонтологов из Чикагского и Мичиганского университетов, давно вымершие акулы Bandringa мигрировали вниз по течению от пресноводных болот в океан, чтобы нереститься в мелководных прибрежных водах и оставили после себя ископаемые свидетельства одного из самых ранних известных питомников акул.
Акулы Bandringa — это донные хищники, которые жили в древней системе дельты рек, что сегодня расположены в Верхнем и Среднем Западе. Они напоминали современных рыбу-пилу и веслоноса с рострумом, длинной до половины длины тела. Молодые особи были от 4 до 6 дюймов в длину, а взрослые особи вырастали до 10 футов (3 метра).
Акулы бандринги были обнаружены в 1969 году и вскоре стали одними из самых ценных окаменелостей из хорошо известных месторождений Мазон-Крик на севере Иллинойса. До сих пор палеонтологи считали, что род Bandringa состоит из двух видов — Bandringa rayi и Bandringa herdinae, один из которых обитает в пресноводных болотах и реках, а другой — на морском мелководье. Что выделялось в этих видах, так это то, что при длине до трёх метров типовой вид Bandringa rayi был намного больше, крупнее и обитал в пресной воде. Однако Bandringa herdinae была крошечной акулой, всего от десяти до пятнадцати сантиметров в длину и обитала в солёной воде. Но после переоценки окаменелостей 24 особей доктор Лорен Саллан и доктор Майкл Коутс пришли к выводу, что Bandringa — это единственный вид, который в разное время в течение своей жизни жил в пресной, солоноватой и солёной воде. Это означает, что Bandringa herdinae теперь является синонимом Bandringa rayi, но это также раскрывает очень интересную и поразительную теорию об этой акуле.
«Физические различия между двумя предполагаемыми видами были связаны с разными процессами сохранения в морских и пресноводных местах. Пресноводные участки, как правило, сохраняли кости и хрящи, в то время как морские участки сохраняли мягкие ткани», — пояснили д-р Саллан и д-р Коутс, соавторы статьи, опубликованной в Journal of Vertebrate Paleontology.
Объединив дополнительные наборы данных из обоих типов окаменелостей и реклассифицируя Bandringa как единый вид, команда получила гораздо более полную картину анатомии вымершей акулы и обнаружила несколько ранее неизвестных особенностей. Они включают в себя направленные вниз челюсти, идеально подходящие для всасывающего кормления со дна, игольчатые шипы на голове и щеках, а также сложный набор органов чувств как на вытянутой морде, так и на теле, подходящих для обнаружения добычи в мутной воде. Окаменелости Bandringa также показывают единственный известный пример миграции акул из пресной воды в солёные воды, а также 310 млн-летний акулий питомник, где в отложениях имели место случаи обнаружения фоссилизируемых яиц и сохранившихся малолетних акул.
«Взрослые акулы Bandringa жили исключительно в пресноводных болотах и реках», — сказали ученые. «Самки, очевидно, спускались вниз по течению к тропическому побережью, чтобы отложить яйца в мелководных морских водах, что является обратной версией миграции современного лосося из моря в реки и ручьи. В то время береговая линия суперконтинента Пангеи проходила по диагонали между пресноводными и морскими участками Мазон-Крик».
Но почему Бандринги решили основать питомники в соленой воде? Пока молодые особи Бандринги жили в неглубоких прибрежных водах, это давало им ряд преимуществ. Мелкие прибрежные воды обычно имеют пропорционально широкое изобилие жизни с почти неограниченным количеством мелких водных животных, от червей до креветок, и даже некоторые виды рыб закапываются в мягких песчаных отложениях. Большое количество мелких животных — это именно то, что нужно молодым акулам, чтобы быстро вырасти. Прибрежные воды также ограничивают передвижения более крупных хищников, давая больше шансов на выживание для молоди Бандринги. Эти воды также имеют тенденцию быть намного теплее, их легче нагревает солнце и, возможно, даже океанические течения, что повышает скорость метаболизма и ускоряет рост молодых акул по сравнению с более холодными реками.
Те же самые мягкие отложения, которые содержали большое количество кормовых животных для молоди Bandringa, также обеспечивали высокий уровень сохранности некоторых из этих молодых экземпляров с сохранением даже мягких тканей. Это помогло подтвердить наличие некоторых особенностей у этих акул, таких как обращённый вниз рот, а также обнаружение новых, не сохранившихся у взрослых особей из-за различных факторов окаменения, не сохраняющих их. К ним относятся небольшие игольчатые шипы на голове и щеках, а также наличие обширного массива электрорецепторов на морде (рострум). Это подтверждает то, что морда Бандринги (как и у других акул с длинной мордой) в первую очередь служила сенсорным органом, обнаруживающим закопанную добычу, а затем использовалась для рытья при её извлечении.
Мы теперь знаем, что один род акул мигрировал из пресноводных вод в солёную, чтобы вырастить детенышей в питомнике, и возникает следующий вопрос, сколько других видов «пресноводных» акул мигрировало в места размножения с солёной водой?
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
Семейство Ктенакантиды † Ctenacanthidae Dean, 1909, Род Кондилакантусы, Акондилакантусы — † Acondylacanthus John & Worthen, 1875 (известен из девонского, каменноугольного и пермского периодов, от 360,7 до 254,0 миллиона лет назад) состоит из следующих видов: Acondylacanthus aequicostatus Saint John & Worthen, 1875; Acondylacanthus attenuatus Davis, 1883; Acondylacanthus browni Branson, 1916; Acondylacanthus colei Davis, 1883; Acondylacanthus distans M'Coy, 1848; типовой вид — Acondylacanthus gracilis Saint John & Worthen, 1875; Acondylacanthus gracillimus Newberry & Worthen, 1866; Acondylacanthus jaekeli (Gross, 1933); Acondylacanthus junceus (McCoy, 1848); Acondylacanthus mudgianus Saint John & Worthen, 1883; Acondylacanthus nuperus St John & Worthen, 1883; Acondylacanthus rectus Saint John & Worthen, 1883; Acondylacanthus remotus (Eichwald, 1861); Acondylacanthus tenuistriatus Davis, 1883; Acondylacanthus tuberculatus Davis, 1883.
Большинство видов известны из каменноугольного периода Америки и с Британских островов. Acondylacanthus jaekeli известен из верхнего Франского (Frasnian, от 382,7 миллиона лет назад до 372,2 миллиона лет назад) фаунистического этапа Германии и верхнего Фаменского (Famennian, от 372,2 миллиона лет назад до 358,9 миллиона лет назад) фаунистического этапа позднего девона России. Этот род полностью основан на описании отдельных шипов спинного плавника и, таким образом, позже может быть найден род-синоним, описанный на основе зубов. Многие виды были отнесены к этому роду, хотя есть некоторая путаница с родом Ctenachanthus. Этот род ещё нуждается в обзоре и исследованиях.
%201f%20fossil.jpg) |
%201f%20fossil.jpg) |
%201f%20fossil.jpg) |
%202f%20fossil.jpg) |
%201f%20fossil.jpg) |
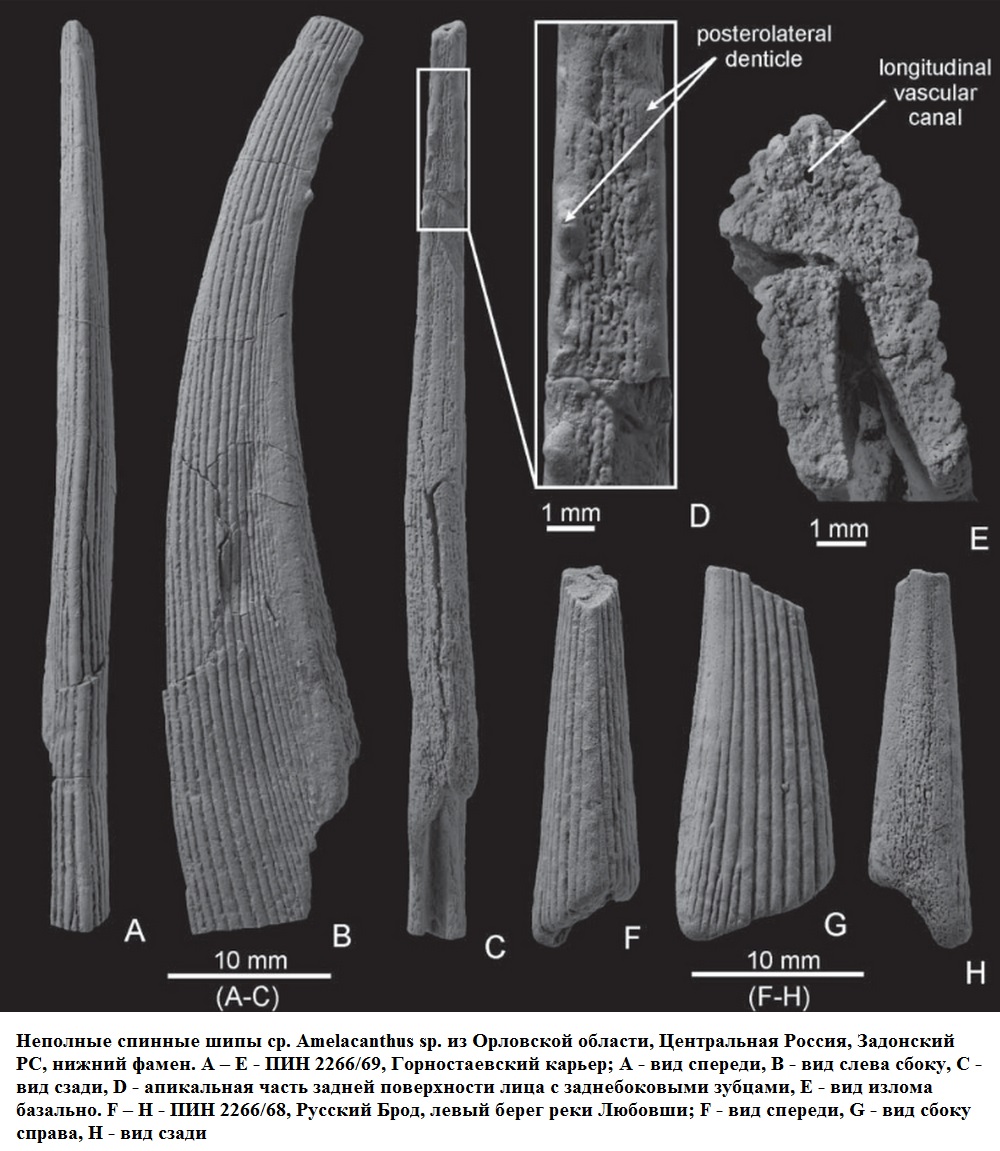
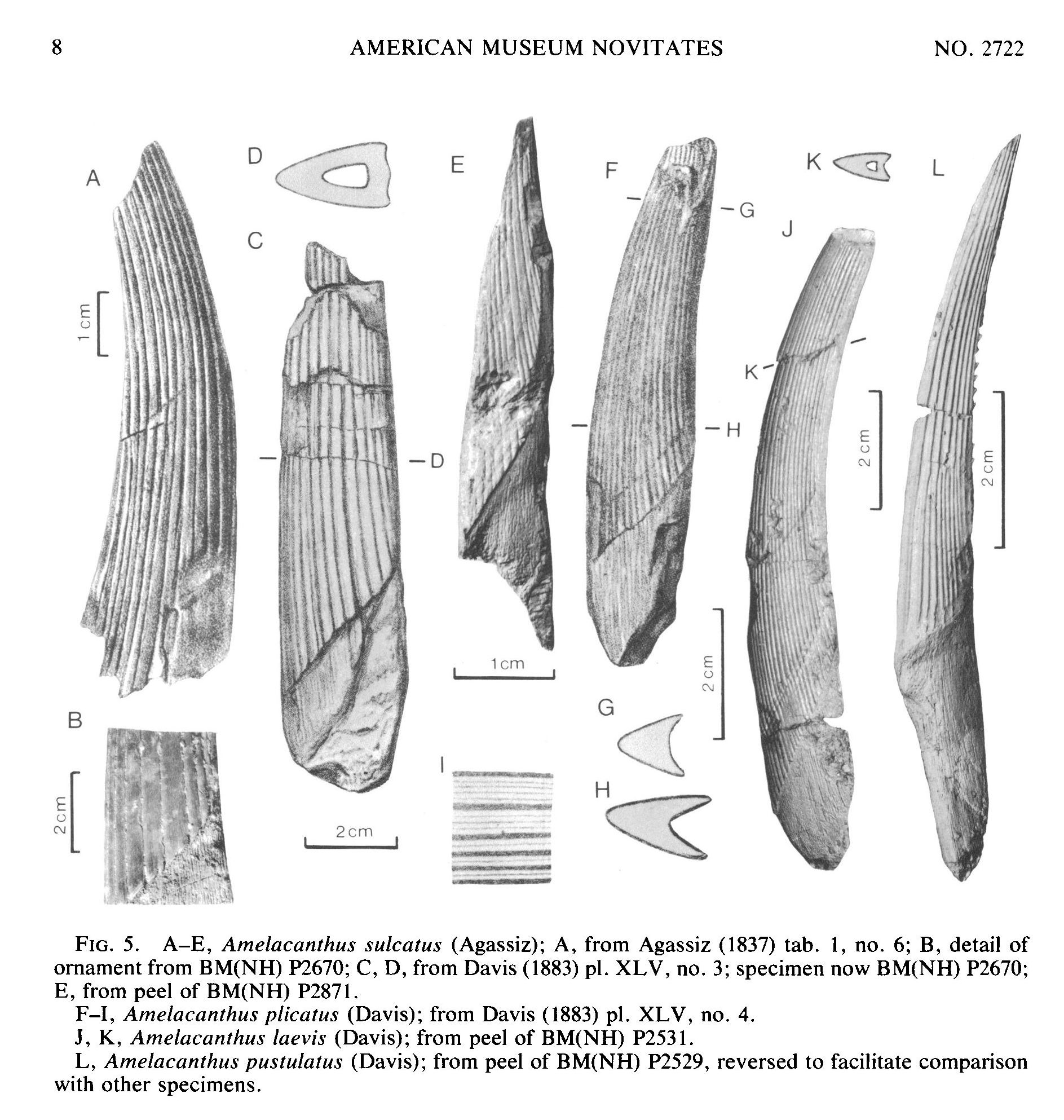
Семейство Ктенакантиды † Ctenacanthidae Dean, 1909, Род Амелакантусы — † Amelacanthus Maisey, 1982 (известен из девонского, каменноугольного, пермского и триасового периодов, от 360,7 до 247,2 миллиона лет назад) состоит из следующих видов: Amelacanthus laevis Davis, 1883; Amelacanthus plicatus Davis, 1883; Amelacanthus pustulatus Davis, 1883; типовой вид — Amelacanthus sulcatus Agassiz, 1837 [синоним — Onchus sulcatus Agassiz, 1837]. [J. G. Maisey. 1982. Studies on the Paleozoic Selachian Genus Ctenacanthus Agassiz: No. 2. Bythiacanthus St. John and Worthen, Amelacanthus, New Genus, Eunemacanthus St. John and Worthen, Sphenacanthus Agassiz, and Wodnika Munster. American Museum Novitates 2722:1-24]
Род известен по шипам плавников, и в настоящее время насчитывает четыре описанных вида. Большинство видов известны из перми и карбона в Северной Америке, Европе и Африке. Известны также из Фаменского (Famennian, от 372,2 миллиона лет назад до 358,9 миллиона лет назад) фаунистического этапа позднего девона в России.
 |
 |
 |
Семейство Ктенакантиды † Ctenacanthidae Dean, 1909, Род Битиакантусы — † Bythiacanthus St.John & Worthen, 1875 (известен из каменноугольного периода, от 306,95 до 303,4 миллиона лет назад) состоит из следующих видов: Bythiacanthus brevis Agassiz, 1837; Bythiacanthus ianishevskyi Khabakob, 1926; Bythiacanthus lopesi Figueroa & Gallo, 2017; Bythiacanthus lucasi Eastman, 1902; Bythiacanthus siderius Leidy, 1870; Bythiacanthus solidus Eastman, 1903; типовой вид — Bythiacanthus vanhornei St.John & Worthen, 1875.
Род известен по шипам спинных плавников, ромбовидной формы, которые несут продольные ряды крупных бугорков с радиальными бороздками, подобными тем, что наблюдаются у рода Heslerodus.
%201f.jpg) |
%201f%20fossil.jpg) |
%202f%20fossil.jpg) |
Семейство Ктенакантиды † Ctenacanthidae Dean, 1909, Род Кладодусы — † Cladodus Agassiz, 1843 (известен из каменноугольного, пермского и триасового периодов, от 345,0 до 221,5 миллиона лет назад). [L. Agassiz. 1843. Recherches Sur Les Poissons Fossiles. Tome III (livr. 15-16). Imprimérie de Petitpierre, Neuchatel 157-390]
Род Кладодусы — † Cladodus Agassiz, 1843 состоит из следующих видов:
• † Cladodus acuminatus Newberry, 1856
• † Cladodus alternatus St. John & Worthen, 1875
• † Cladodus angulatus Newberry & Worthen, 1866
• † Cladodus bellifer St. John & Worthen, 1875
• † Cladodus claypolei Hay, 1899
• † Cladodus concinnus Newberry, 1875
• † Cladodus conicus Agassiz, 1843
• † Cladodus coniger Hay, 1899
• † Cladodus costatus Newberry & Worthen, 1866
• † Cladodus curvus Davis, 1883
• † Cladodus deflexus Newberry & Worthen, 1870
• † Cladodus divaricatus Trautschold, 1874
• † Cladodus elegans Newberry & Worthen, 1870
• † Cladodus eriensis Bryant, 1935
• † Cladodus euglypheus Saint John & Worthen, 1875
• † Cladodus exiguus Saint John & Worthen, 1875
• † Cladodus exilis Saint John & Worthen, 1875
• † Cladodus formosus Hay, 1902
• † Cladodus fulleri Saint John & Worthen, 1875
• † Cladodus gomphoides Saint John & Worthen, 1875
• † Cladodus gracilis Newberry & Worthen, 1866
• † Cladodus hertzeri Newberry, 1875
• † Cladodus ischypus Newberry & Worthen, 1870
• † Cladodus keokuk Saint John & Worthen, 1875
• † Cladodus knightianus Cope, 1891
• † Cladodus lamnoides Newberry & Worthen, 1866
• † Cladodus marginatus Agassiz, 1843
• † Cladodus micropus Newberry & Worthen, 1866)
• † Cladodus milleri Agassiz, 1843
• † Cladodus mirabilis Agassiz, 1843 (типовой вид — type species)
• † Cladodus monroei Eastman, 1900
• † Cladodus newmani Tuomey, 1858
• † Cladodus pandatus St. John & Worthen, 1875
• † Cladodus parvulus Newberry, 1875
• † Cladodus parvus Agassiz, 1843
• † Cladodus pattersoni Newberry, 1875
• † Cladodus politus Newberry & Worthen, 1866
• † Cladodus raricostatus Saint John & Worthen, 1875
• † Cladodus rivipetrosi Claypole, 1893
• † Cladodus robustus Newberry & Worthen, 1866
• † Cladodus romingeri Newberry, 1875
• † Cladodus springeri St. John & Worthen, 1875
• † Cladodus subulatus Newberry, 1875
• † Cladodus terrelli Newberry, 1899
• † Cladodus tumidus Newberry, 1899
• † Cladodus turritus Newberry & Worthen, 1866
• † Cladodus vanhornei St. John & Worthen, 1875
• † Cladodus yunnanensis Pan, 1964
• † Cladodus zygopus Newberry & Worthen, 1866
Кладодусы († Cladodus) — род вымерших хрящевых рыб, предположительно отличающихся зубами с высоким центральным бугорком, широким основанием и одной или несколькими парами боковых бугорков, признак, который, как теперь известно, широко встречался у ранних Пластиножаберные. Жили Кладодусы от карбона до перми и имели всемирное распространение. Их зубы были с множеством бугорков и гладкими краями, что делало их пригодными для хватания, но не для разрывания или жевания жертвы. Окаменелости Cladodus были найдены в Баркипе, Шотландия и в формации Питкин (каменноугольный период) в Арканзасе, США. Кроме того, окаменелости, относящиеся к Cladodus, известны из сланцев каньона Мэннинг каменноугольного периода в штате Юта.
Некоторые систематики относят род Cladodus к семейству Кладоселаховые, Кладоселяховые — † Cladoselachidae Dean, 1894, монотипического отряда Кладоселахообразные, Кладоселяхообразные († Cladoselachiformes Dean, 1909).
Кладодус беллифер — † Cladodus bellifer St. John & Worthen, 1875
%201p.jpg) |
%202f%20fossil.jpg) |
Кладодус маргинатус — † Cladodus marginatus Agassiz, 1843
%201f%20fossil.jpg) |
%202f%20fossil.jpg) |
%203f%20fossil.jpg) |
%204f%20fossil.jpg) |
Род Кладодусы — † Cladodus Agassiz, 1843
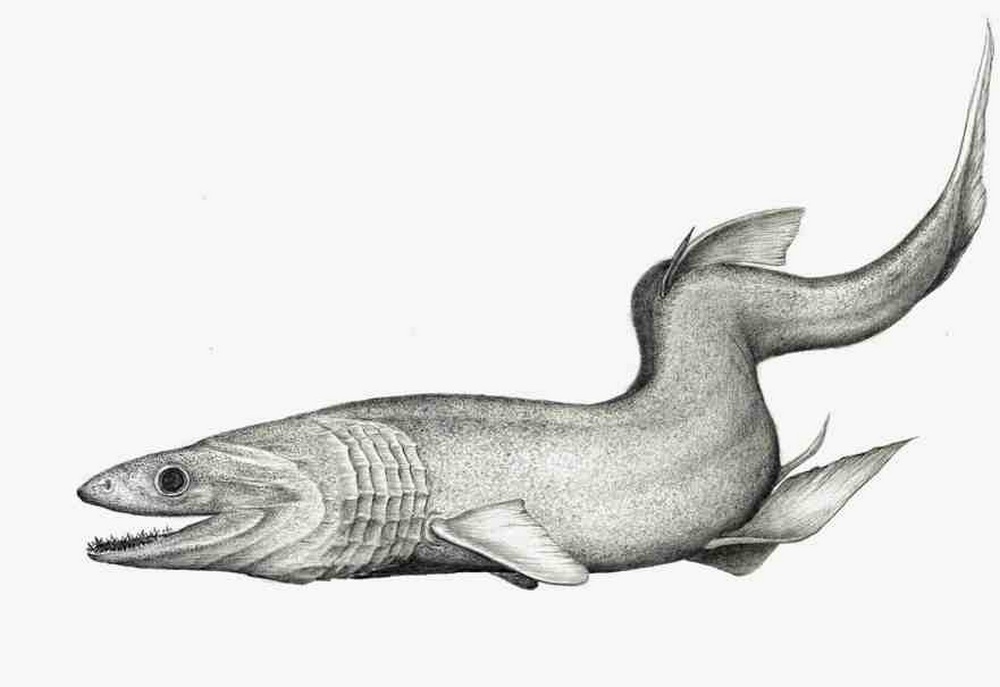 |
 |
 |
 |
 |
 |
Семейство Ктенакантиды † Ctenacanthidae Dean, 1909, Род Кратоселахии — † Cratoselache Woodward, 1924 (известен из каменноугольного периода, от 345,0 до 340,0 миллиона лет назад) состоит из одного вида — † Cratoselache pruvosti (Woodward & Legraye, 1924). [Arthur Smith Woodward & Michel Legraye, 1924. «Un nouvel Elasmobranche (Cratoselache pruvosti gen. et sp. nov.) du calcaire carbonifère inférieur de Denée», Annales de la Société géologique de Belgique [En ligne], Mémoires in-4°, Tome 5: livre jubilaire du 50e anniversaire de la Société, 1874 — 1924, Volume 1 — fascicule 1, 57 — 62]
Ископаемые останки Cratoselache pruvosti были найдены в Европе. Эта рыба должна быть около полутора метров в длину и напоминать по общему виду архаичную «акулу» † Cladoselache из девона. Кратоселахии обладали многочисленными характеристиками, типичными для пластиножаберных (Elasmobranchii Bonaparte, 1838), например, строение грудного пояса, но характеризовались наличием костных или кальцинированных пластин, которые, по-видимому, являлись частью верхней части черепа. Эти бляшки были описаны, как не похожие на дермальные бляшки. Грудной пояс Cratoselache был очень похож на грудной пояс Cladoselache.
Впервые описанный в 1924 году Артуром Смитом Вудвордом (Arthur Smith Woodward) и Мишелем Легрей (Michel Legraye), Cratoselache pruvosti известен по образцу, обнаруженному в почвах нижнего карбона вблизи деревни Дене (Denée), Бельгия. Эти окаменелости из-за странных бляшек на голове были отнесены учёными к различным плакодермам (или пластинокожим рыбам, устаревшее название — панцирные рыбы — класс челюстноротых (Gnathostomata), живших в палеозое, начиная с раннего девона). В 1963 году Стенсио (Stensio) основал группу Кратоселахии (Cratoselachi) и правильно отнёс это животное к пластиножаберным (Elasmobranchii Bonaparte, 1838). Совсем недавно в 1981 году Райнер Зангерл (Rainer Zangerl) указал, что род Кратоселахии — † Cratoselache, должно быть, отклоняющийся от нормы представитель отряда ктенакантообразных.
Семейство Ктенакантиды † Ctenacanthidae Dean, 1909, Род Ктенакантусы — † Ctenacanthus Agassiz, 1837 (известен из каменноугольного, пермского и триасового периодов, от 345,0 до 221,5 миллиона лет назад). [L. Agassiz. 1837. Recherches Sur Les Poissons Fossiles. Tome III (livr. 8-9). Imprimérie de Petitpierre, Neuchatel viii-72; L. Agassiz. 1843. Recherches Sur Les Poissons Fossiles. Tome III (livr. 15-16). Imprimérie de Petitpierre, Neuchatel 157-390]
L. Agassiz. 1837. Recherches Sur Les Poissons Fossiles. Tome III (livr. 8-9). Imprimerie de Petitpierre, Neuchatel viii-72
Род Ктенакантусы — † Ctenacanthus Agassiz, 1837 состоит из следующих видов:
• † Ctenacanthus abnormis Giebel, 1858;
• † Ctenacanthus acutus Eastman, 1897;
• † Ctenacanthus acutus Eastman, 1897;
• † Ctenacanthus amblyxiphias Cope, 1891
• † Ctenacanthus angulatus Newberry & Worthen, 1866;
• † Ctenacanthus angustus Newberry, 1889;
• † Ctenacanthus burlingtonensis St.John & Worthen, 1875;
• † Ctenacanthus buttersi St. John & Worthen, 1883
• † Ctenacanthus chemungensis Claypole, 1885
• † Ctenacanthus cliftonensis Branson & Mehl, 1938
• † Ctenacanthus clarkii Newberry, 1889;
• † Ctenacanthus compressus Newberry, 1889;
• † Ctenacanthus concinnus Newberry, 1875
• † Ctenacanthus costatus Newberry & Worthen, 1866;
• † Ctenacanthus costellatus Traquair, 1884
• † Ctenacanthus coxianus St.John & Worthen, 1883;
• † Ctenacanthus crenatus McCoy, 1855;
• † Ctenacanthus crenulatus Agassiz, 1837;
• † Ctenacanthus cylindricus Newberry, 1889;
• † Ctenacanthus decussatus Eastman, 1902;
• † Ctenacanthus denticulatus McCoy, 1855;
• † Ctenacanthus depressus Newberry, 1897;
• † Ctenacanthus elegans Tuomey, 1858 — из Алабамы (Alabama);
• † Ctenacanthus excavatus Saint John & Worthen, 1875;
• † Ctenacanthus fallax Woodward, 1891;
• † Ctenacanthus formosus Newberry, 1873;
• † Ctenacanthus furcicarinatus Newberry, 1875
• † Ctenacanthus gemmatus Saint John & Worthen, 1875;
• † Ctenacanthus gracillimus Newberry & Worthen, 1866;
• † Ctenacanthus gradocostatus Saint John & Worthen, 1875;
• † Ctenacanthus gurleyi Newberry, 1897;
• † Ctenacanthus keokuk Saint John & Worthen, 1875;
• † Ctenacanthus laevis Davis, 1883;
• † Ctenacanthus littoni Newberry, 1889;
• † Ctenacanthus major Agassiz, 1843 (типовой вид — type species)
• † Ctenacanthus maranhensis Santos, 1946
• † Ctenacanthus marshii Newberry, 1873;
• † Ctenacanthus mayi Newberry & Worthen, 1870;
• † Ctenacanthus minor Davis, 1879;
• † Ctenacanthus mutabilis Branson, 1933;
• † Ctenacanthus nodocostatus Hussakof & Bryant, 1918
• † Ctenacanthus obscuracostatus Branson, 1916;
• † Ctenacanthus pellensis Saint John & Worthen, 1883;
• † Ctenacanthus pugiunculus Saint John & Worthen, 1875;
• † Ctenacanthus pustulatus Davis, 1883;
• † Ctenacanthus randalli Newberry, 1889;
• † Ctenacanthus sculptus Saint John & Worthen, 1875;
• † Ctenacanthus similis Saint John & Worthen, 1875;
• † Ctenacanthus speciosus Saint John & Worthen, 1875;
• † Ctenacanthus terrelli Newberry, 1889
• † Ctenacanthus triangularis Newberry, 1873;
• † Ctenacanthus tumidus Newberry, 1889
• † Ctenacanthus venustus Eastman, 1902;
• † Ctenacanthus vetustus Newberry, 1873;
• † Ctenacanthus wrighti Newberry, 1884;
• † Ctenacanthus xiphias Saint John & Worthen, 1883;
Виды переведённые в синонимию (species brought into synonymy):
• † Ctenacanthus brevis Agassiz, 1837 syn. of Avonacanthus Maisey 2010.
Ктенакантусы жили около 380–258 миллионов лет назад, а их ископаемые останки были найдены в Европе, Северной Америке, Южной Америке, Азии и Гренландии.
Эти вымершие акулы в основном известны отдельными шипами спинных плавников, но есть и более полные экземпляры, которые позволили учёным реконструировать их внешний вид, по крайней мере, ориентировочно. В частности, вид Ctenacanthus compressus также известен по двум неполным экземплярам, состоящим из двух передних половин скелета; голова имела короткую морду, с острыми зубцами и множеством бугорков (кладодонтового типа). Scapularcoracoid, который поддерживал грудные плавники, имел характерную вогнутую форму спереди и заострённую дистально. Оба спинных плавника поддерживались большими шипами круглого сечения, глубоко вставленными в тело животного и опирающимися на хрящ более или менее треугольной формы. Колючки были украшены рядами бугорков, которые делали их похожими на гребни (название Ctenacanthus означает «гребешок с шипами»), и были покрыты веществом, похожим на зубную эмаль.
Классификация рода Ctenacanthus чрезвычайно сложна, в основном из-за того, что подавляющее большинство видов, относимых к этому роду, основаны на изолированных шипах и зубах. Типовой экземпляр был описан Агассисом в 1837 году по шипу спинного плавника, происходящим из нижнего карбона ущелья Эйвон вблизи Бристоля, Англия. Швейцарский учёный отнёс к этому роду другие виды, в том числе Ctenacanthus brevis (шип, которого учёный никогда не видел, а описал только на основе рисунка), Ctenacanthus ornatus (вероятно, принадлежащий к другой группе акантусовых рыб) и Ctenacanthus tenuirostratus (у которого, по крайней мере, часть материала не сохранилась). В последующие годы к этому роду были отнесены многие другие виды, происходящие из многих частей мира и из геологического горизонта между средним девоном и верхней перми, более 120 миллионов лет.
Таксономия рода Ctenacanthus осложнилась благодаря одному неудачному палеонтологическому случаю: в 1873 году, палеонтолог Джон Стронг Ньюберри (John Strong Newberry) предпринял повторную попытку исследования американских образцов Ctenacanthus, а так же были материалы для сравнения, присланные из Шотландии. К сожалению, два шипа по ошибке присланые из Шотландии, принадлежали представителям рода Sphenacanthus, вместо ожидаемых шипов типового вида Ctenacanthus major. Эта ошибка чревата последствиями: Ньюберри, полагая, что он исследовал шипы Ктенаканта, подверг критике первоначальное описание Агассиса и изменил его; все это привело к целому столетию смешения родов Ctenacanthus и Sphenacanthus, которая была разрешена только между 1981 и 1984 годами, когда Джон Мейси (John Maisey) из Нью-Йоркского музея естественной истории (American Museum of Natural History) предпринял пересмотр рода Ctenacanthus. Мейси показал, что из примерно сотни видов допустимыми считаются только пятнадцать: помимо типового вида, по крайней мере, Ctenacanthus amblyxiphias, Ctenacanthus buttersi, Ctenacanthus chemungensis, Ctenacanthus cliftonensis, Ctenacanthus compressus, Ctenacanthus concinnus признаны действительными, Ctenacanthus elegans, Ctenacanthus furcicarinatus, Ctenacanthus major, Ctenacanthus maranhensis, Ctenacanthus nodocostatus, Ctenacanthus terrelli, Ctenacanthus tumidus. Многие другие виды были отнесены к другим родам палеозойских акул: помимо Sphenacanthus, также Hopleacanthus, Bythiacanthus, Amelacanthus, Avonacanthus и Eunemacanthus. Лучшие окаменелости Ctenacanthus происходят из Кливлендских сланцев в Огайо и относятся к виду Ctenacanthus compressus.
Ctenacanthus должен был быть активным хищником, о чём свидетельствует форма тонкого тела и острые зубы с несколькими остриями; в любом случае, шипы на спине выполняли защитную функцию от более крупных животных, таких как хищные плакодермы: был обнаружен образец плакодермы Holdenius с шипом ктенаканта, застрявшим в нёбе.
Шипы спинных плавников акул Ctenacanthus находят в морских отложениях от девона до перми. Как показано выше, у ктенакантид есть пара жестких шипов плавников, которые выступают перед каждым из двух спинных плавников. Название Ctenacanthus буквально означает «гребенчатый шип» и происходит от характерного гребенчатого вида двух шипов плавников. Шипы спинного плавника украшены тонкими рядами материала, похожего на дентин, который в некоторых местах выглядит как нити крошечных бусинок. Шипы ктенакантид были субцилиндрическими и заострёнными и при жизни поддерживались клином из хряща, который был вставлен в бороздку на задней стороне ости плавника.
Ктенакантус Agassiz 1835 входит в отряд Euselachii Hay (1902). Согласно Зангерлу (1981, стр. 51), «палеозойские представители этой группы легко идентифицируются как имеющие два спинных плавника, каждому из которых предшествует шип, апикальный конец которого выходит за пределы эпидермиса и заканчивается внешним ортодентиновым слоем, который также образует орнамент (Maisey, 1975). Каждый шип поддерживается треугольным хрящевым участком, который входит в его центральную полость сзади. Радиальные части парных плавников и брюшная доля хвостового плавника обычно не разделены поперечно, есть небольшая разница в размере и прочности передних и задних лучевых костей. Имеется один анальный плавник. Форма тела веретеновидная.»
Первое сообщение о палеозойской акуле Ctenacanthus amblyxiphias Cope 1891 из нижней перми в графстве Моррис, штат Канзас. В августе 2002 г. в северо-восточной части были обнаружены останки небольшой (1,6 м) палеозойской акулы (FHSM VP-15012), подвергшейся эрозии при обнажении пачки Грант формации Винфилд (группа Чейз, серия Вольфкэмпиан, нижняя пермь) округа Моррис, штат Канзас. Матрица, окружающая образец, представляет собой мелкозернистую кальцитовую глинистую породу, которая интерпретируется как прибрежная мелководная залежь. Остатки включают передний и задний шипы спинного плавника, несколько кусочков кальцинированного хряща, включая левый скапуло-коракоид, чешуйки плакоида и ганоида, а также зубы плакодонта. Длина шипов переднего и заднего плавников почти целых 25 и 18 см. Шипы плавников отчетливо украшены тонкими рядами дентиноподобного материала. Останки были идентифицированы как Ctenacanthus amblyxiphias в сравнении с шипом спинного плавника, описанным Копом из Красных холмов (Пермь) в Техасе. С останками были связаны обильные раковины брахиопод Composita subtilata, а также окремнённые иглы, мшанки и фрагменты лилий. Чашечка лилий, условно идентифицированная как Delocrinus brownvillensis , была обнаружена поблизости на том же стратиграфическом уровне. Обзор литературы и музейных коллекций показывает, что это первое сообщение об этом виде акул из Канзаса.
Останки ктенакантусов были найдены в сланцах формации Блойд (Bloyd Formation) в Арканзасе, США (каменноугольный период), в Южной Америке и в Шотландии. Род Ctenacanthus представлен множеством видов, почти все они описаны на шипах плавников. Шипы спинного плавника акул Ctenacanthus находят в морских отложениях от девона до перми.
Типовой образец, Ctenacanthus major, представляет собой шип из нижнего карбона Эйвонского ущелья близ Бристоля в Англии. Луи Агассис относил к роду Ctenacanthus ряд других материалов. К ним, в частности, относятся Ctenacanthus brevis (шип, который он никогда не видел и который он описал на основе рисунка, сделанного кем-то другим), Ctenacanthus ornatus (вероятно, акантод) и Ctenacanthus tenuistriatus (по крайней мере, часть которого также исчезла). В последующие годы к роду Ctenacanthus был отнесен ряд других экземпляров, в том числе многие шипы с вогнутыми задними поверхностями или широкими эмалированными полосами, а также материалы, которые не имели никакого отношения к этому типу, такие как русские головоногие моллюски.
Однако в 1873 году палеонтолог Ньюберри (Newberry) пересмотрел диагноз Агассиса, он раздобыл два экземпляра Ctenacanthus major для личного осмотра и изучения. К сожалению, по некоторому нелепому совпадению не только оба экземпляра были неправильно идентифицированы, но и оба оказались (гораздо позже) вовсе не ктенакантами. Вместо этого, похоже, что они оба были останками Сфенакантуса (Sphenacanthus). Этот род также был описан Агассисом, но материал голотипа был настолько скудным, что Ньюберри, по-видимому, не распознал сходство. Шипы Sphenacanthus весьма непохожи на шипы Ctenacanthus и «имеют неправильные, широко расставленные ребра, иногда с разбросанными бугорками, но никогда не выступающими, как у Ctenacanthus major, а сам шип вогнутый или плоский сзади».
Учитывая такие противоречивые данные, неудивительно, что Ctenacanthus стал почти бессмысленным таксоном. В последующие десятилетия большое количество кусков и кусочков окаменелостей было отнесено к роду Ctenacanthus. Было даже предложено, чтобы Ctenacanthus и Hybodus были синонимами. Так могло продолжаться бесконечно долго, если бы не обнаружение и постепенная корреляция окаменелостей из Кливлендских сланцев и других мест, а также тщательная сортировка позднепалеозойских акул такими учёными, как Мэйси (Maisey), Зангерл (Zangerl) и Лунд (Lund).
Ctenacanthus sp. на Красном холме (Red Hill) представлен одним шипом плавника. Более полные окаменелости были обнаружены из отложений раннего карбона в Шотландии. Шотландские акулы были относительно небольшими (50 см), но экстраполяция шипа, найденного на Красном холме, позволяет предположить, что эта позднедевонская рыба была значительно длиннее.
У ктенакантидных акул есть пара спинных шипов, украшенных множеством тонких рядов бугорков. Это придает шипам характерный гребенчатый вид, отсюда и название (ctenacanthus = гребешок-шип). Вместо пластинчатых шипов, которые были у многих других ранних акул, шипы ктенакантид были цилиндрическими и заострёнными. Эти шипы, а также детали анатомии плавников и строения их жаберных дуг указывают на то, что ктенакантиды имеют общего предка с более продвинутыми гибодонтами позднего палеозоя и мезозоя и неоселахиями (современными акулами).
%201f%20fossil.jpg) |
%201f.jpg) |
%202f%20fossil.jpg) |
%201f%20fossil.jpg) |
%201f.jpg) |
%201f.jpg) |
%202f%20fossil.jpg) |
%201f.jpg) |
%201f%20fossil.jpg) |
%202f%20fossil.jpg) |
 |
 |
 |
 |
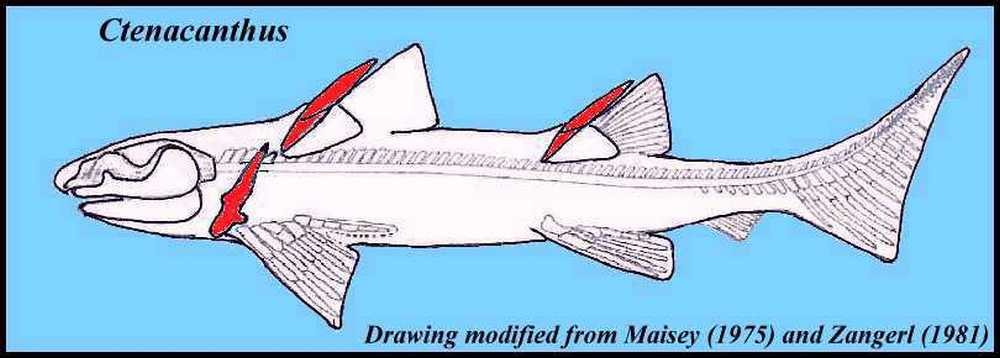 |
 |
 |
 |
Семейство Ктенакантиды † Ctenacanthidae Dean, 1909, Род Евнемакантусы — † Eunemacanthus St. John & Worhten, 1883 (известен из девонского, каменноугольного и пермского периодов, от 360,7 to 268,0 Ma до 221,5 миллиона лет назад) состоит из следующих видов: типовой вид — Eunemacanthus (Ctenacanthus) costatus Newberry & Worthen, 1866; Eunemacanthus heterogyrus McCoy, 1855; Eunemacanthus keytei Branson, 1916; Eunemacanthus venator Khabakob 1928.
Род Eunemacanthus описан по шипу плавника, который первоначально был отнесён Ньюберри и Уортеном (1866) к Ctenacanthus, но впоследствии был переведён в новый род (St. John and Worthen, 1883). Агассис (1837) перечислил шипики Ctenacanthus heterogyrus, но они были описаны и обозначены только позже (McCoy, 1855). Этот вид, вероятно, можно отнести к Eunemacanthus на основании морфологии плавника. Ископаемые останки евнемакантусов были найдены в Европе (Англия) и Северной Америке (Вайоминг, США).
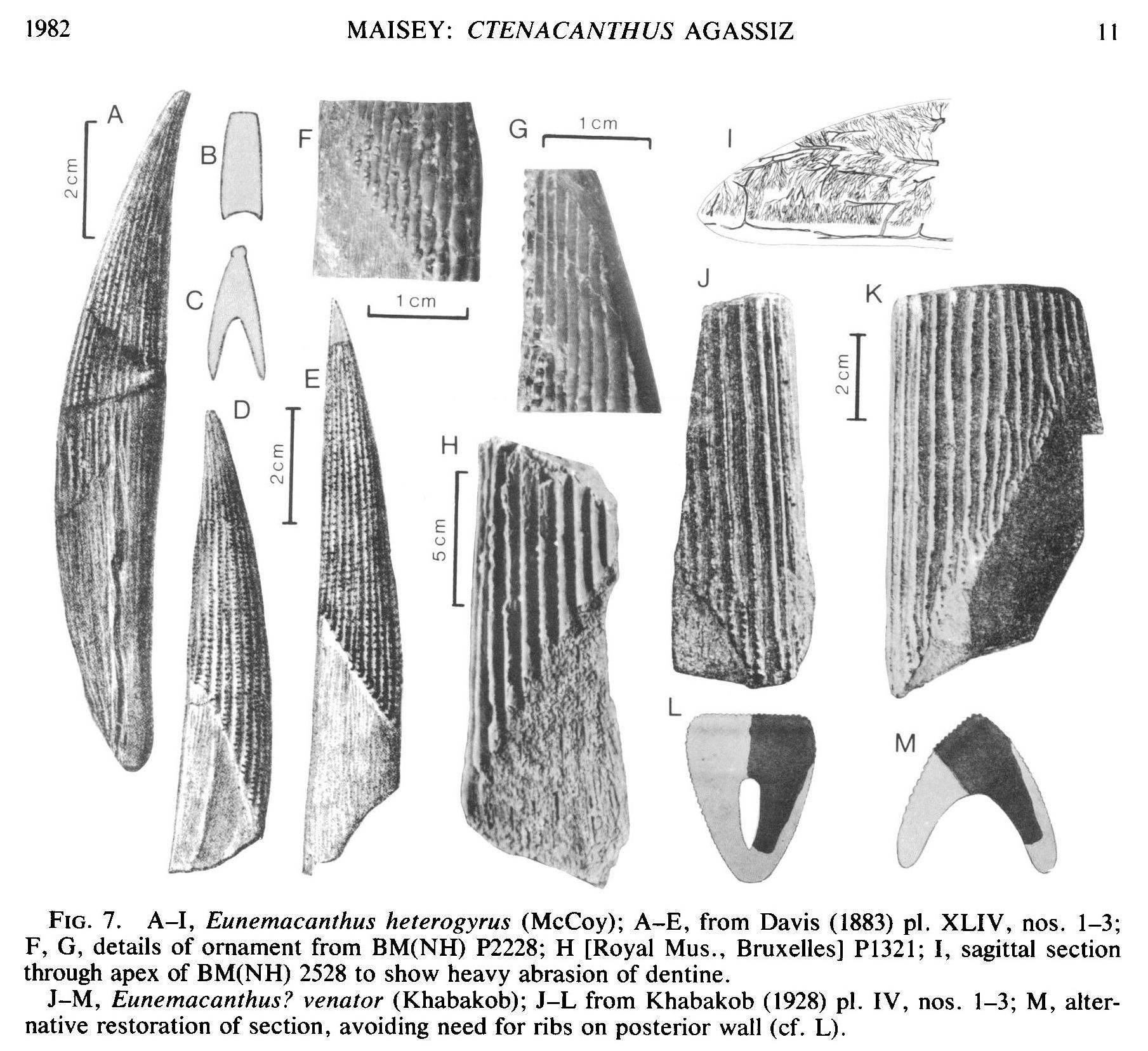 |
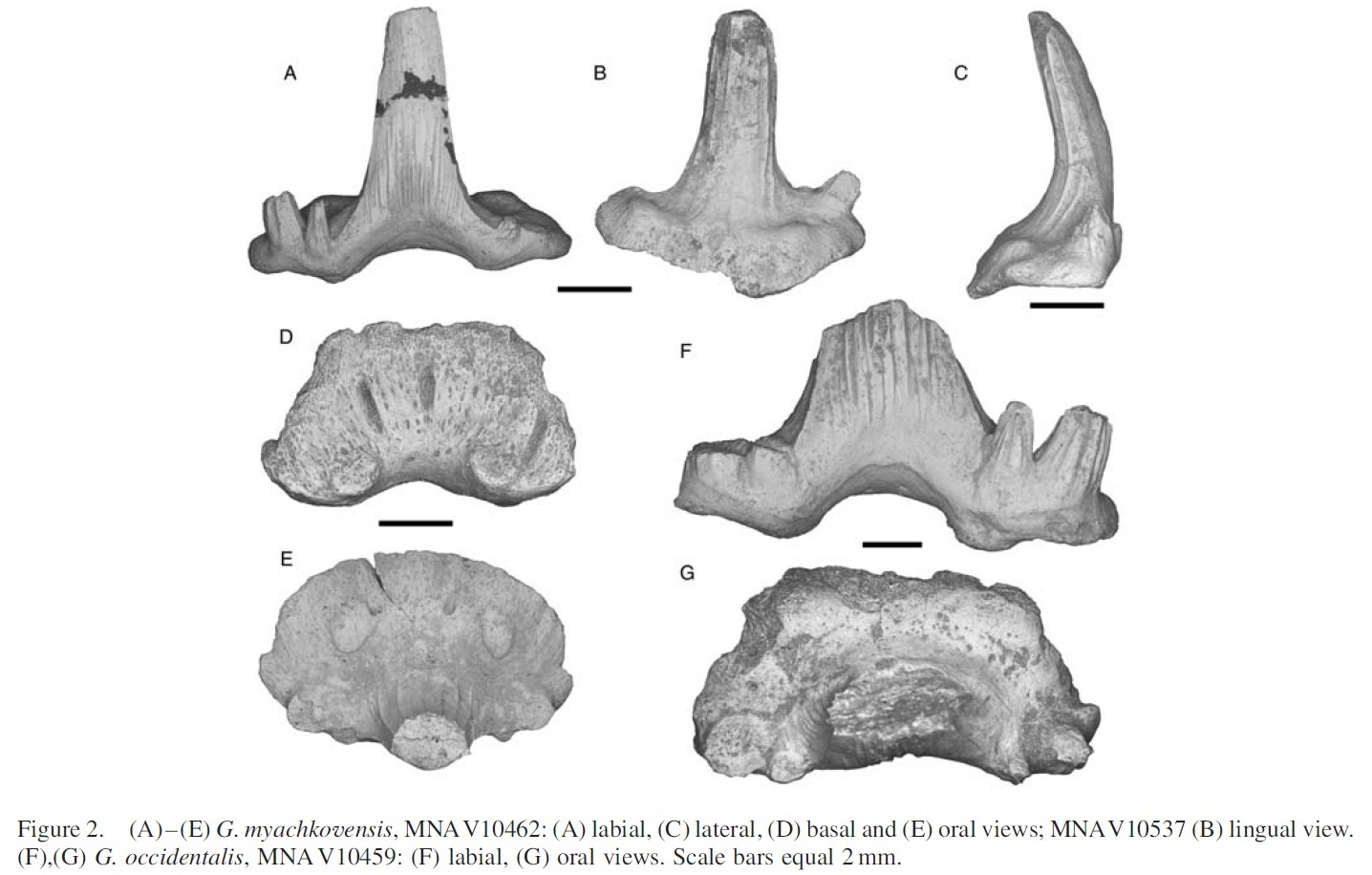
Семейство Ктенакантиды † Ctenacanthidae Dean, 1909, Род Гликманиусы — † Glikmanius Ginter, Ivanov & Lebedev, 2005 (известен из каменноугольного и пермского периодов, от 306,95 to 268,0 Ma до 254,0 миллиона лет назад) состоит из следующих видов: Glikmanius culmenis Koot, Cuny, Tintori & Twitchett, 2013; Glikmanius myachkovensis Lebedev, 2001; типовой вид — Glikmanius occidentalis Leidy, 1859. [M. Ginter, A. Ivanov, and O. Lebedev. 2005. The revision of "Cladodus" occidentalis, a late Palaeozoic ctenacanthiform shark. Acta Palaeontologica Polonica 50(3):623-631]
Ископаемые останки Glikmanius были найдены в Северной Америке и России. Род основан на целом экземпляре из Небраски, США. Род Гликманиус (Glikmanius) назван в честь русского палеонтолога доктора Леонида Гликмана (Dr. Leonid Glikman), который изучал этот род и был «первым, кто предположил его ктенакантообразное родство».
Glikmanius occidentalis (Leidy, 1859). Упомянутые образцы: зубы, MNAV10434, 10459, 10539. Местонахождение и горизонт: MNA LOC 1645, Качина. Нижняя часть пачки ископаемых гор; MNA LOC 118, юго-западный Флагстафф (Flagstaff,), Harrisburg Member.
Расстояние между зубами от 17 до 4 мм мезиодистально. Срединный выступ узкий, с грубыми гребешками. Боковые бугорки узкие и составляют примерно половину длины срединой куспиды. Промежуточные куспиды расположены впереди медианного и латерального бугорков, специфическая черта для Glikmanius occidentalis (Гинтер и др., 2005). На куспидах кили отсутствуют. Основание зуба трапециевидное с закругленным краем. Оролингвальный гребень и базолабиальные выступы образуют два отдельных дискообразных выступа. Оролингвальный гребень расположен близко к язычной границе основания зуба. Базолабиальная впадина хорошо развита и относительно глубокая (Рисунок 2: F, G).
Встречаемость Glikmanius occidentalis в фауне формации Кайбаб (Kaibab Formation) около города Флагстаффа (Flagstaff), штат Аризона, неудивительно, поскольку Ginter et al. (2005) отметили коллекцию Glikmanius occidentalis из пермской группы вблизи города Каунсил-Гроув (Council Grove), округ Уобонси (Wabaunsee County), штат Канзас. Однако в отличие от Канзаса записи о Glikmanius occidentalis в Кайбабе встречается редко, хотя встречается как в окаменелостях Mountain Member, так и в Harrisburg Member. Джонсон (2008) отметил наличие небольшого экземпляра Glikmanius occidentalis из нижней перми в группе (скоплении) Уичито (Wichita Group) из Техаса, но эта акула считается очень редкой в этой группе. Брэнсон (1933) отметил фрагмент срединной куспиды, идентифицированной им как «Cladodus occidentalis» из средней части формации Фосфории (Phosphoria Formation) в Вайоминге. По состоянию экземпляра, указанного в его описании (Брэнсон, 1933, рис. 1; номер 16, с. 182), трудно сказать, Glikmanius это или нет. Только Брэнсон (1933) отметил, что он был похож на другие образцы, найденные в нижней части формации Фосфории, которая считается пенсильванско-пермской границей.
В настоящее время лишь несколько экземпляров Glikmanius occidentalis были собраны в деревне Качина (MNAV10434 и V10459). Одиночный большой, но фрагментарный (17,2 мм мезиодистально) образец основания зуба был найден сотрудниками из Северного Университета Аризоны (MNA LOC 118). Этот образец отличается от других более распространённых образцов Glikmanius myachkovensis промежуточным бугорком, который находится явно впереди срединного выступа, и оролингвальным гребнем, расположенным ближе к языковому край основания зуба.
Glikmanius myachkovensis (Lebedev, 2001). Упомянутые образцы: зубы, MNA V10443, 10447, 10448, 10456, 10462, 10537; сорок зубов MNA V10481, 33 зуба MNAV10481. Местонахождение и горизонт: MNA LOC 1645, Качина. Нижняя часть пачки ископаемых гор; MNA LOC 118, юго-западный Флагстафф (Flagstaff,), Harrisburg Member.
Эти зубы имеют мезиодистально размер от 3,5 до 11,9 мм. Срединный бугорок узкий с крупным гребнем (кристой). Узкие боковые бугры расположены ниже срединной куспиды. Промежуточные куспиды находятся между срединным и боковыми зубцами, на одной линии с боковыми бугорками. Кили отсутствуют на всех бугорках. Основание зуба трапециевидное с закруглёнными краями. Оролингвальный гребень в виде двух отдельных дисковидных выступов на язычном краю основания зуба расположен почти посредине между средним выступом и язычной границей. Базолабиальный гребень состоит из двух закруглённых выступов, разделённых глубоким базолабиальным углублением. Языковая граница основания зуба слегка отклонена вентрально в сторону оролингвального гребня, это отличный признак для Glikmanius mayachkovensis (Ginter et al. 2005) (Рисунок 2: A–E).
Это второй по распространенности таксон, обнаруженный в Кайбабе. В формирование в районе Флагстаффа, особенно в деревне Качина, Glikmanius mayachkovensis встречается гораздо чаще, чем Glikmanius occidentalis. Ginter et al. (2005) заявили, что временной ареал Glikmanius myachkovensis находился в пределах позднего Пенсильванского периода, образец из Кайбаба, таким образом, расширяет временной диапазон Glikmanius myachkovensis в раннюю пермь.
В Мамонтовой пещере в начале 1990-х годов сотрудниками Фонда исследований пещер были обнаружены маленькие верхняя и нижняя челюсти (примерно 11 сантиметров в длину) с элементами жабр и несколькими зубами. Этот экземпляр был недавно исследован и идентифицирован. Зубы определяют этот частичный череп как маленькую ктенакантовую акулу по имени Гликманиус (Glikmanius). Что интересно в этом открытии, так это то, что это первый известный набор черепных хрящей этого вида акул, который когда-либо был идентифицирован. Кроме того, ранее Гликманиус был известен, прежде всего, из пенсильванских (323–298 миллионов лет назад) и пермских (298–251 миллионов лет назад) морских осадочных пород, что сделало окаменелости из Мамонтовой пещеры одним из самых древних примеров (335–340 миллионов лет назад) этого рода.
%201f.jpg) |
%202f%20teeth.jpg) |
%203f%20teeth.jpg) |
%204f%20teeth.jpg) |
%205f%20teeth.jpg) |
 |
 |
 |
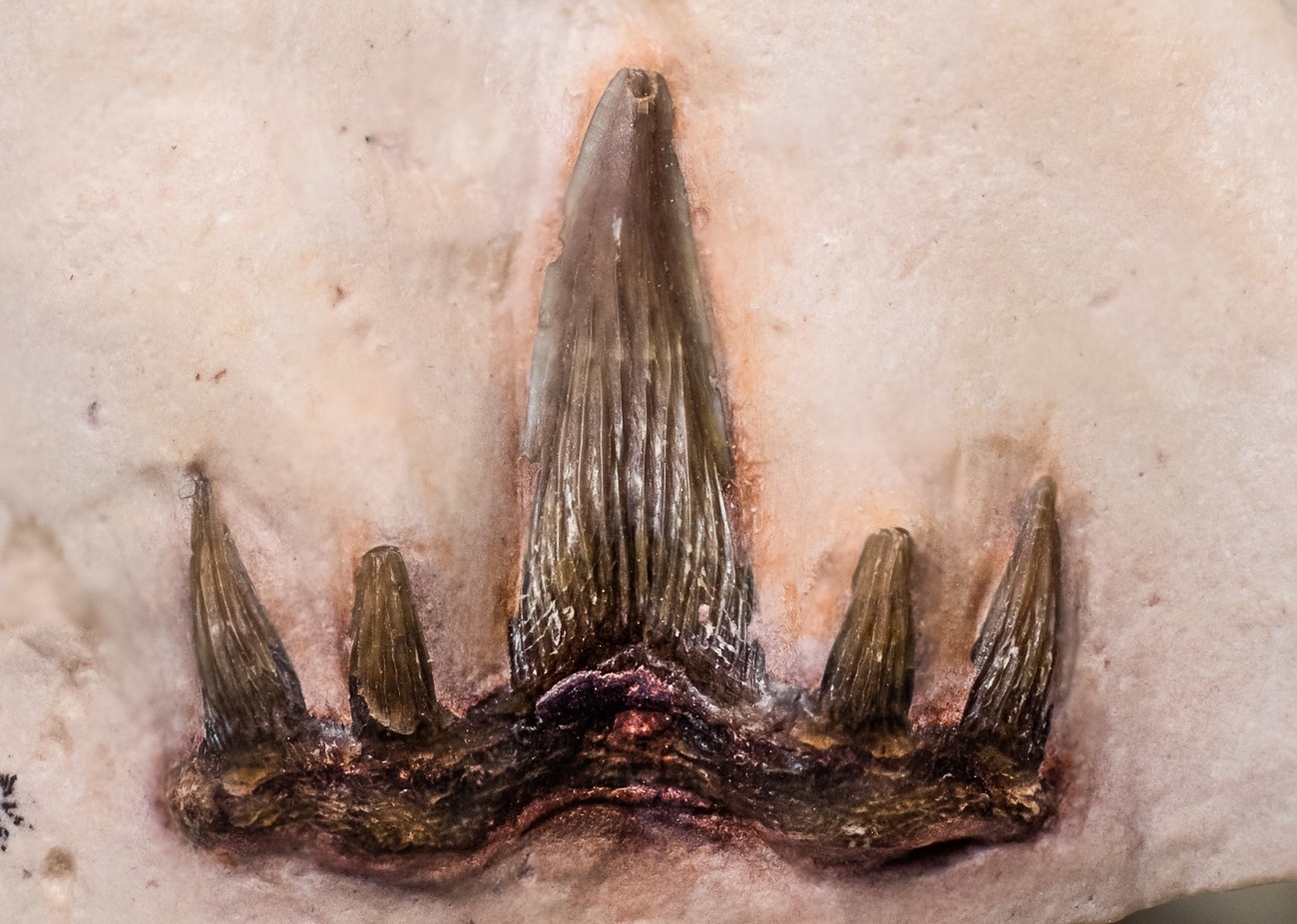 |
 |
 |
Семейство Ктенакантиды † Ctenacanthidae Dean, 1909, Род Глимматакантусы — † Glymmatacanthus St.John & Worhten, 1875 состоит из следующих видов: типовой вид — Glymmatacanthus irishii St.John et al., 1883; Glymmatacanthus petrodoides St.John et al., 1883; Glymmatacanthus rudis St.John et al., 1883.
Типовой вид Glymmatacanthus irishii описан на основании фрагмента шипа спинного плавника, USNM 13537. Шип имеет в поперечном сечении уплощённую форму и узорчатый орнамент, чем напоминает окаменелости шипов представителей рода Bythiacanthus (рис. 4D–G).
Другой вид, также описанный на аналогичном фрагменте шипа, это Glymmatacanthus rudis (рис. 4A–C). Роды Bythiacanthus и Glymmatacanthus временно сохранены как отдельные таксоны, но могут оказаться синонимами. Некоторые виды, такие как Glymmatacanthus petrodoides St. John & Worthen (1883, с. 250), основаны на очень отрывочных материалах, о которых мало что можно сказать (рис. 4H–K).
 |
Семейство Ктенакантиды † Ctenacanthidae Dean, 1909, Род Тамиобатисы — † Tamiobatis Eastman, 1897 (известен из девонского и каменноугольного периодов, от 360,7 до 353,8 миллиона лет назад) состоит из следующих видов: Tamiobatis springeri St. John & Worthen, 1875; Tamiobatis succinctus St. John & Worthen, 1875; типовой вид — Tamiobatis vetustus Eastman, 1897; Tamiobatis wachsmuthi St. John & Worthen, 1875.
Tamiobatis vetustus — очень редкая палеозойская акула, первоначально описанная на хорошо сохранившемся отдельном черепе несколько сомнительного происхождения из Eastern Powell Co., Кентукки. Вторая, гораздо более крупная черепная коробка была отнесена к этому роду, но, как и голотип, полностью отделена от тела. Последний образец, обнаруженный на плоскости напластования в позднедевонских сланцах Кливленда, состоит из части челюстей, зубов, чешуек и отпечатка шипа, который демонстрирует, что это ктенакантообразная акула, отнесённая к семейству Tamiobatidae Hay, 1902 г. Хорошо сохранившаяся базикраниальная область образцов раскрывает ранее неизвестные детали картины кровообращения у этих акул.
В 2018 году ранее неизвестный науке вид древних акул описали ученые из Санкт-Петербургского государственного и Белорусского национального технического университетов. Животное, обитавшее на Земле около 345-350 миллионов лет назад, назвали Tamiobatis elgae — в честь известного эстонского палеоихтиолога Эльги Марк-Курик. Результаты исследования опубликованы в «Estonian Journal of Earth Sciences». У этой акулы очень необычные зубы, отмечают авторы статьи. Каждый представляет собой развернутую корону, где центральный зубец — в два раза выше и в три раза шире, чем другие зубцы. Зубы первобытного хищника усеяны различными каналами и неровностями, наростами и бугорками. Всё основание зубов занимает очень плотная и сложная сеть сосудистых каналов. У них значительно больше число бугорочков, чем у зубов других видов Tamiobatis, а также уникальная «корона», содержащая два столбца второстепенных высоких бугорков, по-разному направленных и искривлённых.
Исследователи сделали снимки зубов в разных ракурсах на микротомографах и сканирующих электронных микроскопах, после чего сравнили их с зубами других древних хищниц. «На микротомограмме видно, что в частоколе вершин зуба застрял зуб небольшой костной рыбы. У меня есть подозрения, что акула питалась мелкими лучепёрыми рыбками: схватила одну, а зуб так и застрял между вершинами в челюсти акулы. Вообще, представители Tamiobatis были не очень активными хищниками и ели достаточно мелкую пищу: молодых особей других рыб, нетвердых ракообразных и моллюсков», — говорит один из авторов работы Александр Иванов, доцент СПбГУ, кандидат геолого-минералогических наук.
Новый вид относится к хрящевым рыбам. Ранее ученые неоднократно находили их останки на территории Белоруссии, в основном в южной части страны. Находки включали в себя отдельные зубы, чешуйки и маленькие фрагменты хряща.
Выделить новый вид в роду Tamiobatis ученым помогли три зуба акул, найденные в скважине Туров-106 недалеко от города Житковичи (Гомельская область, Белоруссия). Уникальные окаменелости из палеозойской эры нашли геологи, которые в 1970-х годах опробовали керн скважин, пробуренных для поиска соляных месторождений. Породу растворили для извлечения микроостатков древних организмов, чтобы определить по ним относительный возраст пород. Оказалось, что на глубине около 200 метров есть слои глины, которые позволили зубам буквально законсервироваться и сохранить свою уникальную форму.
«Пожалуй, ни у одной из известных ископаемых акул не было такого количества вершин у зубов: четыре основных, несколько вершинок первого и второго порядков — всего более двадцати, — рассказал доцент СПбГУ кандидат геолого-минералогических наук Александр Иванов. — Мы изучили зубы на микротомографе и увидели, какая у них удивительно разветвленная система каналов: крупных, мелких, горизонтальных и восходящих, тогда как у большинства современных хрящевых рыб она гораздо проще».
Известно, что и современные, и крупные доисторические акулы способны в течение всей своей жизни менять зубы: если один выпал, ему на смену вырастает новый. Именно микротомографические исследования, проведённые с помощью оборудования Научного парка СПбГУ, позволяют исследователям больше узнать об эволюционных изменениях зубной системы рыб. Все же, как отмечает палеоихтиолог, уже сегодня понятно: со временем и количество зубных каналов у рыб уменьшалось, и цикл смены зубов становился гораздо короче.
По словам исследователя, хозяин необычных зубов был не самой крупной акулой: в длину представители рода Tamiobatis составляли около полутора-двух метров, а на их спинах располагались два плавника с шипами. В целом эти рыбы известны как довольно хорошие пловцы, обитавшие в неглубоких и тёплых морях. Кстати, один из найденных зубов подсказал ученым, как питались древние акулы.
Сегодня акул из группы Tamiobatis палеонтологи находят по всему миру: в США известны рыбы из Кливлендских сланцев, также подобные фрагменты были найдены в Германии, в Польше, даже в Подмосковье — в карьерах окрестностей города Серпухова. Теперь ученым известны зубы этой акулы, найденные на территории Белоруссии. Сейчас они хранятся в коллекциях Отдела геологии Таллинского технологического университета.
%201f%20teeth.jpg) |
%202f%20teeth.jpg) |
%203f%20teeth.jpg) |
%204f%20teeth.jpg) |
%201f%20fossil.jpg) |
%202f%20fossil.jpg) |
 |
Род (incertae sedis) Астероптихиусы — † Asteroptychius Agassiz, 1843 (известен из девонского и каменноугольного периодов, от 360,7 до 318,1 миллиона лет назад) состоит из следующих видов: Asteroptychius bellulus Saint John & Worthen, 1875; Asteroptychius bellus Branson, 1906; Asteroptychius elegans Newberry, 1889; Asteroptychius gracilis Newberry, 1897; Asteroptychius keokuk Saint John & Worthen, 1875; типовой вид — Asteroptychius ornatus Agassiz, 1843; Asteroptychius sanctiludovici Saint John & Worthen, 1875; Asteroptychius semiornatus M'Coy, 1848 (карбон, Ирландия); Asteroptychius tenellus Saint John & Worthen, 1883; Asteroptychius tenuis Saint John & Worthen, 1875; Asteroptychius triangularis Newberry & Worthen, 1870; Asteroptychius vetustus Saint John & Worthen, 1875. [L. Agassiz. 1843. Recherches Sur Les Poissons Fossiles. Tome I (livr. 18). Imprimerie de Petitpierre, Neuchatel xxxii-188]
Род Asteroptychius отличается от рода Acondylacanthus сплошными ребрами плавниковых шипов и мелкими бугорками между этими рёбрами.
%201f%20fossil.jpg) |
%202f%20fossil.jpg) |
Род (incertae sedis) Пикнотилакантусы — † Pyknotylacanthus Mutter & Rieber, 2005 (известен из триасового периода, от 247,2 до 235,0 миллиона лет назад) состоит из следующих видов: Pyknotylacanthus humboldtensis Davidson, 1919; типовой вид — Pyknotylacanthus spathianus Mutter & Rieber, 2005. [R. J. Mutter and H. Rieber. 2005. Pyknotylacanthus spathianus gen et sp nov, a new ctenacathoid from the Early Triassic of Bear Lake (Idaho, USA). Revista Brasileira de Paleontologia 8(2):139-148]
Название рода Pyknotylacanthus происходит от греческих слов πυκνος (pyknos) — «плотный», «густой», τυλ (tyl) — «выпуклость», «отвердевшая кожа», и ακανθα (akanta) — «колючка», «шип», и относится к большим, плотно расположенным бугоркам на шипе плавника.
Шип плавника сжат с боков, треугольной формы в поперечном сечении; примерно верхняя треть боковых стенок окаймленна продольными или слегка изогнутыми рядами густо расположенных, крупных, звёздчатых бугорков (у родов Bythiacanthus и Glymmatacanthus бугорков меньше и они менее плотно расположены); бугорки с широкой базальной пластинкой (у Glymmatacanthus она менее выражена или даже отсутствует); оба края задней стенки шипа изнутри окаймлены продольным рядом очень мелких бугорков, некоторые из которых на кончиках загнуты назад; задняя стенка без бугорков или иногда присутствуют очень маленькие бугорки с загнутыми кончиками (отсутствуют как у Bythiacanthus, так и у
Glymmatacanthus).
Pyknotylacanthus spathianus описан по образцам PIMUZ A/I 3730 шипа плавника и PMUZ Z/I 3730/1-9 дермальных зубчиков, обнаруженных у горячих источников возле Медвежьего озера (Bear Lake) на юго-востоке штата Айдахо, США. Видовой эпитет spathianus происходит от названия временного периода Spathian, относящегося к раннему триасу — времени датировки найденных окаменелостей.
Шип плавника имеет высоту 178,0 мм, максимальная ширина 34,0 мм и максимальная толщина 16,2 мм. Апикальная половина шипа изогнута кзади (рис. 1). Судя по форма шипа плавника и углу, под которым шип мог быть вставлен в тело рыбы (Maisey 1978: 664), образец PIMUZ A/I 3730 может представлять передний, а не задний шип плавника. По крайней мере, верхняя треть поперечно бороздчатой поверхности стенок покрыта округлыми, обычно овальными, близко расположенными бугорками, тогда как нижняя часть, почти две трети длины шипа, лишена какого-либо орнамента (рис. 2). Шип плавник довольно острый, треугольный в поперечном сечении. Передний край, возможно, первоначально был окаймлён овальными бугоркамии. Примерно равноудалённое расположение бугорков с обеих сторон значительно отличается (рисунки 2—3). Грубо говоря, бугорки выстроены в продольные ряды, но этот рисунок прерывается участками неправильного расположения бугорков, особенно на левой стороне (это не из-за кислотной обработки спинного шипа); бугорки по передней и задней линиям расположены ближе, чем бугорки по продольным линиям на боковых стенках. Кончик шипа плавника лишен какого-либо орнамента, вероятно, по причине износа; возле вершины и дальше по ости плавника на большинстве бугорков видны следы износа.
Вид сбоку: передний край округлый и сильно изогнутый, тогда как задняя стенка глубоко вогнутая, по очертаниям менее изогнута (рис. 3). Очень тонкая задняя стенка глубоко вогнута и лишена какого-либо орнамента, кроме боковых краёв, окаймлённых зубчиками. Бугорки различаются по размеру от мелких (0,8 мм в диаметре) в апикальной задней части до больших (2,5 мм) в базальной передней части, где так же небольшие бугорки встречаются среди более крупных. Большинство коронок бугорков кажутся слегка заострёнными. Примерно от 8 до 14 рядов спускаются от коронки вершины и обычно раздваиваются. Дополнительные мелкие бороздки могут по-разному исходить снизу вершины. На заднебоковых краях остатки, вероятно, двух рядов зубчиков (рисунки 3C, 4).
Кожные зубчики. Около 40 дермальных зубчиков (рисунки 6—7) были обнаружены непосредственно с шипом плавника. Несмотря на другие остатки рыб, найденных рядом, возможно, что самые многочисленные зубчики (рисунки 6A—D2 и 7A—D) принадлежат одному и тому же виду акулы, как и спинной шип, потому что они обнаружены в том же тонком слое, что и шип плавника. Образцы дермальных дентикул можно подразделить на группы, а некоторые могут быть предварительно идентифицированы либо по
кодированной системе ихтиолитов (Doyle et al. 1974; Твей 1979, 1984; Johns et al. 1997) или путём сравнения с другими образцами зубчиков Щележаберных, описанных в отчётах (более или менее современных).
Найденные дермальные зубчики имеют размер от 0,25 до 1,3 мм в длину (рисунки 6—7). Зубцы состоят из базальной ножки и апикальной коронки, обычно состоящей из, по крайней мере, главного бугорока, а у некоторых образцов зубчиков присутствует шейка — более или менее выраженное сочленение коронки с ножкой (см. рисунки 6F1, F2 и рисунок 7F). Ножки примерно ромбовидные, в большинстве зубцов, при взгляде снизу, пронизаны отверстием центрального канала (и иногда различными периферическими отверстиями). Бугорки могут быть тупыми, острыми, одиночными, множественными или разделёнными («зубчатыми»). Некоторые бугорки тупые, имеют или не имеют боковых выступов (редкие образцы, рис. 7E—H). Большинство дермальных зубчиков обладают заострённым центральным бугорком (частый и обычный тип, рис. 7A—D), по бокам которого расположены два боковых выступа, мезиальный гребень (задняя часть) и мезиальная «платформа», которая обычно представляет собой выпуклый передний гребень. Боковые выступы могут сужаться к передней части. У гребня может быть развит зубчатый край, имеющий от 1 до 3 зубцов. Мезиальный (задний) гребень довольно слабо развит. Главный куспид может быть продолговато-прямым, дугообразным, заострённым или тупым с широкой плоской или выпуклой поверхностью, а гребни могут быть сильно или слабо развитые. Форма ножки обычно отличается в зависимости от формы коронки, а форма и размер отверстий варьируется в зависимости от вида зубчиков.
Зубцы либо (редко) толстые, но обычно длинные, острые ланцетовидные, одно- или трёхзубчатые, редко тупые или чрезвычайно острые, обычно симметричные и имеют два боковых гребня или крыла (рисунок 7). Мезиальную платформу часто заменяет центральный гребень, который может быть зубчатым (рис. 7В). Два
зубца характерны тем, что демонстрируют коронку, которая имеет неправильную форму, многоугольная по очертаниям с вогнутыми сторонами и увенчанная несколькими рёбрами (рис. 7G).
Эти находки являются замечательным свидетельством «архаичной» ктенакантоидной акулы и подразумевают выживание этой загадочной группы при переходе через пермско-триасовую границу. Хорошо сохранившийся экземпляр особенно важен, потому что его спатийский возраст хорошо подтверждается фауной аммоноидей (Columbites parisianus), подходящей для межрегиональной и временной корреляции. Диапазон морфологии индивидуальных дермальных дентикул удивительно широк и предполагает, что индивидуальные вариации чешуйчатого покрова у многих ископаемых и современных акул в целом недостаточно хорошо изучены, чтобы использовать их как диагностические признаки на более низком таксономическом уровне.
Ещё один образец (проксимальная часть шипа плавника) описанный Дэвидсоном (1919), как Cosmacanthus humboldtensis, был найден у Западного хребта Гумбольдта (средний триас) в Неваде, США. Хотя шип плавника сохранился очень фрагментарно, он имеет относительно плотно расположенные бугорки с характерными базальными пластинками, как у Pyknotylacanthus spathianus. Этот вид временно включили в род Pyknotylacanthus, как Pyknotylacanthus humboldtensis. Этот вид можно отличить от Pyknotylacanthus spathianus по менее треугольному поперечному сечению шипа с более изогнутыми (выпуклыми) боковыми стенками и менее глубокой задней полостью.
%201f.jpg) |
%202f.jpg) |
%203f.jpg) |
%204f.jpg) |
%205f.jpg) |
%206f.jpg) |
%207f.jpg) |
Род (incertae sedis) Ромбакантусы — † Rhombacanthus Williams, 1985 (известен из верхнего каменноугольного периода) состоит из одного вида — † Rhombacanthus zangerli Williams, 1985 [M. E. Williams. 1985. The "Cladodont level" sharks of the Pennsylvanian black shales of central North America. Palaeontographica Abteilung A 190:83-158]
Частичный скелет Rhombacanthus zangerli был обнаружен в чёрных сланцах Пенсильванского периода в центральной части Северной Америки.
Род (incertae sedis) Водника — † Wodnika Münster, 1843 [синоним: Radamas Münster, 1843] (известен в основном из поздней перми, от 259,0 до 15,97 миллиона лет назад) состоит из следующих видов: Wodnika althausi Münster, 1840; Wodnika ocoyae Jordan & Gilbert, 1919; типовой вид — Wodnika striatula Münster, 1843. [G. Münster. 1843. Nachtrag zu der Beschreibung einiger merkwürdigen Fische aus den Kupferschiefern. Beiträge zur Petrefacten-Kunde 6:47-52]
Вид Wodnika ocoyae Jordan & Gilbert, 1919 датируется миоценом, от 23,03 до 15,97 миллиона лет назад.
Окаменелости обнаружены в Германии, Англии и России. Длина этих акул составляла около 1 м, а форма хвоста указывала на то, что они, вероятно, хорошо плавал. Внутри некоторых окаменелостей сохранился хрящевой скелет, что довольно редко встречается у вымерших акул.
Wodnika — известна также как акула Цехштейна (Zechstein shark). Zechstein (немецкий) — это название слоёв осадочных пород среднего и позднего пермского периода, расположенных в Европейском Пермском бассейне, который простирался от восточного побережья Англии до северной Польши. Имя Zechstein раньше также использовалось как единица времени в геологической шкале времени, но в настоящее время оно используется только для соответствующих осадочных отложений в Европе.
Водника, вероятно, была очень похожа на современную бычью акулу (bullshark — Carcharhinus leucas). При жизни она была маленькой акулой, которая плавала в море Цехштейна, которое покрывало большую часть северной Европы, где Wodnika вероятно, курсировала около дна в поисках добычи, например ракообразных. Когда она находила добычу, то использовала свои закруглённые зубы, чтобы раздавить твердые панцири и добраться до более мягкой плоти.
Хотя ископаемый образец хрящевого скелета Wodnika известен, наиболее распространёнными ископаемыми являются закруглённые зубы и спинные шипы. Эти шипы поднимались перед первым и вторым спинными плавниками, и в то время рассматривались учёными в качестве опоры для этих плавников. Шипы так же делали Wodnika трудной добычей для любых потенциальных хищников. Также известно, что у самцов Wodnika были класперы (claspers) — придатки на нижней стороне тела для передачи спермы. Это также позволяет идентифицировать самцов и самок этого вида, поскольку у самок эти класперы отсутствуют.
%201f.jpg) |
%201f%20teeth.jpg) |
 |