Abramis brama (Linnaeus, 1758)
Лещ, Обыкновенный лещ, Подлещик, Ласкирь, Берберка, Восточный лещ, Дунайский лещ, Азовский лещ, Аральский лещ, Каспийский лещ, Волжский лещ, Днепровский лещ, Донской лещ, Невский лещ, Псковский лещ, Ильменский лещ, Волховский лещ, Кубанский лещ, Средневолжский лещ, Уральский лещ, Двинский лещ, Северно-каспийский лещ, Каспийско-волжский лещ, Куринский лещ, Печорский лещ, Восточносибирский лещ, Камышовый лещ, Абрамис брама (Rus),
Bream, Common bream, Freshwater bream, Carp bream, Eastern bream, Danube bream, Aral bream, Bronze bream, Sweaty bream, Leshtsch, Buggenhagen’s carp, Pomeranian bream
(Eng)
Синонимы:
Abramis argyreus Valenciennes, 1844
Abramis brama (Le Sueur, 1819)
Abramis brama bergi Grib & Vernidub, 1935 Аральский лещ (Rus), Aral bream (Eng)
Abramis brama danubii Pavlov, 1956 Дунайский лещ (Rus), Danube bream (Eng)
Abramis brama orientalis Berg, 1949 Аральский лещ, Восточный лещ, Восточносибирский лещ (Rus), Aral bream, Eastern bream (Eng)
Abramis brama sinegorensis Lukash, 1925 A hybrid between Blicca bjoerkna x Abramis brama (Eng), согласно Бергу 1949: 767, низовья реки Вятки, Россия.
Abramis brama vetula (Heckel, 1836)
Abramis gehini Blanchard, 1866
Abramis heckelii Selys-Longchamps, 1842
Abramis media Koch, 1840
Abramis melaenus Agassiz, 1835
Abramis microlepidotus Valenciennes, 1844
Abramis vetula Heckel, 1836
Abramis vulgaris Mauduyt, 1849
Brama lugdunensis Bleeker, 1868
Ciprinus latus Dralet, 1821
Cyprinus brama Linnaeus, 1758
Cyprinus brama subsp. major Forster, 1767
Cyprinus buggenhagii Bloch, 1784 A hybrid between Abramis brama x Rutilus rutilus (Buggenhagen’s carp) (Eng)
Материал подготовил Фактистов И. Е., 2024 г. Сайт www.fishbiosystem.ru
Вид Abramis brama описал в 1758 году, как Cyprinus brama, учёный Карл Линней (Карл Линней (Карл Линнеус) — швед. Carl Linnaeus, Carl Linné, лат. Carolus Linnaeus, 1707 — 1778, шведский естествоиспытатель (ботаник, зоолог, минералог) и медик, учился в Лундском университете (Lund University), затем в Уппсальском университете (Uppsala University), в 1732 году в одиночку совершил научное путешествие в Лапландию (Lapland), преодолев за пять месяцев более 2000 км, в 1735 году получил степень доктора медицины в университете Хардервейка (University of Harderwijk), между 1735 и 1738 годами жил и учился за границей, где опубликовал первое издание своей «Systema Naturae» в Нидерландах, с 1741 года до конца жизни — профессор Уппсальского университета (Uppsala University), создатель единой системы классификации растительного и животного мира, участвовал в создании Шведской королевской академии наук (Royal Swedish Academy of Science), был избран членом многих академий и научных обществ, с 1959 года Линней считается лектотипом вида Homo sapiens, является автором описания более 400 новых таксонов рыб). [Linné, C. (1758) Systema Naturae per regna tria naturae, regnum animale, secundum classes, ordines, genera, species, cum characteribus differentiis synonymis, locis. Ed. X., 1: 824 p. Stockholm (L. SALVIUS)]
Лещ, Обыкновенный лещ, Подлещик, Ласкирь, Берберка, Восточный лещ, Дунайский лещ, Азовский лещ, Аральский лещ, Каспийский лещ, Волжский лещ, Днепровский лещ, Донской лещ, Невский лещ, Псковский лещ, Ильменский лещ, Волховский лещ, Кубанский лещ, Средневолжский лещ, Уральский лещ, Двинский лещ, Северно-каспийский лещ, Каспийско-волжский лещ, Куринский лещ, Печорский лещ, Восточносибирский лещ, Камышовый лещ, Абрамис брама (Abramis brama) — вид лучепёрых рыб из отряда карпообразные (Cypriniformes Bleeker, 1859) семейства карповые (Cyprinidae Rafinesque, 1815), обитающих в пресных и солоноватых водах Европы и прилегающей Западной Азии. Является единственным видом монотипического рода Abramis Cuvier, 1816.
Название рода Abramis происходит от греческого слова αβραμις (abramis) — древнего название леща или кефали. По мнению Бикеса (Beekes), это либо догреческое слово, либо заимствование из египетского, означающего «тиляпия». Видовой эпитет биномена brama, происходит от abramis (греч. αβραμις), древнее название леща — «колючий лещ». Английское слово «bream» — «лещ» происходит от среднеанглийского «breme» — названия леща, широко распространённой европейской пресноводной рыбы. Первое известное использование слова «bream» относится к 14 веку. В свою очередь, «breme» происходит от англо-французского слова (от старофранцузского «braisme» — «лещ») германского происхождения (ср. древневерхненемецкое «brahsima»). Название также давалось различным похожим рыбам в других местах. Русское название «лещ» и украинское — «лящ» (Этимологический словарь русского языка Макса Фасмера) происходит от праславянского языка и, несомненно, связано с латышским «leste» — «плоский» и эстонским «lest» — «камбала (речная)». Есть также версия, что бурное поведение леща во время нереста — шлёпанье о воду и шумное плескание («лескание» или «ласкание»), послужило поводом для названия рыбы — «лещ» или «лящ» (в Словакии — «плескач»). Abramis brama называют на юге лящ, на Дону чебак, мелкий лещ на Азовском море — ласкирь, на Волге старые — чебаки, средние — киляки, молодые особи практически везде — подлещик. Рыбаки часто ласково называют леща — «Лёша». В разных районах России существуют свои характерные названия для Abramis brama (Linnaeus, 1758) (см. ниже «Словарь названий пресноводных рыб СССР.» Г.У. Линдберг и А.С. Герд 1972 г.).
Найденные окаменелости (fossils) отнесённые к Abramis brama (наиболее ранние) были датированы средним плейстоценом (Бихарий — Biharian, 0,8–0,1 млн лет назад — Бихарский ярус). Окаменелости аналогичны возрасту пласта Кромерского леса. Местонахождение ископаемых останков: Литл-Оукли — Little Oakley, Соединенное Королевство (United Kingdom) (51,9° с.ш., 1,2° в.д.; палеокоординаты 51,9° с.ш., 1,2° в.д.). Впервые обнаружены в 1939 году. Умеренный климат, региональная растительность смешанных листопадных лесов, обширные сухие луга в пойме реки и болотистые районы вблизи реки. Окружающая среда — речная; литология — нелитифицированный алеврит и песчаник, ископаемые илы и пески. Материал взят из скважин и ям из широкого русла ископаемого ила и песка. 50 кг насыпных образцов были взяты из последовательных захватов толщиной 25 см на глубину 350 см, что составило 1,5 тонны осадка и просеяно во влажном состоянии через сита 1 см и 1 мм. Весь материал хранится в Университетском музее зоологии (University Museum of Zoology) в Кембридже, а небольшое количество — в Музее естественной истории или Музее естествознания (British Museum of Natural History — BMNH) в Лондоне. [A. M. Lister and J. M. McGlade. 1990. The Early Middle Pleistocene vertebrate fauna from Little Oakley, Essex. Philosophical Transactions of the Royal Society of London B 328:359-385]
Окаменелости Abramis brama также были найдены в другом районе Англии, средний плейстоцен (0,8—0,1 млн лет назад). Местонахождение ископаемых останков: Пресноводное дно Вест-Рантон (West Runton Freshwater Bed), Формация Кромерского леса (Cromer Forest Bed Formation), Норфолк (Norfolk), Соединенное Королевство (United Kingdom) (52,9° с.ш., 1,2° в.д.; палеокоординаты 52,9° с.ш., 1,2° в.д.). Умеренный климат. Умеренные леса, травы и разнотравье с ольховыми болотами, тростниковыми болотами и водной растительностью поблизости. Окружающая среда — эстуарно-пойменная; литология — речной гравий, отложения глин и илов, а также песков. [A. J. Stuart. 1995. Insularity and Quaternary vertebrate faunas in Britain and Ireland. In R. C. Preece, ed., Island Britain: a Quaternary perspective. Geological Society Special Publication No. 96, London. 111-125]
Более ранние ископаемые останки отнесённые к cf. Abramis sp. (cf. — conformis, подобный, похожий) — представителя рода Abramis Cuvier, 1816 были датированы ранним миоценом (Бурдигалиан — Burdigalian, 20,44—15,97 млн лет назад — Бурдигальский ярус). Местонахождение ископаемых останков: Лангенау 1 (Langenau 1), группа пластов Кирхберг (Kirchberg beds Member), формация Браквотер Молассе (Brackwater Molasse Formation), Баден-Вюртемберг (Baden-Württemberg), Германия (48,5° с.ш., 10,1° в.д.; палеокоординаты 48,7° с.ш., 8,4° в.д.). Окружающая среда — морское побережье, представляет собой переплетение отложений солоноватого водоема (морской песок) с отложениями гравийной реки, пришедшей с северо-запада, из горного массива Швабского Альба (Böttcher, 1987); литология — галечный, серый, жёлтый, песчанистый мергель и нелитифицированный, пиритовый, чёрный, серый, голубой аргиллит. Несколько тысяч останков позвоночных (рыб, амфибий, рептилий, птиц и млекопитающих) были извлечены из трёх ископаемых пластов. Окаменелости были обнаружены в ходе трехмесячных спасательных раскопок летом 1976 года и хранятся в Государственном музее естественной истории в Штутгарте (Staatliches Museum für Naturkunde Stuttgart — SMNS). [E.P.J. Heizmann, L. Ginsburg, and C. Bulot. 1980. Prosansanosmilus peregrinus, ein neuer machairodontider Felide aus dem Miocän Deutschlands und Frankreichs. Stuttgarter Beiträge zur Naturkunde, Ser. B. 58:1-27]
На территории Российской Федерации окаменелости Abramis sp. обнаружены в Ростовской области, поздний / верхний миоцен (Туролийский период, турол — Turolian, 8,7—5,3 млн лет назад). Местонахождение ископаемых останков: Морская-2, зона MN 12, Ростовская область, РФ (47,2° с.ш., 38,9° в.д.; палеокоординаты 47,5° с.ш., 38,4° в.д.). Окружающая среда — флювиально-дельтовая; литология — зелёный аргиллит. Материал собран с 1932 г. и хранится в Геологическом институте (ГИН) РАН, г. Москва и Ростовском государственном университете (РГУ), г. Ростов-на-Дону. [N. V. Zelenkov and A. V. Panteleyev. 2015. Three bird taxa (Aves: Anatidae, Phasianidae, Scolopacidae) from the Late Miocene of the Sea of Azov (Southwestern Russia). Paläontologische Zeitschrift 89:515-527]
Ископаемые останки вида † Abramis bliccoides Schtylko, 1934 (см. фото) — представителя рода Abramis Cuvier, 1816 были найдены на Украине, поздний / верхний миоцен (11,6—5,3 млн лет назад). Местонахождение ископаемых останков: Шкодова гора (Shkodova Gora), Одесская область, Украина (46,5° с.ш., 30,7° в.д.; палеокоординаты 46,8° с.ш., 30,1° в.д.). Окружающая среда — флювиально-дельтовая; литология не сообщается. [O.M. Kovalchuk, L.V. Gorobets, E.V. Syromyatnikova, I.G. Danilov, V.V. Titov, T.V. Krakhmalnaya, L.I. Rekovets and N. V. Zelenkov. 2017. Vertebrates from the Pontian of the Shkodova Gora Locality (Northwestern Black Sea Region, Upper Miocene). Paleontological Journal 51(4):414-429]
Голотип † Abramis bliccoides — верхний миоцен (MN 12—13), Павлодарская свита (стратотипический аналог таврских молассовых красноцветов понта). Почти полный отпечаток. Местонахождение ископаемых останков: Россия, Омская область, Горьковский районн, село Серебряное. Место хранения ископаемых останков: Центральный научно-исследовательский геологоразведочный музей имени академика Ф. Н. Чернышёва, № 53/3242. Вид был описан в 1934 г. Штылько Б. А. на основе морфологических различий с родственными видами. Реконструкция скелета † Abramis bliccoides на основе отпечатка скелета ПИН № 1763-78, Западная Сибирь (см. схему). Более подробное описание Abramis bliccoides смотрите в статье описания вида Blicca bjoerkna (Linnaeus, 1758).
Тело у леща довольно высокое, овальное и коренастое, с вытянутым овалом в поперечном сечении, сильно сжатое с боков (недаром рыбаки часто называют его «фанерой»). Наибольшая высота тела в 2,5—3 раза меньше его длины (без С). Спина приподнятая, за затылком резко поднимается вверх, образуя «горб», особенно у крупных особей. У недоедающего или плохо питающегося леща развивается так называемая «спина-нож» — край спины очень острый и плоский. У молодых лещей (подлещиков) тело более продолговатое. В среднем в длине тела у лещей (22 самки) из Невской губы длина хвостового стебля 15,1%, антедорсальное расстояние 57,3%, высота D 20,6% (Л.С.Берг, 1948). Спинной плавник высокий, но короткий, его начало немного позади места прикрепления брюшных плавников или над серединой вертикали между брюшным и анальным. Состоит из 3 жёстких неветвистых (неразветвлённых) и 9—10, обычно 9, мягких ветвистых (разветвлённых) лучей (D III 9—10). Анальный плавник длинный, его основание в два раза длиннее основания спинного плавника. Он имеет вогнутый край, передняя часть более развита, чем задняя. Начинается за задним краем основания спинного плавника, состоит из 3 жёстких неветвистых (неразветвлённых) и 23—30, чаще 25—29, мягких ветвистых (разветвлённых) лучей (А III 23—30). Между брюшными плавниками и анальным есть не покрытый чешуёй киль (см. рисунок), а перед спинным плавником находится свободная от чешуи борозда (Дорогин, 2011). Грудные плавники (P I 15—17) немного не доходят до основания брюшных (V II 8—9), иногда доходят или даже заходят (Л.С.Берг, 1948). Брюшные доходят до анального отверстия или до анального плавника (Л.С.Берг, 1948). Хвостовой плавник (C III 23—28, по данным FishBase, Ref. 2196, — C 19) сильно выемчатый, с более длинной нижней лопастью.
Голова со слегка вытянутой мордой и глаза сравнительно небольшие. У обыкновенного леща соотношение между размером глаза и длиной головы значительно меняется в течение жизни рыбы, в молодом возрасте это соотношение как у густеры (длина головы от кончика рыла до задней части жаберной крышки примерно в четыре раза больше диаметра глаза, а высота головы более чем в два раза больше), но у взрослой рыбы длина головы может быть в шесть-семь раз больше диаметра глаза, а высота головы в четыре-пять раз. Измерения проводятся по линиям, разделяющим глаз пополам в обоих направлениях. В среднем диаметр глаза около 2/3 длины рыла у особей более 10 см SL (см. рисунок). Рот полунижний, слегка повернутый вверх, без усиков, с толстыми губами и выступающей верхней челюстью, небольшой, но может сильно выдвигаться, образуя длинную трубку, направленную вниз (см. фото). Глоточные зубы однорядные, обычно по пять с каждой стороны (см. рисунок): 5—5, редко 6—5 или 5—6, в очень редких случаях встречается слаборазвитый зуб второго порядка (1,5—5,0 или 0,5—5,1). Жаберных тычинок на 1-й дуге 18—30, обычно 23—28. Чешуя циклоидная, среднего размера, плотно сидящая в коже (см. фото). Боковая линия в передней части слегка изогнута вверх (см. фото), l.l. 49 (10—15)/(6—8) 60. Чешуй в боковой линия: 49—60. Рядов чешуек над боковой линией: 10—15. Рядов чешуек ниже боковой линии: 6—8. В позвоночнике 43—47 позвонков. 2-й и 3-й позвонки плотно срослись (П. В. Тюрин). Число хромосом: 2n = 50. Количество хромосомных плеч (фундаментальное число): NF = 80.
Окраска у леща меняется с возрастом рыбы и зависит также от цвета грунта и воды. Озёрные лещи, как правило, темнее речных. У взрослого леща окраска спины варьирует от свинцово-серой до коричневой или черноватой с зеленоватыми тонами. Бока желтовато-золотистые, бронзовые, золотисто-коричневые, золотисто-зеленоватые. Брюхо желтоватое с золотистыми оттенками. Все плавники тёмные, серого цвета, часто с более тёмными краями. Окраска спины у молодых рыб (подлещиков) серая, серовато-серебристая, бока более светлые, серебристые с синеватым отливом, брюшко белое с серебристым оттенком. Плавники полупрозрачные, сероватые или черноватые, часто с голубыми или розовыми отблесками. С возрастом подлещики начинают сереть, плавники темнеют, постепенно в окраске тела появляется золотистый оттенок, но все плавники остаются серыми. Трёхгодовалые приобретают уже жёлто-золотистый оттенок с более тёмной спиной. В репродуктивный период у самцов на голове, плавниках и передней части тела появляются брачные бугорки белого или желтоватого цвета, так называемая «жемчужная сыпь», а основания брюшных и анального плавников приобретают красноватый оттенок. Тело их становится шероховатым на ощупь.
Максимальная зарегистрированная длина: 82,0 см, обычная длина 25,0 см. Наибольший зафиксированный вес (опубликованные данные) — 6 кг. Максимальный заявленный возраст (опубликованные данные) — 23 года, обычно до 12—14 лет (по данным FishBase). По другим данным максимальный зарегистрированный возраст составляет 32 года для экземпляра из Каспийского моря, лещ вырастает до максимальной длины около 90 см и достигает веса 9 кг. В июле 1912 г. в Финляндии, в озере Веси (Vesijarvi), около Lahtis, пойман лещ весом 11,55 кг (Fiskeritidskrift for Finland, 1930, № 3, p. 43). По состоянию на 2022 год текущий европейский рекорд по весу леща, пойманного с помощью удочки с катушкой, составляет 10,32 кг (22,8 фунта), он был пойман в Соединенном Королевстве. [British Record (Rod Caught) Fish Committee (19 July 2021).] В водоёмах средней полосы России обычно встречаются особи длиной 50—55 см, весом до 5 кг. Возраст такого леща около 20 лет. Средние размеры леща 30—34 см, вес 500—900 граммов, возраст 7—9 лет.
Из-за большого сходства густеры — Blicca bjoerkna (Linnaeus, 1758) с молодым лещом, рыбаки зачастую принимают молодь леща за густеру (см. фото). Но их можно отличить (см. видео) по более крупной и плотной чешуе у густеры, которая остаётся такого же размера как вдоль боковой линии, так и в области спины. У леща (подлещика) вдоль боковой линии чешуя крупная, но ближе к спине она становится заметно мельче, а у самого спинного плавника и вовсе мелкая, тогда как у густеры этот признак выражен не так заметно. Подсчет рядов чешуй от передней части спинного плавника до боковой линии является самым надежным первым шагом в определении вида. У обыкновенного леща 13 чешуек или больше, а у густеры 9—11, включая чешую боковой линии. Количество чешуек в боковой линии у густеры составляет 43—51 чешуй, наичаще 45—48, а у бронзового леща — 49 и больше, а чаще значительно больше 50. Также наиболее чётко эти два вида различаются цветом парных плавников (грудных и брюшных): если у леща плавники тёмно-серые, то у густеры эти плавники обычно красноватые с серыми кончиками (см. рисунок). Анальный плавник у густеры сравнительно короткий и менее выемчатый, чем у леща (см. рисунок). Конец основания спинного плавника не доходит до вертикали начала анального плавника. Глаза густеры очень большие по сравнению с головой, и эта характеристика также отличает её от обыкновенного леща, а также от всех других европейских карповых, за исключением уклейки — Alburnus alburnus (Linnaeus, 1758). Длина головы густеры от кончика рыла до задней части жаберной крышки примерно в четыре раза больше диаметра глаза, а высота головы более чем в два раза больше, независимо от возраста рыбы. У обыкновенного леща соотношение между размером глаза и длиной головы значительно меняется в течение жизни рыбы, в молодом возрасте будучи похожим на соотношения этих размеров у густеры, но у взрослой рыбы длина головы может быть в шесть-семь раз больше диаметра глаза, а высота головы в четыре-пять раз. Измерения проводятся по линиям, разделяющим глаз пополам в обоих направлениях. Глаз у густеры также расположен очень близко к концу рыла и близко к верхней части головы, но у леща это гораздо менее характерно (см. рисунок). Рот у Blicca bjoerkna косой, полунижний, маленький, не выдвигающийся в виде трубки, как у леща. Глоточные зубы у леща — однорядные 5—5, редко 6—5 или 5—6 (см. фото), у густеры двухрядные (см. фото): 2,5—5,2 или 3,5—5,2(3). Густера редко имеет слизь на своем теле, а если и имеет, то в небольшом количестве. Напротив, лещ почти всегда очень скользкий (сопливый), особенно в молодом возрасте.
Более крупные особи гораздо легче различить из-за более заметных различий в цвете и форме тела. К тому времени, как оба вида достигают половой зрелости, их становится сравнительно легко отличить друг от друга, чешуя густеры остается яркой, блестящей, серебристой на протяжении всей их жизни, в то время как чешуя обыкновенного леща начинает приобретать различные оттенки, от тёмно-коричневого до светло-охристо-жёлтого. Однако некоторые лещи действительно остаются серебристыми на протяжении всей своей жизни, в зависимости от среды обитания. Путаница возникает, когда половозрелую густеру и неполовозрелую особь леща бегло сравнивают в полевых условиях, потому что лещ очень быстро вырастает до тех же размеров, что и половозрелая густера, будучи ещё совсем молодым, когда они все почти всегда серебристого цвета.
Лещ от представителей рода Баллерусы, Лещи-баллерусы — Ballerus Heсkel, 1843 Синца (см. рисунок) и Белоглазки отличается меньшим числом ветвистых лучей в анальном плавнике (лещ — А III 23—30, синец — A III 34—44, белоглазка — A III 35—44) и меньшим числом позвонков (см. видео). Кроме того, у синца рот конечный, обращенный вверх, чешуя мелкая, а у белоглазки большие серебристые глаза. Менее высокотелая рыба — рыбец — Vimba vimba (Linnaeus, 1758). От леща он отличается более коротким анальным плавником и наличием хорошо заметного покрытого чешуёй киля на спине между спинным и хвостовым плавниками. В бассейне Амура обитают два очень похожих на леща вида карповых с длинным анальным плавником, очень высоким, сжатым с боков телом и маленьким ртом. От леща их можно легко отличить благодаря острой гладкой колючке в спинном плавнике. Это — Белый амурский лещ (White Amur bream) — Parabramis pekinensis (Basilewsky, 1855) с длинным, не покрытым чешуей килем на брюхе, простирающимся от грудных плавников до анального отверстия, и Чёрный амурский лещ (Black Amur bream) — Megalobrama terminalis (Richardson, 1846). В наших водах Чёрный амурский лещ очень редок, он внесён в Красную книгу РФ. Нередкие гибриды леща с другими карповыми демонстрируют промежуточные характеристики, и их достаточно трудно идентифицировать.
Распространение (карта): пресные и солоноватые воды Европы (к востоку от Пиренеев и к северу от Альп) и прилегающей Западной Азии (75° с.ш. — 40° с.ш., 11° з.д. — 73° в.д.). В Центральной и Северной Европе встречается в реках и озёрах бассейнов Северного, Балтийского, Белого, Баренцева, Азовского, Чёрного и Каспийского морей, то есть в большинстве европейских водосборов от Адура (Adour, Франция) до Печоры (бассейн Баренцева моря, восточная часть), в бассейне Эгейского моря, в озере Волви (Lake Volvi, Греция) и водосборах рек Струма (Struma) и Марица (Maritza), присутствует в Англии и Ирландии, за исключением северной Шотландии, Корнуолла и Девоншира. В Северной Двине лещ немногочислен и, вероятно, проник сюда по каналам из Волжской системы. В Закавказье встречается лишь в немногих местах (у Ленкорани и в озере Палеостоми, а также в Мингечаурском водохранилище). На территории России акклиматизирован в Сибири в озере Байкал, в бассейнах рек Обь, Иртыш и Енисей (карта). В 1930-х годах интродуцирован в Зауральские озёра. В Азии обитает в бассейне Мраморного моря (Турция), в Аральском море, в озере Балхаш и низовье Сырдарьи, в реках Сары-Су и Чу. На Крайнем Севере и на юге территории бывшего СССР его нет. В природе отсутствует на Пиренейском полуострове (Испания, Португалия), в Адриатическом бассейне, Италии, Греции, Албании, Шотландии, Исландии, Скандинавии к северу от Бергена (Норвегия) и 67° с.ш. (Финляндия). Местами завезён в Ирландию, Испанию и северо-восточную Италию. В пределах своего ареала распространения лещ образует две географические расы — западную и арало-каспийскую. В бассейнах наших южных морей (в низовьях Днепра, Дона, Волги и северной части Аральского моря), кроме пресноводного жилого леща, имеется ещё полупроходной лещ, кормящийся в солоноватой воде, а для икрометания подходящий к низовьям рек. В южной части ареала, в Средней Азии, встречается мелкая тугорослая камышовая форма.
Ареал каспийского леща — нижнее течение и дельты всех рек, впадающих в Каспийское море: Волги, Урала, Терека, Куры; речки Ленкоранского побережья, редко встречается в Самуре и Атреке. На Нижней Волге вид представлен жилой и полупроходной формами (Казанчеев, 1981; Иванов, Комарова, 2012).
Лещ встречается преимущественно в крупных и средних реках, водохранилищах и во многих больших и заливных озёрах, реже в проточных прудах и каналах. Наиболее многочислен в богатых питательными веществами водах: заводях, нижних частях медленно текущих рек, солоноватых эстуариях и тёплых и мелких озёрах. Он избегает холодной воды, густых зарослей растений и небольших быстрых каменистых речушек, а предпочитает большие глубины, стоячие или слаботекущие воды, тихие тёплые водоёмы с песчано-илистым, иловатым или глинистым дном с обильной прибрежной растительностью. Излюбленное место пребывания взрослого леща — глубокие яры и глинистые ямы в заливах рек и в проточных озёрах. Держится вдоль подводных обрывов, особенно если дно идёт уступами с наличием бровок (см. рисунок). На песчаном дне и местах с быстрым течением бывает только проходом, но часто посещает реки с умеренным течением и более твёрдым грунтом. Abramis brama — стайный донный вид, образует стаи (см. фото), состоящие из особей разного размера и возраста, часто в ассоциации с другими видами карповых. Молодые рыбы часто образуют совместные стаи с близкородственным видом густерой. Больших перемещений стаи леща не делают и могут длительное время посещать одни и те же места, если они богаты кормом.
Лещ способен длительное время переносить относительно высокие температуры воды (33—34 °С), но при температуре выше 28 °С скорость его роста значительно снижается. Требователен к качеству воды (оптимум: 10°C — 24°C; pH: 7,0—7,5; диапазон dH: 15—18), одним из первых гибнет при заморах и сбросах загрязнённых вод. Обладая умеренно светобоязненным характером, лещ проявляет максимальную активность в сумеречные часы и в пасмурные дни. Обычно, лещи придерживаются береговой зоны, не более 10—15 метров глубины, но в крупных озёрах могут опускаться на глубину более 20 метров. Днём более крупные особи держатся на глубине, а ночью перемещаются в сторону побережья, где обычно держатся неполовозрелые особи. На кормёжку выходят на мелководье близко к берегу, в заливы, старицы и заросли кувшинок по руслу, большей частью по ночам (см. рисунок). Лещи очень осторожны, при малейшем шуме уходят на глубину. Зимой, в самые суровые периоды, лещи собираются в стаи, насчитывающие даже тысячи особей, которые перемещаются в более глубокие и спокойные места водотоков, где проявляют пониженную активность или пребывают в латентном состоянии. Впрочем, лещ никогда не засыпает крепко, а в тёплую зиму часто активно перемещается по водоёму в поисках пищи. Живущие у устья Волги частью зимуют в море, а частью входят в реку. Нерестовые миграции свойственны лещам, обитающим в водохранилищах. Они перемещаются как в верховья, так и в устьевые участки рек, впадающих в водохранилища.
Лещ по происхождению — типично пресноводная рыба, обитающая во многих реках и озёрах. Однако на Каспии, как в Азовском и Аральском морях, лещ является полупроходным видом, значительно расширившим свой ареал обитания за счет выхода из устьев рек в опреснённые, а также в солоноватоводные участки, и освоившим более богатые по сравнению с реками кормовые площади моря. Взрослые особи переносят концентрацию соли 12,9 ‰, а возможно, даже 14 ‰, а икру могут оплодотворить даже в солоноватой воде при 10,2 ‰. В любом случае оптимальная концентрация соли составляет от 2 до 4 ‰. Молодь леща держится в море на меньших глубинах, чем вобла, и ареал нагула молоди меньше. В многоводные годы ареал нагула расширяется. Зону распространения сеголеток ограничивает изогалина 8—9 ‰. Наибольшее количество сеголеток нагуливается в водах соленостью до 4 ‰ и глубиной до 4 м (Левашина, 2020). Проходные популяции кормятся в море, но размножаются и зимуют в более глубоких частях нижнего течения крупных рек и главных рукавов дельт. Весенние репродуктивные миграции начинаются с таяния льда вверх по течению и одновременного прогревания морских вод. После завершения размножения рыбы спускаются вниз по течению и расходятся в море на места нагула. В конце лета и осенью, когда вода охлаждается до 8—10 °С, интенсивность питания леща в море снижается, происходит осенняя миграция вида в реки, где он остаётся на зимовку. Перед заходом в дельту лещ концентрируется в авандельте, где наибольшее его количество наблюдается в конце сентября и октябре. В реках массовый ход леща продолжается в течение октября, и лишь к концу ноября интенсивность миграции снижается (Казанчеев, 1981; Иванов, Комарова, 2012). В российском районе Каспийского моря лещи обитают на глубинах не более 4—5 м, но Книпович (1921) сообщает о поимке особей на глубине 14,6—16,5 м в иранских водах.
Лещ — это типичный всеядный бентофаг, питающийся растительностью, органическими остатками и живыми организмами, обитающими на грунте и в грунте дна водоёмов. Способен переходить на питание взвешенными частицами или фильтровать при высокой численности зоопланктон. Обыкновенный лещ (Common bream) — Abramis brama (Linnaeus, 1758) способен регулировать размер отверстий своего «процеживающего аппарата» в зависимости от размера кормовых объектов (см. видео). Характер питания взрослых рыб зависит от типа водоёма, но преимущественно ориентирован на животную пищу. Обычно в рацион входят личинки водных насекомых, главным образом, хирономид (мотыль), ручейников (трихоптеры), подёнок (эфемероптеры), разнообразные черви, трубочники (род кольчатых малощетинковых червей — Tubifex Lamarck, 1816 из семейства Наидиды — Naididae Ehrenberg, 1831), ракообразные, моллюски (ракушки и улитки, см. рисунок), органические остатки, детрит и различные водоросли. Незначительную долю в корме составляют воздушные насекомые (см. рисунок) и высшая растительность. Выбор червей и ракообразных обусловлен численностью в донной фауне данного водоёма и их доступностью на местах откорма. Излюбленным кормом леща являются двухстворчатые моллюски из рода Шаровки, Шарики — Sphaerium Scopoli, 1777 семейства Писидииды, Горошинки, Шаровки (Peaclams) — Pisidiidae Gray, 1857 = Sphaeriidae Deshayes, 1855 (1820), которых лещи высасывают из песка своим трубкообразным ртом. Весной Abramis brama истребляют много икры других, ранее отнерестившихся рыб. Любят они и раков, которых во время линьки иногда вытаскивают из нор. Крупные лещи могут проявлять «хищный» характер, питаясь мальками и мелкой рыбой, поэтому нередко попадаются на спиннинг, в основном на джиг (см. фото) и мелкие колеблющиеся или вращающиеся блёсны. Личинки питаются зоопланктоном. Мальки по достижении длины 30 мм переходят на питание бентосом. Анализ состава рационов леща различных возрастных групп Чудско-Псковского озера показывает, что основу пищи составляют личинки хирономид, планктонные ракообразные, моллюски, с ярко выраженным преобладанием личинок хирономид по сравнению с ракообразными. До двухлетнего возраста в пище преобладает рачковый планктон. Сеголетки питаются, главным образом, ветвистоусыми (босмины) и веслоногими рачками (копеподиты циклопов); двухлетки — ветвистоусыми (дафнии, босмины).
В солоноватых участках южных морей полупроходная форма леща питается преимущественно ракообразными (мизидами, кумовыми рачками), а в Азовском море также и червями — полихетами. Исследования содержимого желудков особей, пойманных в северной части Каспийского моря, показывают, что их рацион состоит из моллюсков семейства Сердцевидки — Cardiidae Lamarck, 1809 родов Адакны — Adacna Eichwald, 1838 (69% веса) и Монодакны — Monodacna Eichwald, 1838, кумовых ракообразных и корофиид (амфипод из семейства Corophiidae Leach, 1814), хирономид, полихет, гаммарид (амфипод из семейства Gammaridae Latreille, 1802), мизид и олигохет (Oligochaeta — подкласс кольчатых червей из класса Поясковых Clitellata Michaelsen, 1919), лещи также могут питаться планктоном (Muus, Dahlstrom, 1999).
Лещ ищет пищу на дне, используя выдвижной рот, чтобы «дуть» на мусор и обнажать добычу, выкапывать или высасывать из мягкого грунта кормовые объекты своим трубкообразным ртом. Выдвижной рот даёт возможность лещу добывать пищу из грунта до глубины 5—15 см (см. видео). После такого «обеда» на дне остаются характерные отпечатки — небольшие воронки диаметром около 15—20 см, называемые «лещовыми ямками». Как и другие белые рыбы, Abramis brama также образуют в водоёме так называемые кормовые маршруты, которые посещаются в разное время суток. Места кормежки можно обнаружить по цепочкам всплывающих пузырьков болотного газа, появляющихся на поверхности воды, которые высвобождаются, когда стая лещей «переворачивает» очередной участок дна. Большие стаи, особенно в крупных реках и озёрах, очень быстро расходуют запасы пищи в одном месте и затем уходят дальше в поисках корма. Наиболее активное питание летом, после нереста. В это время у лещей быстро повышается упитанность. Почти все время лещ проводит в придонных слоях воды. В теплую тихую погоду, чаще перед восходом и перед заходом солнца, лещ может подниматься к поверхности воды — «плавится», показывая спинной плавник и часть хвостового, издавая при этом негромкий всплеск. Осенью его аппетит немного ослабевает, зимой лещ тоже питается, но не так активно.
В некоторых небольших водоёмах лещ может встречаться в больших количествах, что иногда приводит к нехватке донного корма. Тогда Abramis brama вынужден питаться, фильтруя планктон с помощью своих жаберных тычинок, причем основной добычей являются дафни и циклопы (см. видео). По мере роста рыбы жаберные тычинки уже располагаются слишком далеко друг от друга, чтобы ловить мелкую добычу, и в таких водоёмах лещ не вырастет больше 40 см. Если лещ недоедает или плохо питается, у него может развиться так называемая «спина-нож» — «knife back», край спины становится острым и плоским.
В зависимости от популяции половая зрелость обычно достигается между третьим и пятым годами жизни, самцы созревают раньше самок и имеют меньшие размеры. Полупроходной лещ созревает раньше жилого. Азовский лещ становится половозрелым в возрасте 3—4 лет. В озёрах, где вид очень обилен, и в северном секторе своего ареала, где рост рыбы медленный, половая зрелость достигается в возрасте 10 лет, когда размеры особей составляют около 20 см. Самцы леща Чудско-Псковского озера в массе созревают в шестигодовалом, самки в семигодовалом возрасте. Только к 9 годам при достижении линейных размеров 36—38 сантиметров все особи становятся половозрелыми. Минимальные размеры половозрелых самцов — 30 см, самок — 32 см. Нерестовое стадо леща характеризуется большой растянутостью возрастного состава (шести — пятнадцатигодовики) и размерного ряда (30—58 см). [Концевая Н.Я. Современное состояние плодовитости леща Псковско-Чудского озера // Биологические ресурсы и продуктивность разнотипных водоемов Псковской области: сб. науч. трудов ГосНИОРХ. Вып. 249. Л.: Промрыбвод, 1986.]
Многие популяции начинают нерестовую миграцию осенью (особенно полупроходные формы), замедляют зимой и продолжают весной. Ход леща в Волгу со взморья начинается ещё в феврале подо льдом, а валовой ход бывает около середины апреля. Нерест леща проходит в период с апреля по июнь. Азовский лещ начинает подходить к устьям Дона для нереста в конце марта — в апреле. Нерест начинается здесь в конце апреля. Нерестовые миграции свойственны и лещам, обитающим в водохранилищах. Здесь лещи перемещаются как в верховья, так и в устьевые участки рек, впадающих в водоём. В озёрах и прудах лещи часто мигрируют в прилегающие каналы и другие водотоки. По Днепру Abramis brama для нереста мигрируют вверх по реке на расстояния до 100 км. В нерестовый период у самцов леща на голове и боках тела появляются многочисленные мелкие плотные тупоконические эпителиальные бугорки — «жемчужная сыпь», сначала белого, потом янтарно-жёлтого цвета (см. фото).
Начало нереста в средней и северной России не ранее первых чисел мая, на юге с середины апреля. Температура воды должна быть не ниже 12—15°C. В низовье Волги лещ нерестует при температуре 17—20° С, обычно наиболее интенсивный нерест происходит в начале мая. Продолжительность нереста от нескольких дней до нескольких недель, в зависимости от температуры воды (чем холоднее, тем дольше). Оптимальная температура — около 21 °С, нерест проходит в кратчайшие сроки — в течение недели. В случае внезапного похолодания лещ прерывает процесс нереста и продолжает его при более высоких температурах. Во многих водоёмах наблюдается несколько подходов леща на нерестилища, за что разные группы особей получали специфические названия: черемшеник, никольник, колосовик, вербовик, дубовик и другие, связанные с сопровождающими нерест леща явлениями природы (цветением растений) или праздниками. Так у донских лещей выделяются две биологические группы: лещ-вербовик, нерестящийся рано, и лещ-дубовик, отличающийся более поздним нерестом (Федоров, 1960).
В поисках подходящих мест для нереста производители перемещаются большими стаями. Одной из характерных черт нерестилищ в озёрах и водохранилищах является наличие зарослей водной и прибрежно-водной растительности, в основном осок, телореза, рдестов. Глубина на местах нереста незначительная — до 1 м, а иногда и до 0,5 м. В реках для нереста выбираются мелководные прибрежные участки со слабым течением, травянистые отмели, неглубокие заливы, богатые подводной растительностью, или с прошлогодней растительностью и затопленными кустарниками, а во время сильных паводков и затопленные луга. На мелких участках реки лещ может нереститься в местах с каменистым или гравийным дном и большим количеством крупных подводных камней. Самцы часто защищают нерестовые территории вдоль береговой линии. Обычно нерест очень шумный, происходит рано утром, ухаживания самцов энергичные и сопровождаются громкими всплесками по поверхности воды и среди растений (см. фото). Это лещи выпрыгивают из воды на высоту до полуметра и плашмя падают в воду (см. фото).
Нерест происходит в 1—3 этапа, с интервалом 7—14 дней. Самки полупроходного леща выметывают от 68 до 390 тысяч икринок, а самки жилой формы — 89—240 тысяч (30000—40000 икринок на кг массы или около 140000 у самок средней величины). Икра липкая, диаметром от 1,0 до 2,0 мм, размер икры увеличивается с возрастом самки. Лещи из Чудско-Псковского озера характеризуются высокой популяционной плодовитостью. Видовая плодовитость, рассчитанная на 8—9 разовое икрометание в течение жизни равна 1675,2—1884,6 тыс. шт. Наиболее плодовитыми являются десяти-тринадцатигодовалые особи, имеющие наибольшие абсолютные и относительные показатели плодовитости. Современная средняя плодовитость составляет 209, 5 тыс. икринок. (Н.Я. Концевая, 1986)
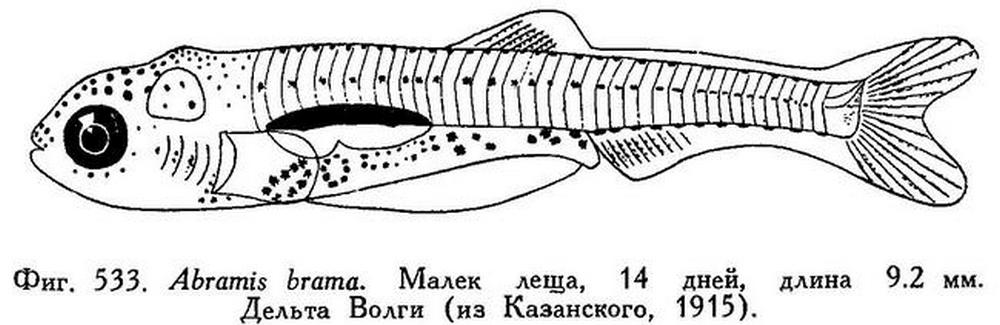
Abramis brama — типичный фитофил. Икра предпочтительно откладывается на водные растения или на залитую паводковыми водами прибрежную растительность (см. рисунок). При отсутствии таковой, самка леща приклеивает икру к камням или к подходящему древесному субстрату. После оплодотворения эмбриональное развитие происходит сравнительно быстро, личинки вылупляются через 3—12 дней в зависимости от температуры воды. Например, инкубационный период у азовского леща при температуре 18—20 °С продолжается около 3 суток. Выведшиеся личинки имеют длину около 3,9—4 мм и до всасывания желточного мешка неподвижно висят первые двое суток в стадии покоя, прикрепившись к водным растениям с помощью специальных клейких желез (см. фото). Затем личинки переходят к активному питанию сначала мелкими животными толщи воды — зоопланктоном, но очень скоро, по достижении примерно 30 мм длины, молодь леща переходит на питание донными организмами. Скат молоди леща из низовьев Дона в море происходит в середине июля. В дельте Волги скат выражен более резко, здесь он происходит в первой половине июля. Скатывается молодь леща преимущественно в дневное время. В море молодь распределяется в предустьевых участках и интенсивно питается. Взрослые отнерестившиеся лещи некоторое время «жируют» на местах нереста, а затем уходят в глубокие ямы рек или в опреснённые участки морей, и дисперсно распределяются в высококормных биотопах водоёма. Полупроходные лещи во время нагула держатся несколько дальше от устьев рек, чем молодь. Растет Abramis brama в различных регионах далеко не одинаково. Наиболее быстрым ростом обладает лещ в наших южных морях. Хуже всего растет он в водоёмах севера Европейской части России и в Финляндии.
Минимальное время удвоения популяции Abramis brama, низкое: 4,5—14 лет (K = 0,06—0,17; tm = 3—5; tmax = 17; Плодовитость = 90 000—340 000). Рост в условиях Нижнего Иртыша: возраст 1+, длина по Смиту 16—18 см, масса тела 80—100 г; в возрасте 3 года лещ достигает 25—28 см в длину (вес 320—370 г), а через 7 лет — около 30—40 см (1300—2100 г). Такой темп роста леща закономерен, так как в результате исследований было установлено до возраста 3 лет у леща идёт активный соматический рост, а после 3-летнего возраста начинается активное формирование половых продуктов. При этом следует отметить, что среди выловленных особей леща в возрастных группах 6+ и 7+ у самцов присутствовали медленнорастущие особи, что, вероятно, связано с какими-либо неблагоприятными условиями (низкий уровень залития поймы, низкое развитие кормовой базы) в период их соматического роста. [Либерман Е. Л. , Чемагин А. А. Размерно-весовые характеристики леща Abramis brama (Linnaeus, 1758) в условиях нижнего Иртыша // Современные проблемы науки и образования. 2017. № 1. С. 125.]
Гибриды леща с другими видами карповых, которые мало отличаются по времени размножения, местонахождению и поведению, являются обычным явлением. Известны плодовитые гибриды с Blicca bjoerkna и Rutilus rutilus. Так называемый померанский лещ (Pomeranian bream) — Cyprinus buggenhagii Bloch, 1784, вероятно, является гибридом обыкновенного леща и плотвы (см. рисунок). Более подробное описание гибридов можно найти ниже на этой странице в отрывке из книги Л.С. Берга «Рыбы пресных вод СССР и сопредельных стран.» (1948 г.).
Обыкновенный лещ подвержен заболеваниям бактериального и вирусного происхождения, и сам может являться хозяином и переносчиком различных видов паразитов. На этих рыбах обычно паразитируют моногенные трематоды (Dactylogyrus) и ленточные черви (Caryophyllaeus, Ligula, Digramma, см. рисунок). Цикл развития ремнецов родов Ligula и Digramma (см. рисунок) с двумя промежуточными хозяевами: первый — веслоногий рачок (циклоп, диаптомус), второй — пресноводная рыба (карповые, вьюновые, колюшки, бычки). Плероцеркоиды, развивающиеся в полости тела рыбы, — опасные паразиты, вызывающие массовые эпизоотии и гибель рыб. Попадая в кишечник рыбоядных птиц, они через 30—50 ч становятся половозрелыми ремнецами. Яйца для дальнейшего развития должны попасть в воду (см. видео). Фауна паразитических простейших леща Рыбинского водохранилища представлена 37 видами. Наибольшая численность паразитов и разнообразие их форм приходится на Волжский плес — 33 вида. По систематическому положению найденные паразиты распределились следующим образом: инфузории — 16 видов, миксоспоридии — 12, жгутиконосцы — 4, Dermocystidium — 1 вид (И. Я. Колесникова, 1994). Раненые и/или больные рыбы могут заразиться грибковыми инфекциями. На Abramis brama охотятся многие виды ихтиофагов и хищных рыб. Икра, личинки и мальки могут быть атакованы водными насекомыми, личинками насекомых и другими беспозвоночными, такими как ракообразные и пиявки. Мелкими особями леща питаются такие рыбы, как окунь, судак, налим, а более крупными — щука и сом. Лещ также является добычей многих рыбоядных хищников, таких как евразийская выдра (Lutra lutra, см. фото), европейская норка (Mustela lutreola), западная скопа (Pandion haliaetus, см. фото), кваква (Nycticorax nycticorax), большой баклан (Phalacrocorax carbo, см. фото), цапли (семейство Ardeidae, см. фото), поганки (семейство Podicipedidae, см. фото), чайки (см. фото), пеликаны и водяные змеи. В Каспийском море он входит в рацион эндемичного тюленя (Pusa caspica Gmelin, 1788).
В некоторых водоёмах лещ покрывается тёмными пятнами (точками) различных размеров (см. фото). Это болезнь пресноводных рыб — постодиплостомоз (постодиплостоматоз, неаскоз, чёрно-пятнистая или чернильная болезнь), вызываемая метацеркарией дигенетического сосальщика — трематоды Posthodiplostomum cuticola (von Nordmann, 1832) Dubois, 1936 из семейства Diplostomidae Poirier, 1886, класс Trematoda Rudolphi, 1808. Постодиплостомозом болеют более 40 видов рыб. Каждая точка — это место, в котором находится капсула с личинкой гельминта. Места локализации паразита у рыб: кожа, плавники, жабры, подкожная мускулатура, слизистая оболочка рта, роговица глаза. Жизненный цикл трематоды Posthodiplostomum cuticola протекает с участием трёх хозяев: первого промежуточного — брюхоногих моллюсков-катушек Planorbis planorbis, Planorbis carinatus и других, второго промежуточного (дополнительного) — рыб (в основном представители семейства карповых) и дефинитивного (окончательного) — голенастых птиц, питающихся рыбой, — серая, рыжая и желтая цапли, квакши. Постодиплостомоз пресноводных рыб встречается почти повсеместно, но наиболее широко распространён, как в естественных водоёмах и водохранилищах, так и в прудовых хозяйствах южных регионов России, что связано с обитанием на водоёмах большого количества рыбоядных птиц, особенно цапель. В организме рыбы метацеркарии сохраняются до 1,5 лет, хотя часть их погибает через 3—4 месяца. После гибели личинок паразита пигментные пятна или бугорки остаются. Находясь в теле рыбы, церкарии не выделяют опасных для человека токсинов. Согласно соответствующим ГОСТам при наличии на теле товарной рыбы единичных чёрных точек она допускается в продажу без всякой специальной обработки. Так что рыбу, поражённую постодиплостомозом, в пищу употреблять можно, с одной оговоркой. Рыба должна быть правильным образом приготовлена (сварена, пожарена, посолена), чтобы исключить возможность сохранения в ней в жизнеспособном состоянии каких-нибудь других паразитических организмов.
Вид Abramis brama по-прежнему остаётся относительно распространённым и многочисленным на большей части своего ареала. Лещ расширяет свой ареал за счёт безрассудных выпусков, производимых в целях его любительской ловли. На местном уровне вид может оказаться под угрозой исчезновения из-за строительства непреодолимых искусственных плотин и изменений качества воды, вызванных городским, промышленным и сельскохозяйственным загрязнением. На местном уровне некоторые популяции могут сокращаться. Учитывая важность спортивного рыболовства, в некоторых странах Центральной и Восточной Европы были введены минимальные размеры выловленной рыбы и периоды запрета для любительской ловли. В Красном списке МСОП (Международного союза охраны природы и природных ресурсов) вид отнесён к категории «Вызывающие наименьшие опасения» (LC).
Лещ — ценный промысловый вид, особенно в Северной и Центрально-Восточной Европе. Мясо оценивают по-разному, у экземпляров весом более 1 кг считают хорошим и вкусным и сравнимым с карпом. Мировой вылов достигал 62,9 тысяч тонн в 1999 году. Максимальный вылов в СССР составлял в конце 1930-х годов 120 тысяч тонн ежегодно, а в конце 1990-х годов отечественный вылов варьировал от 25 до 32 тысяч тонн. Основной промысел в реках и озёрах осуществляется неводами и мережами, а в море — ставными неводами и ставными сетями. Реализуется в торговле в вяленом, копчёном, свежем, охлаждённом и мороженом виде. Используется также для изготовления консервов, в том числе и икра леща (см. фото).
Обыкновенный лещ является объектом любительской рыбалки. В некоторых европейских регионах это добыча, которую ценят, прежде всего, за размеры, которых она может достичь. Ввиду своей всеядности лещ ловится на растительные и животные насадки (см. Список статей). Для ловли обычно используется поплавочная удочка (см. рисунок), а также фидер и различные виды донок и закидушек (см. рисунок). В период ледостава применяют снасти с насадками (червём, мотылём, репейником, чернобыльником или короедом) на мормышку (см. рисунок) или зимнюю поплавочную удочку и безнасадочные способы ловли (безмотылка, см. фото).
Считается, что мясо леща очень костлявое и сухое, но это касается только молодняка. Крупные нагульные лещи, особенно осеннего улова, содержат до 9% жира, мясо их жирное, нежное, мягкое. Несмотря на то, что в рыбе много мелких костей, крупных лещей заслуженно считают первоклассным рыбным товаром. У мелких лещей мясо тощее, суховатое, а обилие мелких костей особенно сильно снижает их качество. Лучшим лещом считается азовский осеннего улова. Это обычно наиболее крупная и упитанная рыба. Мясо леща превосходный источник калия, фосфора, жирной кислоты омега-3, в его состав так же входят кальций, магний, натрий, хлор, железо, хром, фтор, молибден, никель, витамин В1, В2, С, Е, РР, А и другие. Лещ оказывается одной из самых жирных рыб. Он по жирности уступает лишь уклейке и белуге. Жир леща наиболее полезен, потому что он не подвергается ни какой обработке, очищает сосуды, а витамин D содержащийся в рыбе хорошо укрепляет кости. Рыбий жир является уникальным продуктом. Он содержит полиненасыщенные жирные кислоты омега-3. По данным института питания именно эти кислоты снижают уровень холестерина в крови, растворяя холестериновые бляшки, что снижает риск инсульта, ишемической болезни, гипертонии. У рыбьего жира практически нет противопоказаний, за исключением индивидуальной непереносимости, которая встречается крайне редко.
Леща рекомендуют хранить при температуре 0 °С — 1 сутки, при температуре –5 °С — 14 суток, при температуре –10 °С — 4 месяца. Его фаршируют, запекают в духовом шкафу, филе используют в салаты и в качестве начинки для пирогов. Рыбу можно мариновать, солить, жарить (см. фото), тушить (см. фото), коптить (см. фото). Мясо леща сочетается с гарнирами из разных круп и тушёных овощей. Лещ образует гармоничные композиции с апельсином и лимоном, грибами и сыром, зеленью и соусами. Во многих странах пользуется большим спросом вяленый лещ (см. фото).
Великолепный рецепт приготовления леща под лимонно-апельсиновым соусом и оливковым маслом. Этот рецепт помогает сохранить весь жир в рыбе. Берётся укроп, мелко нарезается. Им наполняется внутренность леща. Затем выдавливается сок из одного апельсина и половины лимона. Соки смешиваются и выливаются на леща. Дают настояться 2—3 часа. Потом поливают 2—3 столовыми ложками оливкового масла сверху и вовнутрь, и запекают в фольге или просто в духовке. Рыбу ещё можно начинить гречневой кашей.
Атлас-определитель рыб: Книга для учащихся. Н.А.Мягков 1994 г.
* Лещ — Abramis brama (Linnaeus, 1758)
Внешне похож на густеру. Анальный плавник длинный. За затылком по голове и спине проходит борозда, лишенная чешуи. Глоточные зубы одно- или двухрядные. У обыкновенного леща рот полунижний. D 12—13. А 26—33. В боковой линии менее 60 чешуек (49—59). Жаберных тычинок 19—24. Окраска тела серовато-серебристая, спина темная. Грудные плавники ровные серые, остальные с более темными концами. Длина тела в среднем 50 см, масса до 6 кг. Широко распространен в пресных и реже солоноватоводных водоемах Европы (кроме севера) на восток до Эмбы (Закаспийская низменность). Ценный объект промысла, акклиматизированный во многих водоемах Сибири и Казахстана. Известен подвид:
Лещ восточный — Abramis brama orientalis
Длина тела не более 40 см. Обитает от южного течения Волги до реки Чу и на юг до Узбоя. Встречается в Тереке, Куре.
Иллюстрированная энциклопедия рыб. Ст.Франк 1983 г.
Лещ (Abramis brama) - высокотелая рыба, живущая, кроме рек Пиринейского, Аппенинского полуостровов, всюду в Европе. Достигает 70 см и 12 кг веса. На удочку удается поймать только редкие экземпляры. Более крупные особи в 4-5 кг ловятся в сети, главным образом ночью, вместе с плотвой, сазанами и другими карповыми рыбами. За один замет нередко ловится 8-12 центнеров рыбы. С рассветом лов сетью кончается. В связи с большим промысловым значением леща, рыбоводы заботятся о его размножении. Искусственные нерестилища из веток хвойных растений и мха размещаются и укрепляются в регулируемых водотоках. Рыбы используют их при откладывании икры.
Словарь названий пресноводных рыб СССР. Г.У.Линдберг и А.С.Герд 1972 г.
12.1.2 (1). Abramis brama (Linne, 1758) — Лещ (Б. : 768).
[1] русск. Берберка Крым — Кл. Беребёр пек.: гд., печ., середк. — ПОС; А. Г. Беребёрка (мелк.) — Гр. : 147; пек.: гд., печ., середк. — ПОС; А. Г.; твер. — Даль; крым. — Зол. Беребёрочка (мелк.) пек.: гд., печ., середк. — ПОС; А. Г. Беребра (мелк.) пек.: гд., печ., середк. — ПОС; А. Г.; твер. — Даль; крым. — Сабанеев : 501. Беребрина пек.: гд., печ., середк. — ПОС; А. Г. Беребро (мелк.) ЭССР; пек.: гд., печ., середк. — ПОС; А. Г. Беребрушка пек.: гд., печ., середк. — ПОС; А. Г. Берёзовый лещ Вышний Волочек; Осташков — Кессл., 1870 : 255. Благородный лещ — Кессл., 1864 : 90. Большина (крупн.) Ильмень, Волхов — Домр, и Правд. : 53. Большой лещ Ильмень, Волхов — Шам. Вимба — ПОС. Всякая рыба Днепр; Волга: казан. — Зол. Вся-рыба Днепр — Зол.; Волга: нижегор. — Варп., 1891 : 27—28; Волга: казан. — Зол. Галагалок (мелк.) пек.: печ. — ПОС. Головной лещ Ильмень: Поозерье; Волхов — Домр, и Правд. : 53. Дубовик казан.: р. Сура — Зол. Калиник Вышний Волочек, Осташков — Кессл., 1870 : 255. Карточка (мелк.) Ильмень, Волхов — Шам. Киляк (мелк.) Дон — Кл. Кляпуха (молодь) Волхов — Кессл., 1864 : 90. Кляпушина (мелк.) Волхов — Шам. Кляпушонка (мелк., весом до 1 фунта) Ильменьг Волхов — Шам. Колосовой лещ Вышний Волочек, Осташков — Кессл., 1870 : 255. Коростелёк (мелк., весом до 1 фунта) Ильмень, Волхов, Коростель (мелк., весом до 1 фунта) Ильмень, Волхов — Шам. Костлявка урал.: pp. Вишера, Кама, Колва — Ревн. : 217. Костляк Пек;, и Чудск. оз.: Сыренец — Кузн., 1915. Красик (мелк.) Белое оз. — Арнольд, 1925 : 13. Крупный лещ (более 30 см) пек.: гд. — ПОС; А. Г; Ильмень, Волхов — Шам. Ласкирь (мелк.) Азов. м. — Б. : 768. Ледка (мелк.) Ильмень, Волхов — Шам.; смол.— Добр. Ледочка (мелк.) Ильмень, Волхов — Шам. Лепестняк (мелк.) Ильмень, Волхов, Лётка (мелк.) Ильмень, Волхов — Домр, и Правд. : 53. Лещ арх.: озера Кенозеро, Лача — А. Г.; в. т. Вятки — Лукаш, 1925 : 38; карел.: Водлозеро — А. Г.; Белое оз. — Кучин : 349; р. Шексна — А.; ленингр.: ефим. — А.; пек.: повеем. — ПОС; А. Г.; Ильмень, Волхов — Варп., 18866 : 39; н. т. Волхова — А. Г.; новг.: озера Уклеино, Валдайское, старорус., pp. Ловать, Б. Тудер, М. Тудер; калин.: озера Вселуг, Стерж, Селигер — А. Г.; смол.: Днепр — Воронцов, 1930 : 143; pp. Сож, Ипуть, Остер — Воронцов, 1927 : 25; ср. т. Десны — Воронцов, 1928 : 72; брян. — КБС; кур.: р. Сейм — Гладков : 49; А.; р. Свапа — Гладков : 49; рыльск., льгов., глушк. — А.; орл.: Ока — Тарачков : 34; ворон. — А.; пенз. — Магн.: 19; Кубань — Есипов, 1930 : 46; н. т. Дуная — Гриц.; вся Волга — Кессл., 1870 : 255; нижегор.: pp. Ветлуга, Линда, Пьяна, Кудьма, Ока, Теша, Ворсма, оз. Валдайское; озера Киркидеевское, Сеухинское, Озерина, Светлое, Светлая Лужа — Варп., 1891 : 57—68; ср. т. Волги; симб. — BP. 1905, № 7 : 341; казан.: pp. Цивиль, Свияга, Б. Кокшага, М. Кокшага, оз. Таир, pp. Казанка, М. Черемшан — Варп., 1886а : 45—63; урал.: повеем. — Ревн. : 217; оренб., Урал. обл. — А.; Сырдарья, Амударья — Б., 1905 : 167; р. Чу — Г. Ник., 1931 : 257. Лещ-выросток (весом до 2 кг) Ильмень, Волхов — Шам. Лещ-плотичник (лещ, который нерестится одновременно с плотвой) Онеж. оз.: Чолмужи — ПоКров., 1936 : 64. Лещата (мн. ч.; мелк.) урал.: pp. Уфа, Урал, Синара — Ревн. : 217. Лещик Ильмень, Волхов — Шам.; Урал. обл. — А. Листок (мелк.) Белое оз. — Арнольд, 1925 : 13. Лопырёк (мелк., весом до 100 г) Ильмень, Волхов — Шам. Ляпок (мелк.) Белое оз., Ляпушка (мелк.) Белое оз. — Кучин : 349. Лящ южн. — Б. : 768; н. т. Дуная — Гриц.; урал. — Ревн. : 217. Мелкий лещ Ильмень, Волхов — Шам. Молдаванка Азов. м. — Кл. Нерестовый лещ (лещ, идущий через две недели после плотичника) Онеж. оз.: Чолмужи — Покров., 1936 : 64. Палтан (молодь) урал.: Кама — Меньш. и Букирев : 52. Палтуха (молодь) Кама: перм. — Меньш., 1928 : 396; pp. Кольтма, Колва, Пильва, Чусовая, Сылва — Ревн. : 217. Перечень новг. — Зол.; Ильмень, Волхов — Домр, и Правд. : 53. Плотичник (лещ, нерестующий одновременно с плотвой) Онеж. оз.: Чолмужи — Покров., 1936 : 64. Плотишник Вышний Волочек, Осташков — Кессл., 1870 : 255. Подлещ (мелк.) н. т. Волхова; новг.: озера Уклеино, Валдайское; калин.: оз. Вселуг — А. Г.; ворон.; кур.: льгов. — А. Подлещик (мелк.) арх.: озера Кенозеро, Лача; карел.: Водлозеро — А. Г.; сев.-зап. часть Онеж. оз. — Зборов. : 78; Пек. и Чудск. оз. — Кессл., 1864 : 90; Ильмень, Волхов — Варп., 18866 : 57; н. т. Волхова; новг.: озера Уклеино, Валдайское; калин.: оз. Вселуг — А. Г.; кур.: р. Сейм — Гладков : 49; А.; р. Свана — Гладков : 49; глушк., льгов., рыльск.; ворон. — А.; пенз. — Магн. : 19; Волга — Кессл., 1870 : 255; казан. — Варп., 1886а : 44; симб. — BP, 1905, № 7 : 341; урал.: pp. Кама, Кольтма, Колва, Пильва, Чусовая, Сылва — Ревн. : 217; Кама: перм. — Меньш., 1928 : 396; Урал. обл. — А.; среднеаз. — Б., 1905 : 167. Подлещичек карел.: Водлозеро — А. Г.; пек.: гд. — ПОС; А. Г. Полтан (молодь) урал.: pp. Кама, Кольтма, Колва, Пильва, Чусовая, Сылва — Ревн. : 217. Получебак, Синяк (мелк.) н. т. Волги — Зол. Средний лещ (длиной 20—30 см) пек: гд. — ПОС; А. Г. Сугава Белое м., Сухорёбка кур.: льгов. — А. Тарабара карел.:. Водлозеро, Тарабарка карел.: Водлозеро — А. Г. Ходовый лещ Ильмень. Волхов — Шам. Чабак Дон — Миртов; Кубань — Б., 19126 : (20. Чебак (крупн.) южн. — Кессл., 1864 : 90; Дон — Миртов; Моздок — Караулов; Азов. м. — Кл.; Кубань — Б., 19126 : 120; н. т. Дуная — Гриц.; зкв. — Барач, 1941 : 219. Черемошник Вышний Волочек, Осташков, Черемховый лещ Вышний Волочек, Осташков — Кессл., 1870 : 255. Черкесик — Зол. Чернокрылка Ильмень, Волхов, Черноперка Ильмень, Волхов — Варп., 18866 : 57. Чибак Дон — Миртов. Чубак н. т. Дуная — Гриц. Шабёр Дерпт — Зол.; Пек. и Чудск. оз.: Горки, Брага — Кузн., 1915. Ширман (мелк.) р. Самара — Кессл., 1870 : 255. Ширманщик (мелк.) pp. Волга, Самара, сарат. — Кессл., 1870 : 255. Щабёр пек., Щебёр — Гр. : 147. Щебёра пек. — ПОС; А. Г. Щебёрка Ильмень, Волхов — Кессл., 1864 : 90; Пек. и Чудск. оз. — А. Ник. : 680. ~ Лещ, лещь — Грам. кн. Мих. Андр., 1460 г.; Соймонов : 347; Росс, целл.; Гмелин, I : 3, 90, III, 2 : 340—341; Лепехин, I : 56; Паллас, I : 31, 202, 423—424, И, 2 : 15; Нордстет; Начерт. : 401; Зуев, 1787 : 228; Озерецк., 1791 : 79; Озерецк., 1792 : 36, 225; САР; Блуменбах : 413; Севергин : 13, 138; Озерецк., 1808 : 41, 43, 49; Озерецк., 1817 : 123, 130, 155, 172—173; Теряев : 53; Ловецкий : 488; Сл. 1847 г. Лещик — Кн. прих.- расх. Кир.-Белоз. мон., 1606—1607 гг.; Нордстет; САР. Подлещик — Кн. прих.-расх. Болд.-Дорог. мон., 1598—1600 гг.; Гмелин III, 2 : 341. Чебак — Паллас, II, 2 : 15; Сл. 1847 г. Lestsch, Peretschen (мелк.) новг., Podlestschi (мелк.) Дон, Podlestschik новг., Polutschabak Дон, Tschabak Дон — Pallas : 325. [2] укр. Девер — Колюшев : 27. Козел (молодь) Днепр, Буг — Марк. : 121. Козлик н. т. Днепра, Козловаха (молодь) н. т. Днепра, Козловашка (молодь) Днепр — Тат. : 77. Ляска (мелк.) Десна — Б., 1948 : 64. Лящ — Марк. : 121. Лящик, Пiдлящ (молодь), Пiдлящик (молодь) — Тат. : 77. Плискованя — Колюшев : 27. Таранник — Тат. : 77. Tpicкa — Колюшев : 27. Чабак полтав. — Тат. : 77. Чебак Сев. Донец, н. т. Дуная — Марк. : 121. [3] белор. Лещ, Подлещик (молодь) — Жуков : 69. [4] польск. Bleye мазур., кашуб., Brass мазур., кашуб. — Benecke : 118. Kleszcz — Majew. Leszcz мазур., кашуб. — Majew.; Staff : 157. ~ Kleszcz, Leszcz (старопольск.)— Budz. : 130. [5] чеш. Dlesec, Dlest — Budz. : 130. Velky cejn — Hrabe : 61 ~ Dlesc (старочеш.) — Budz.: 130. [6] словацк. Pleskac, Plieskac — Budz. : 130. [7] болг. Платина — Дренски : 102. [8] сербскохорв. Ain, Ajna, Anj, Anja, Crnovka, Dever, Devericanka, Deverika, Diver, Diverika, Djeverika, Gologlavka, Hajin, Hajma, Hajn, Hajna, Hanj, Hanja, Kecel, Ke- sezica, Kesega, Kesega deverika, Kesegica, Kesiga, Kesigica, Koscak, Koza, Kozara, Kozel, Kozica, Lapara, Ljeskar, Ljeskarak, Ljeskarica, Okaca, Pecenica, Platnica, Plosk, Ploska, PloSic, Senj, Sinj, Sinjac, Sinjaca, Sinjacica, Sinjakusa, Sinjarak, Sinjavac, Sinjevac, Sinjur, Sivac, Sivacica, Sivak, Struja, Siljan, Sinjozica, Sirajka, Susta — Hirtz. [11] н.-луж. Klesc — Budz. : 130. [12] латыш. Breksis — ZK. Bressen, Kaza — Гр. : 147. Plaudis —ZK. Skilts — Гр. : 147. [13] лит. Karszis — Majew. Palzas — Гр. : 147. Plakis — Majew. [14] дат. Brasen — Hoek : 41; O-n : 120. [15] шведск. Braxen — Smitt: 812; Hoek : 41; В., 1940 б : 20; O-n : 120. Faren — O-n : 120. [16] норв. Brase — Hoek : 41. Brasen, Brasme — O-n : 120. [18] англ. Bream — Smitt: 812; Hoek : 41; Канада — Ricker : 60. Carpbream — Hoek : 41. [19] нем. Bläuer, Blei сев.-нем., Bleier — Benecke : 119. Bleiha, Bleihe, Bleikja — В., 1948 : 63. Brachsen — Hoek : 41. Brassen южнонем., Brechsen, Bressem, Halbfisch, Reissfisch, Schlaffke — Benecke : 119. ~ Blehe, Plotze — В., 1948 : 63. [20] голланд. Brasem — Hoek : 41. [22] франц. Breme commune — Hoek: 41. [23] итал. Abramide — Yasiliu : 193. [26] рум. Cirjanca, Cirjencuta, Foaia plopului, Frunza plopului, Lapita, Latifa, Plataica, Platica, Platica de butoi, Platica de Dunare, Platica riioasa, Platica ruginoasa, Sinet, Sinet — Yasiliu : 193. [33] венг. Dever, Keszag — Yasiliu : 193. [36] финск. Lahna — Hoek : 41; В., 19406 : 20; Brofeldt: 157. Sulkava — Пром. p. :418. [37] карел. Лахну, Лахнализу (мелк.) Крошнозеро, Парули (мелк.) Коткозеро — A. Lahna — Паллон : 5. ~ Лагна — Сл. зап. XYII в. [38] эст. Lasna — Б. : 768. Latik — В., 19406 : 20. [41] коми Йоды Печора, Кама — Ревн. : 217. Jоды р. Летка — Дрягин, 1927:136. Ляпа (мелк.) Печора, Кама — Ревн.: 217. ~ Jodi — Pallas : 325. [43] марийск. Лавал pp. Кама, Уфа, Ай, Ловал pp. Кама, Уфа — Ревн.: 217. Олама, Оланг, Сюба, Хурбан — Гр. : 147. [45] ненецк. Лысу Печора — Ревн.: 217. [47] азерб. Чапах, Чипах — Барач, 1941 : 219. [48] тат. (казан.) Кара-канат (мелк.), Курбан — Рузский, 1887 : 53. Курбан-балык, Чабар —- Ревн. : 217. Чебак, Чипах — Гр. : 147. ~ Кurban-balyk, Tschabak — Pallas : 325. [53] казах. Тран р. Чу — Г. Ник., 1931 : 257. [54] калм. Зыбы, Цууба, Цюба, Чыбе — Гр. : 147. ~ Tschybe, Zyybi — Pallas : 325. [58] чуваш. Супах — Б., 1948:71. Сюба, Хурбан (мелк.) — Варп., 1886а : 33.
12.1.2 (2). Abramis brama orientalis Berg, 1949 — Аральский лещ (Б. : 774).
[1] русск. Аральский лещ — Г. Ник., 1940 : 126. Беляк н. т. Волги, Касп. м. — Пром. р. : 407. Восточный лещ — Б. : 774. Лещ — Линдб., 1947 : 37; среднеаз. — Б. : 774. Серуха южн. часть Касп. м. — Кессл., 1864 : 115. Чабак Арал. м. — Пром. р. : 410. [18] англ. Aral bream Канада — Ricker : 60. [47] азерб. Чапаг — Абдур. : 241. [53] казах. Тран Арал, м., Чебак Арал. м. — Турд. : 90. [57] каракалп. Чабак — Б. : 774.
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
Лещ (Abramis brama) — ценная
промысловая рыба, распространенная шире других
видов этого рода. На севере он достигает
бассейна Белого моря и восточной части
Баренцева моря (р. Печора),
акклиматизирован в водоемах Сибири (оз. Убинское,
р. Обь), Казахстана (оз. Балхаш и др.).
Лещ предпочитает спокойную теплую воду
с песчано-иловатым и глинистым дном
и поэтому обычен в заливах рек, в озерах.
В нижнем течении Днепра, Дона, Волги
и в северной части Аральского моря лещ
образует две формы — жилую и
полупроходную. Полупроходные формы кормятся
в море, а для икрометания поднимаются
в низовья рек. Особенно многочисленны
стада донского и аральского леща. В
дельте Аральского моря встречается третья
форма — мелкий камышовый лещ,
имеется он и в дельте р. Или.
Окраска меняется в зависимости от
возраста рыбы, цвета грунта и воды в
водоеме. Мелкий лещ серо-серебристый, в
старшем возрасте темнеет и приобретает
золотистый отлив. В торфяных озерах лещ имеет
бурый цвет. Все плавники у леща серые,
в анальном плавнике 23—30 ветвистых
лучей. Лещ достигает длины 45 см, веса
2,5—3 кг; живет до 20 лет, но обычно
меньше. В южных районах растет он
быстро, половой зрелости достигает в 3—4 года, при длине около 25 см. В северных
озерах и водохранилищах половозрелым
становится в 5—8 лет, при длине 30 см и
более, камышовый лещ созревает в 3 года,
имея длину 12—15 см.
Нерест леща начинается при
температуре воды 12—16°С, в конце апреля — в мае
на юге, в конце мая — в июне на севере.
Полупроходной лещ из моря поднимается
для нереста в реки и откладывает икру
на свежезалитую паводковыми водами
растительность так называемых займищ
на Дону, полоев на Волге.
Самцы леща созревают раньше самок,
и более мелкие рыбы на нерестилищах,
как правило, самцы, кроме того, их легко
отличить по хорошо развитой «жемчужной
сыпи» на голове и боках тела. Нерест леща
происходит с шумными всплесками,
обычно рано утром на мелководье среди
зарослей подводной или свежезалитой
растительности. Лещи выскакивают из воды
и падают плашмя в воду, шум
нерестящегося леща слышен на далекое расстояние.
Это шлепанье в воду («лескание» или
«лясканье») послужило поводом для названия
рыбы «лещ» или «лящ» (а в Словакии —
«плескач»). Нерест начинается и
заканчивается дружно, чаще он длится один
день, с утра до полудня.
В бассейнах Балтийского, Каспийского,
Азовского, Черного морей жилой и
проходной лещ мечут икру одновременно,
лещ Аральского моря, в Фархадском
водохранилище мечет икру порционно.
Самка выметывает в среднем 100—150
тыс. икринок, прикрепляющихся к
растительности. В озерах и водохранилищах
на нерестилище леща держатся мелкие
неполовозрелые лещи, плотва, возможно
частью подбирающие икру. Во многих
озерах наблюдается несколько подходов
леща на нерест. Лещи разных подходов
могут иметь специальные названия,
отражающие связь между временем подхода
лещей на нерест и фенологическими
явлениями; например, лещ-дубняк мечет икру,
когда распускаются листья дуба. Из
отложенной икры через 3—6 суток
выклевываются личинки. Первые двое суток они
пребывают в стадии покоя, прикрепившись
к водным растениям, затем приступают
к активному питанию. Сначала пищей им
служит зоопланктон, но после
достижения 30 мм молодь переходит на питание
бентическими беспозвоночными. В
солоноватых участках южных морей лещ
питается ракообразными: мизидами, кумовыми
рачками, в питании леща Азовского моря,
кроме того, большое значение имеют полихеты. Пищей леща в Аральском море
являются бокоплавы, личинки хирономид,
моллюски. В озерах лещ питается
личинками хирономид, ручейников и других
насекомых, моллюсками горошинками.
В озерах, водохранилищах сроки
нереста леща близки к срокам нереста других
рыб, и поэтому нередко появляются
гибриды. Лучше других изучен гибрид леща
и сибирской плотвы из озера Убинского.
Гибрид растет быстрее плотвы, но
медленнее леща. Половой зрелости самцы
достигают на 3-м году, а самки — на 4-м году
жизни. В развитии гонад часто
замечаются различные отклонения: неравномерное
развитие правой и левой гонады или
отдельных участков гонад.
Численность леща в различных
водоемах сильно колеблется и зависит прежде
всего от успешности нереста.
Благоприятным условием для нереста леща в южных
морях является высокий паводок. После
зарегулирования стока рек южных морей
нерестовые площади для леща сильно
сократились. Для сохранения запасов
леща на высоком уровне созданы
специальные нерестово-выростные хозяйства,
проводятся работы по спасению молоди из
отшнуровавшихся и потерявших связь
с рекой мелких водоемов.
В водохранилищах многочисленные
поколения леща появляются в многоводные
годы. Для обеспечения нереста леща в
водохранилищах применяют плавучие
нерестилища. На численность леща в
водохранилищах большое влияние оказывают
также эпизоотии, и прежде всего
заболевание лигулозом.
Благодаря высоким пищевым
качествам лещ имеет важное промысловое
значение.
Ужение леща считается нелегким делом.
Надо хорошо знать особенности
поведения леща, которое очень различно: в одних
водоемах он постоянно обитает в чаще
водных растений и питается только ночью,
в других придерживается кромки водных
растений и кормится на утренних и
вечерних зорях, в третьих предпочитает
глубокие ямы около коряг, завалов,
обрывистых берегов или вблизи каменистых
или песчаных отмелей. На глубоких
местах лещ питается в течение всего дня, а
на мелководья выходит на зорях, играет
у поверхности воды, высовываясь из нее,
или бесшумно «разводит круги». Перед
ловлей на удочку леща обычно
приваживают той насадкой, какую рыболов
выбрал для ужения: пшенной кашей,
дождевым червем, горохом и т. д.
Рыбы пресных вод СССР и сопредельных стран. Л.С.Берг 1948 г.
1. Abramis brama (Linne). — Лещ
(Молодые подлещик; на юге лящ, на Дону чебак, мелкий лещ на Азовском море ласкирь, немцы Brachsen, Blei, финиы lahna, карелы лахна, эсты lasna (родственно русскому лящ).)
Cyprinus brama Linne, Syst. nat., ed. X, 1758, p. 326 (Европа; тип из Швеции).
Abramis brama Heckel und Кner, Süsswassf. Oestr., 1858, p. 104, фиг. 54, 55 (Дунай, Рейн, Эльба, Одер, Висла, Нева, Финляндия). — Кесслер, Рыбы СПб. губ., 1864, стр. 90. — Fatiо, Poissons Suisse, I; 1882, p. 328 (Швейцария только в бассейне Рейна; l.l. 50—55, позвонков 43—45). — Smitt, Scandin. fish., II, 1895, p. 812, табл. XXXIV, фиг. 2 (Швеция). — М. von zur Мühlеn, Arch. f. Naturkunde d. Ostbaltikums, XIV, № 2, Dorpat, 1920, p. 5—16 (биология). — Солдатов, Тр. Плов. морск. инст., вып. 3, M., 1923, стр. 16 (устье Сев. Двины в солоноватой воде, единично в Печоре). — Домрачев и Правдии, Рыбы Ильменя и Волхова, 1926, стр. 52, табл. II—IV (рост, возраст, нерест, питание). — Егерман, Тр. Черн.-азовск. опытн. ст., II, вып. 1, 1926, стр. 35—51 (Кучурганский лиман Днестра; рост, питание, измерения). — Домрачев, Изв. Отд. прикл. ихтиол., IX, вып. 3, 1929, стр. 309 (Псковский водоем; питание, рост). — Маркун, там же, X, вып. 2, 1929, стр. 126 (Псковский водоем; рост). — Паллон, Рыбы Сегозера, 1929, стр. 5, 30 (Сегозеро). — Сыроватский, Тр. Ихтиол. опытн. ст. Херсон, IV, вып. 2, 1929, стр. 102 (дельта Днепра). — Егерман, там же, V, вып. 1, 1929, стр. 51 (Днепр ниже порогов). — Недошивин, Тр. Азовско-черном. эксп., IV, 1929, стр. 37 (низовья Дона). — Телегин, там же, стр. 202 (Таганрогский зал.). — Н. Дмитриев, Тр. Азовско-черном. эксп., VI, 1931, 104 стр. (Азовское море, Дон, Кубань). — Овчинник, Всеукр. Акад. Наук, Тр. природ.-техн. вiдд., № 31, 1931, стр. 95 (пороги Днепра; возраст, рост). — Берг, Рыбы пресных вод СССР, I, 1932, стр. 497 (частью), рис. 437. — Гриб и Вернидуб, Учен. зап. Ленингр. унив., I, биол., 1935, стр. 106 (Финский зал. у Петергофа; биометрика). — Прав дин, Тр. Карел. рыбохоз. ст., I, 1935, стр. 28 (оз. Топозеро в бассейне р. Поньгомы, впадающей в Белое море). — В. Воробьев, Тр. Азовско-черном. инст. рыбн. хоз. и океан., XI, 1938, стр. 97 (Азовское море; распределение). — Дрягин, Изв. Инст. озерн. и речн. рыбн. хоз., XXI, 1939, стр. 107—108 (порционное икрометание в Ильмене). — Берг, там же, XXIII, 1940, стр. 20 (Финский зал.; темп роста у Сестрорецка). — Великохатько, Зоол. журн., XX, № 1, 1941, стр. 101 [нижнее течение Днепра, дельта, лиман; D III 9—10; А III (22) 23—28 (29), в среднем 26.2; l.l. (50) 51—53 (60), в среднем 54.1 у 168 экз. позвонков 43—45, в среднем 44 у 16 экз.; биология].
D III 9 (10), А III (23) 24—30, в среднем 27.4 (у 83 экз.); l.l. (50) 51—60, в среднем 55.2 (у 70 экз.); жаберных тычинок 19—24, в среднем 22.5; позвонков (44) 45—46, обычно 45 (у 11 экз.) (все данные для Невской губы, Гриб и Вернидуб, 1935). Для Псковского водоема: А III 23—28, в среднем 25.1 (у 26 экз.); l.l. 52—57 (59), в среднем 54.2 (у 25 экз.); жаберных тычинок (22) 23—25, в среднем 23.6 (у 20 экз.); позвонков (43) 44—45, в среднем 44.1 (у 21 экз.) (М. И. Маркун). Для Ильменя у 30 экз. позвонков 43—45 (П. В. Тюрии). У 50 лещей из бассейна Дона в А в среднем III 25.7 лучей (Н. И. Николюкии). 2-й и 3-й позвонки плотно срослись (П. В. Тюрин). Наибольшая высота тела у взрослых 35—40% длины тела без С; молодые более вытянуты в длину. Рот полунижний, маленький. Грудные плавники немного не доходят до основания брюшных, иногда доходят или даже заходят. Брюшные доходят до анального отверстия или до анального плавника. В среднем в длине тела у лещей (22 самки) из Невской губы длина хвостового стебля 15.1%, антедорсальное расстояние 57.3%, высота D 20.6%. Грудные плавники серые, прочие с черными концами, у старых все плавники делаются черными. У половозрелых самцов тело и плавники покрываются бугорками. Длина до 280—450 мм, но встречаются до 750 мм и весом до 6 кг и более. В июле 1912 г. в Финляндии, в оз. Веси (Vesijärvi), около Lahtis, пойман лещ весом 11.55 кг (Fiskeritidskrift för Finland, 1930, № 3, p. 43)
По данным А. В. Климовой, исследовавшей 25 самцов (длина тела без С 214 мм) и 25 самок (длина тела без С 283 мм) из оз. Яск-ярви в Карелии, в бассейне р. Вуоксы, близ Энсо, ветвистых лучей в А у самцов 25.7, у самок 25.2, l.l. 54.9 и 54.6, жаберных тычинок 25.2 и 22.2, позвонков (у самцов и у самок) 44.5. Наибольшая высота тела в длине тела у самцов 37.5, у самок 37.9.
О. И. Потапова (1949) подробно исследовала 50 лещей из Сямозера (бассейн Онежского озера). D III (8) 9—10, средн. 9.5, А 23.4, l.l. 52.5, жаберных тычинок 17—25, средн. 19.8, позвонков 41—46, наичаще 44. Наибольшая высота тела в длине тела 41.6%. Лещ этот отличается высокотелостью, меньшим числом лучей в А, жаберных тычинок и чешуй в l.l. Средние размеры промыслового сямозерского леща 31—36 см (без С). Наблюдаются три подхода леща к местам нереста — в конце мая, в первой декаде июня и в середине июня.
Европа к востоку от Пиренеев и к северу от Альп. В Швейцарии только в бассейне Рейна. Низовья Роны (E. Blanchard, Poissons des eaux douces de la France, 1866, p. 355 (Рона. y Авиньона). Возможно, однако, что проник в Рону из Рейна по каналу.). Англия, Ирландия. Бассейны Северного, Балтийского, Черного, Каспийского морей. Река Марица, низовья Струмы, оз. Бешик-гель (п-ов Халкидика, бассейн Эгейского моря), р. Пеней в Фессалии. В Финляндии повсюду в бассейне Балтийского моря. В Финском зал. повсеместно, но главным образом в восточной опресненной части.
Есть также в водах, принадлежащих к бассейну Белого моря, например в реках, впадающих в Карельский берег Белого моря, в Топозере, в Керетьозере, в оз. Сегозере (бассейн р. Выг), в Чарандском озере (бассейн р. Онеги), в Сев. Двине от Кубенского оз. (много) до Архангельска (Для р. Сысолы, притока Вычегды, леща указывал еще Лепехин (Дневн. зап., III, 1780, стр. 267).), в реках Кулое (Наблюдался Е. С. Кучиной в озерах бассейна Сояны, притока Кулоя в 1932 г.), Мезени (Доставлен сотрудниками Мезенской экспедиции Института озерного и речного хозяйства П. Г. Данильченко и Крупиным (1932).) и Печоре (Здесь обнаружен Л. И. Васильевым между устьями рек Ижмы и Уса в 1927 г. В низовьях Печоры найден в 1935 г. (Е. С. Кучина).), но далее на восток отсутствует, равно как нет его и во всей Сибири, за исключением верховьев Исети (на восточном склоне Урала), куда пересажен искусственно. (3 X 1924 в Иртыше под Тобольском пойман лещ длиною 276 мм (Б. Чаликов, Бюлл. Общ. изуч. Тобольск. севера, 1927, № 1, стр. 18). — О разведении леща на Урале см.: А. Подлесный, Тр. Урал. отд. Инст. озерн. и речн. рыбн. хоз., I, 1939, стр. 111.) Низовья Риона (оз. Палеостом). В бассейнах Каспийского и Аральского морей заменен подвидом orientalis.
Г. В. Никольский обнаружил близ оз. Лаче (бассейн р. Онеги) много остатков крупных лещей (до 42 см) в стоянке неолитического возраста (около 4000 лет тому назад). (Бюлл. Моск. общ. испыт. прир., биол., XLIV, 1935, стр. 114.)
Предпочитает не очень быстро текущие и стоячие воды. Не избегает и солоноватых вод Балтийского, Черного и Азовксого морей.
Об Abramis leuckarti (помесь леща и плотвы) см. ниже.

Как видим из таблицы, темп роста на юге быстрее, чем на севере. Особенно надо отметить быстрый темп роста леща из дельты р. Урала и из Азовского моря. Весьма быстро растут также лещи в старших возрастах на днепровских порогах.
В Финском зал., в районе Петродворец (Петергоф) — Ломоносов, нерест начинается в конце мая и заканчивается в середине, а иногда в конце июня. Нерест происходит довольно далеко от берега, в зарослях рдеста, который растет на глубинах 0.4—2 м. На зимовку лещ залегает в районе Кронштадта. Повидимому, осеннего хода леща из Финского зал. в Неву не происходит (Гриб и Вернидуб, 1935).
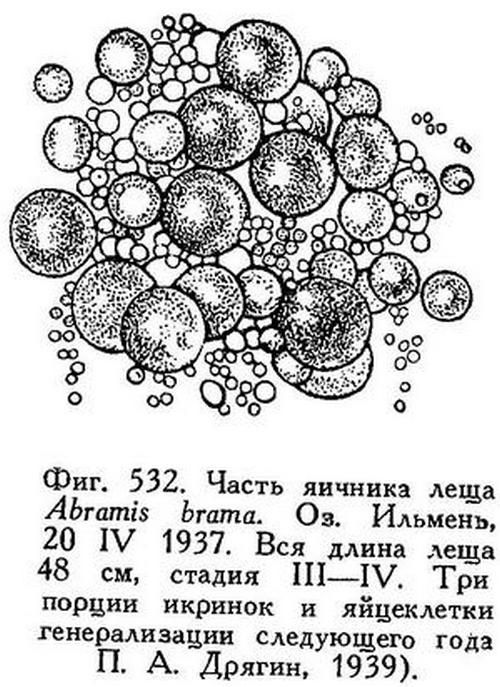
Лещ в Волхове и Ильмене достигает 537 мм (до основания хвостового плавника) при весе в 3.1 кг, но попадаются лещи до 4 и даже, как редкость, до 6 кг. Заходит ли лещ из Ладожского оз. в Волхов, вопрос этот пока не выяснен. Но из Ильменя лещ весною входит массами в притоки и после нереста возвращается обратно в озеро. На зиму лещ залегает в больших количествах как в самом Ильмене, так и в Волхове. Упоминают о зимних уловах в 5 т за одну тоню. Нерест в Ильмене и Волхове происходит с конца апреля до начала июня; он совершается в 3 приема, разделенные промежутками в 7—14 дней; начинается нерест обычно в первой декаде мая; происходит он в озере у берегов, а также в дельтах притоков, особенно в дельте Ловати (Домрачев, 1926). П. А. Дрягин (1939) подробно исследовал порционный нерест леща в Ильмене. По его наблюдениям, нерест происходил в следующие сроки:

У самки, пойманной 20 IV 1937 в Ильмене, имевшей (абс.) длину 48 см и весившей 1.2 кг, вес икры составлял 116 г; в яичнике видны были три порции икринок, которые должны были быть выметаны в том же году, а кроме того, яйцеклетки генерации следующего года (Дрягин, 1939; см. рис. 532). Нерест леща в Ильмене начинается при температуре воды не ниже 12—13°.
В Псковском водоеме лещ питается преимущественно моллюсками, насекомыми (особенно личинками хирономид) и ракообразными (Cladocera, Copepoda). Молодые лещи питаются преимущественно планктоном из Copepoda и Cladocera. Самым обычным объектом питания у взрослых в Псковском водоеме являются личинки хирономид, тогда как в дельте Волги — ракообразные (Домрачев, 1929), в Азовском море — тоже ракообразные (Ostracoda, Дмитриев, 1931).
Ф. Д. Великохатько (1941) различает в низовьях Днепра две формы: озимую и яровую. Озимые лещи крупнее, более темной окраски, более многочисленны; они подымаются из лимана в Днепр не выше чем на 100 км, а дальше сменяются жилою формою, которая не производит миграций. Яровые («беляк») в Днепре встречаются не выше дельты; они хорошо переносят, в отличие от озимых, солоноватую воду и встречаются у Очакова и даже на взморье. Относительно этих форм требуются дальнейшие наблюдения.
Осенняя миграция озимого леща из лимана в Днепр начинается в конце сентября или в октябре и продолжается всю зиму, хотя с похолоданием интенсивность миграции уменьшается. Весною количество подымающегося леща снова увеличивается. Яровой лещ подымается, согласно Ф. Д. Великохатько, только весною, по окончании или под конец нерестовой миграции озимого. Нерест лещей происходит порционно, в 2, а может быть в 3 приема. В 1933 г. нерест озимого начался 1 мая, в 1934 г. около 20 апреля, в 1935 и 1936 гг. около 5 мая, в 1938 г. около 20 апреля. В 1937 г. озимый лещ начал нерестовать 20—25 апреля, а яровой только 6 мая. В это время температура воды достигает 10—12°. Разгар нереста в низовьях Днепра приходится на середину мая. Отнерестившиеся лещи скатываются в лиман, где нагуливаются до осени. Большинство лещей созревают в возрасте почти четырех лет, некоторые, — имея почти 3 года. Развитие половых продуктов от III до V стадии происходит в течение одного месяца. Средняя плодовитость 57 озимых лещей составляет 225 000 (пределы 104—587 тыс.). Средний вес осеннего днепровского леща 600 г; бывают лещи в 2.4 кг, длиною 54 см. Годовики и двухлетки в основной своей массе остаются вплоть до половой зрелости в предустьевой части лимана.
Подобно многим другим азовским рыбам (осетровые, судак, рыбец и др.), и лещ входит в Дон 2 раза в году: весною — с марта по июнь, и осенью — с сентября по ноябрь. Весенний нерестовый ход леща («чебака») в дельте Дона начинается одновременно со вскрытием реки, обычно во второй половине марта и продолжается до 5—15 июня; разгар хода бывает в апреле. Осениай ход гораздо менее интенсивен, хотя продолжается всю зиму. В это время лещ, не подымаясь высоко вверх, залегает на ямах. По наблюдением Н. И. Сыроватской (Докл. Акад. Наук СССР, т. 66, № 5, 1949), у донского леща небольшая часть самок откладывает икру в один прием, большая же часть в два приема. В кубанском районе массовая миграция леща приходится на май.
В 1938 г. месячные уловы леща составили (в %):

Средняя плодовитость донского леща 151 000 икринок (пределы 56—282 тыс.). Нерест леща в дельте Дона происходит с середины мая до середины июня (разгар с конца мая до первой половины июня), в аксайско-донском займище (низовье Дона повыше Ростова), которое является основным на Дону нерестилищем леща, — с середины апреля да конца июня, начала июля. Икра приклеивается к подводной или затопленной мягкой наземной растительности. Массовый скат отнерестившегося леща из Дона в Азовское море происходит в конце мая — первой половине июля. На эти же сроки падает скат малька; покатный малек имеет в длину 25—30 мм. Впервые донской лещ созревает в возрасте 3 лет (главным образом самцы), но массовое созревание наступает в возрасте 4—6 лет.
Общие уловы леща во всем Азовском море таковы (в тысячах центнеров) (Дойников, стр. 30):

Все многоводные годы (1926—1929 и 1932) отличаются большим приплодом леща (Дойников). (К. Дойников, Азовский лещ. Рыбн. хоз., 1939, № 7, стр. 27—32. )
В Кубань лещ входит из Азовского моря в небольшом количестве. Средние размеры леща в Азовском море в 1923 и 1924 гг.:

Средний вес ходового леща в Таганроге в 1923 г. 1.2 кг, в 1924 г. — 0.9 кг, в 1925 г. — 0.94 кг.
Е. В. Кучина приводит данные о лещах Сев. Двины. Оказывается, что по малому числу позвонков двинской лещ приближается к волжскому, но вместе с тем обладает таким же количеством жаберных тычинок на 1-й дуге, как и лещ Финского зал. Возможно, что в Сев. Двине произошло скрещивание местного леща (аборигена) с пришлым, волжским, проникшим сюда через каналы в сравнительно недавнее время. Средние биометрические данные для лещей Сев. Двины, по Е. В. Кучиной, таковы: А ветвистых лучей 25.8, l.l. самцы 55.3, самки 54.2; жаберных тычинок самцы 22.1, самки 22.6.
la. Abramis brama orientalis Berg, nom. nov. — Восточный лещ
(Русские лещ, казахи на Аральском море тран (ср. русск. тарань — наименование не леща), чабак, каракалпаки чабак.)
Abramis brama Кeсслep, Изв. Общ. любит. ест., X, вып. 1, 1872, стр. 68 [Сыр-дарья у Перовска (Кзыл-орды)]. — Берг, Рыбы Туркестана, 1905, стр. 167 (Сыр-дарья вверх до Чиназа, Аму-дарья до Шейх-джели); Ежегодн. Зоол. муз. Акад. Наук, XVII, 1912, стр. 55 (Сары-су у Тохты). — Терещенко, Тр. Астрах. ихтиол. лаб., IV, вып. 2, 1917 (дельта Волги; биология) . — Берг, Изв. Отд. прикл. ихтиол., V, вып. 1, 1926, стр. 72 (Аральское море). — Филатов; Бюлл. Ср.-Аз. унив., № 14, 1926, стр. 219 (дельта Аму-дарьи). — Дуплаков, там же, № 15, 1927, стр. 361—373 (низовья Сыр-дарьи, биология). — Кагановский, Изв. Отд. прикл. ихтиол., V, вып. 2, 1927, стр. 240 (Аральское море: Урга, Муйнак; возраст, рост, половой состав) . — Маркун, там же, VI, вып. 2, 1927, стр. 133 (р. Урал; возраст, темп роста); IX, вып. 1, 1929, стр. 22 [Аральское море, возраст, рост, биометрика 50 самцов и 50 самок; D III 9—10, в среднем 9.1; А III 23—28, в среднем 25.7, l.l. (49) 50—55 (56), в среднем 52.3; жаберных тычинок (20) (21) (22) 23—30, в среднем 26.0, у самцов 26.4, у самок 25.6; Аральское море у Муйнака; тип Abramis brama bergi Grieb et Wernidub]. — Г. Никольский, Тр. Зоол. инст. Акад. Наук, XXXII, 1931, стр.257 (р. Чу в 60 км ниже пересечения Турк.-сиб. ж. д.). — Берг, Рыбы пресных вод СССР, I, 1932, стр. 497 (частью: бассейны Каспийского и Аральского морей); II, 1933, стр. 855—856 (бассейн Каспия). — Беляев, Тр. Азербайдж. рыбохоз. ст., III, вып. 3, 1933, стр. 16 (по Куре поднимается повыше Мингечаура, Кумбашинка, Дивичи). — Державин, Тр. Азербайдж. отд. Закавк. фил. Акад. Наук СССР, зоол., VII, 1934, стр. 108 (Каладагна, Энзелийский, Астрабадский зал.). — Гладков, Тр. Аральск. отд. Инст. морск. рыбн. хоз., III, 1934, стр. 115—121, 142—144 (озера Камышлы-баш, Акча-тау, Каракуль, Раим — в низовьях Сыр-дарьи; размеры, рост, вес) . — Г. Никольский и Панкратова, там же, стр. 172 (пресное Судочье оз. в бассейне Аральского моря — камышовая форма; оз. Тили-куль в бассейне р. Чу). — Панкратова, там же, IV, 1935, стр. 214 (питание в Аральском море).
Abramis brama bergi (nomen praeoccupatum) Гриб и Вернидуб, Учен. зап. Ленингр. унив., I, сер. биол., вып. 1, 1935, стр. 112 (Аральское море у Муйнака по данным Маркуна, 1929; оз. Ясхан на Узбое, 26 экз.: А III 25—30, в среднем 26.6, l.l. 47—55, в среднем 51.1, позвонков у 6 экз. 41—44, обычно 44; р. Сары-су). — Г. Никольский, Рыбы Аральского моря, 1940, стр. 126 (Аму-дарья до Таш-сака, Сыр-дарья до Ферганы).
Abramis brama Лукин, Тр. Общ. ест. Казанск. унив., LVI, вып. 1—2, 1939, стр. 75. (средняя Волга; рост). — Казанова, Изв. Акад. Наук СССР, сер. биол., 1940, № 5, стр. 744 сл. (распределение и миграции молоди в северной части Каспийского моря).
лещ В. Танасийчук, Тр. Волго-касп. рыбохоз. ст., IX, вып. 1, Астрахань, 1947, стр. 115—137 (сев. Каспий; молодь леща, биология) . — Летичевский, там же, стр. 95—114 (нерест в авандельте Волги).
Abramis brama subsp. Г. Шапошникова, Тр. Зоол. инст. Акад. Наук СССР, VIII, 1948, стр. 467 (Волга близ Куйбышева, 60 половозрелых самцов и 60 таких же самок, средняя длина (без С) самцов 335, самок 386 мм; среднее количество позвонков 44.5; ветвистых лучей в А самцы 25.8, самки 26.0; l.l. самцы 53.6, самки 53.3; sp. br. самцы 25.2, самки 25.3; наибольшая высота тела самцы 38.7, самки 39.5%, высота D самцы 25.3, самки 23.9%).
D III 9—10, обычно 9; А III 23—28, в среднем 25.7 (у 321 экз.), l.l. 49—57, наичаще 51—54, в среднем у 307 экз. 52.6; жаберных тычинок на 1-й дуге у 316 экз. (20) (21) 22—30, обычно 23—28, в среднем 25.6 (У 66 других экз. из дельты Аму-дарьи 25—29 (30), в среднем 26.8 (П. Н. Морозова).); позвонков у 94 экз. (42) 43—44, в среднем 43.4 (все данные по экземплярам из Аральского моря, исследованным М. И. Маркуном и П. Н. Морозовой, 1937). Ветвистых лучей в А: Кама у Оханска у 29 экз. (22) 24—28, в среднем 25.5; Урал у Гурьева у 72 экз. (22) 23—28, в среднем 25.6 (М. И. Маркун); Энзели, в среднем 24.9—25.2 (А. М. Шуколюков). — l.l.: Кама у 27 экз. 50—55 (56) (57), в среднем 53.2; Урал у Гурьева у 102 экз. 50—56 (57), в среднем 52.5; Энзели зимний у 109 экз., в среднем 52.3, весенний у 105 экз. 51.1. — Число позвонков: Кама у Оханска у 36 экз. 44.3. — Жаберных тычинок: Кама у Оханска у 32 экз. 22—26, в среднем 23.7; Урал у Гурьева у 52 экз. (20) 21—26, в среднем 24.0.
Этот подвид, типом которого является аральский лещ, описанный под именем Abramis brama bergi, отличается от типичного Abramis brama brama, описанного из бассейна Балтийского моря, главным образом большим количеством жаберных тычинок на 1-й дуге; в среднем у аральских 25—26, у средневолжских 25, тогда как у лещей из Финского зал. 22.5. Кроме того, восточный лещ отличается от типичного меньшим числом позвонков и чешуй в боковой линии.
Пропорции 100 экз. аральских лещей с Муйнака (50 самцов, 50 самок), длиною (до основания С): — самцы в среднем 278 мм, самки 326 мм, по М. И. Маркуну (1929), таковы (в процентах длины тела, до основания С): длина головы 20—25, в среднем 22.1, высота тела 35—44, в среднем 39.2, длина хвостового стебля 11—16, в среднем 13.6, антедорсальное расстояние 54—63, в среднем 58.5, Р—V 20—27, в среднем 23.6, наименьшая высота тела 9—12, в среднем 10.5, длина Р 17—24, в среднем 20.6, высота D 22—30, в среднем 25.5, высота А 15—22, в среднем 18.3, длина А 24—33, в среднем 28.0, диаметр глаза 3—6, в среднем 4.6, длина рыла 5—9, в среднем 7.4, ширина лба 7—9, в среднем 8.0. Глоточные зубы у 92 аральских: у 70 экз. 5—5, у 17 экз. 6—5, у 5 экз. 5—6.
У самок восточного леща из Аральского моря и из Гурьева (р. Урал) несколько меньше жаберных тычинок, чем у самцов (М. И. Маркун, П. Н. Морозова):

Но у леща из средней Волги, по данным Г. X. Шапошниковой, обратно: у 60 взрослых самок в среднем 25.3, у 60 взрослых самцов 25.2. В чем тут дело, должны выяснить будущие исследования.
Бассейны Каспийского и Аральского морей. Волга, сев. Каспий, Урал, Терек, Кура вверх до района Мингечаура, речки ленкоранского побережья (много леща входит в Кумбашинку), реки южного берега Каспийского моря от Энзелийского до Астрабадского зал., низовья Атрека. Озера по Узбою (оз. Ясхан). Аральское море по всем берегам, Аму-дарья по озерам вверх до Таш-сака (выше Турткуля), Сыр-дарья вверх до Ферганы (в озерах), в Чу почти до пересечения Туркестано-сибирской ж. д., в Сары-су от оз. Тили-куль вверх по крайней мере до ур. Тохты. В Зеравшане и Кашка-дарье отсутствует.
По данным Н. А. Остроумова (1948), лещ Печоры по морфологическим признакам ближе к лещу из Аральского моря. Очевидно, он проник сюда из бассейна Камы, подобно здешней плотве, которая, по данным Н. А. Остроумова, относится к Rutilus rutilus fluviatilis.
В Каспийско-волжском районе, т. е. в низовьях Волги и в северной части Каспия, выловлено, по оффициальным данным, в 1902 г. 612 млн. шт. леща, а в 1914 г. лишь 16.1 млн. Но цифры эти малонадежны.
Средние годовые уловы в Волго-каспийском и Уральском районах в центнерах таковы (A. И. Александров, Бюлл. Всекасп. рыбохоз. эксп., 1932, № 5—6, стр. 118.).

Максимальный улов за период с 1898 по 1930 г. приходится на 1930 г., когда в названных районах выловлено 344 900 ц. Средний вес леща сев. Каспия в граммах таков:

Средняя длина в дельте Волги составляла (до основания хвостового плавника, как и ниже; по Киселевичу):

Ход леща в Волгу бывает весною, в марте и апреле; в мае и июне он ослабевает, в июле усиливается, а в августе и сентябре достигает максимума. Весенний ход леща, подымающегося в дельту, не достигает даже с. Петропавловского. Главные нерестилища лежат в нижней части дельты, в местах, поросших водяными растениями, где глубина 25—60 см. В верхней части дельты нерест слабый. Наиболее пригодны для икрометания полой с травянистой или древесной растительностью. Нерест начинается при температуре 17—20°; в нижней части дельты начало приходится на начало мая, максимум на середину мая, конец на начало июня. (Т. Дементьева (Рыбн. промышл. СССР, I, 1945, стр. 24—27) обнаружила на чешуе у северно-каспийского леща нерестовые знаки.) Молодь леща остается на местах вывода около месяца, а в конце июня, в связи с быстрым понижением уровня, начинает скатываться в море и в первой декаде сентября уходит вся. В море лещ распространяется по области, занятой пресной водой с глубинами не свыше 4.5 м. С начала октября молодь начинает группироваться в подводной части дельты. В октябре часть молоди входит в реку и здесь залегает на зиму; главная же масса, повидимому, остается зимовать в предустьевом пространстве. Лещ, входящий в дельту во второй половине лета, залегает здесь начиная с августа на дне реки, «на ямах», иногда совместно с сазаном, судаком и другими рыбами. Весною, в апреле, особенно после морян, лещ подымается с ям и идет на нерестилища; иногда же, в случае сильных морян раннею весною, повидимому, уходит обратно в море. Как правило же, в течение февраля и марта он лежит на ямах. Среди весенних ходовых лещей преобладают двухгодовалые (собственно 2 года 10—11 месяцев), затем идут трехгодовалые, четырехгодовалых мало; годовиков столько же, сколько 4-годовалых. Осенью и на ямах приблизительно такой же состав. Шести- и семигодовалые попадаются единичными экземплярами. Из более чем 100 000 рыб найдено 2 экз. в возрасте 13 лет 4 месяцев и 13 лет 10 месяцев. Без сомнения, лещ мог бы дожить и до большего возраста (см. ниже). Половой состав леща на ямах таков. Исследование свыше 170 000 экз. показало, что здесь в среднем 53% самок и 47% самцов. В отдельных случаях преобладали самцы; так, 21 (8) III 1914 в улове из 28 300 лещей 48% было самок и 52% самцов. На тонях средний результат получился тот же, что и на ямах. Среди молодых лещей, годовиков и двухгодовалых, т. е. среди поколений, еще не затронутых ловом, число самцов и самок почти одинаково, но с возрастом замечается все большее преобладание самок, — очевидно потому, что самцы вылавливаются в большом количестве на нерестилищах. Как самки, так и самцы впервые созревают на 3-м году. В середине августа у двухгодовалых, т. е. на 3-м году, начинает закладываться икра и молоки, и в следующем году весною, в исходе 3-го года жизни, происходит нерест. От 25 до 40% особей нерестятся на 4-м году. Обычно более крупные экземпляры созревают раньше, но это бывает далеко не всегда, и, например, в одной серии уловов наименьшая половозрелая самка имела 210 мм, а наибольшая неполовозрелая 298 мм. В качестве исключения была найдена одна половозрелая самка длиной 164 мм, обычная же минимальная длина половозрелой самки 195 мм. После первого нереста не все лещи нерестуют каждый год. Об этом можно судить по тому, что во всех уловах перед самым процессом икрометания среди трехлетних лещей и старше попадаются рыбы незрелые, которые в данном году нереститься не будут. У самцов старше 8 лет и у самок старше 12 лет половые органы находятся в стадии дегенерации. Плодовитость леща в дельте Волги такова: в возрасте 2 лет 11 мес. в среднем 81 400 икринок, в возрасте 8 лет 11 мес. 442 300. Максимум наблюдался в 941 000. Наибольшее количество икры по весу производят самки на 5—7-м году жизни; у них икра достигает иногда свыше 12% живого веса тела. Диаметр совершенно зрелой икринки 1.0—1.5 мм. Величина зрелых икринок увеличивается с возрастом. Возрастной состав косяков леща, входящего в дельту Волги, по данным подсчета 172 000 шт. в 1913—1915 гг. (с сентября по апрель), таков:

Таким образом, главную массу леща, около 65%, составляют особи в возрасте 2 лет 5 мес. — 2 лет 10 мес. Данные о каспийско-волжском леще из монографии Терещенко, 1917.
Возрастной состав в дельте Волги за 1930—1931 гг. (А. Александров, 1932, стр. 119):

Исследованные М. В. Логашевым в Волге у Тетюш 2 леща имели:

На ср. Волге (в районе Самарской луки) лещ, по данным Г. X. Шапошниковой (1948), начинает входить в пойму вскоре после окончания ледохода, когда значительная часть поймы уже залита паводковыми водами. Нерест леща обыкновенно совпадает с наивысшим подъемом воды. В 1940 г. нерест здесь начался 10 мая, разгар нереста пришелся на 12—13 мая при температуре воды в 13—16°. К 20 мая нерест закончился. Повидимому, порционного икрометания леща на ср. Волге не наблюдается (Шапошникова). Нерестилища леща расположены в наиболее удаленной, поздно заливаемой части поймы; нерест обычно происходит среди кустов шиповника или тальника; икра откладывается на сухую растительность (стебли осоки и т. п.). Самцы в основной массе нерестятся в возрасте 6—7 лет, самки в возрасте 8—9 лет. Самые старые самки были в возрасте 16 лет (1939 г.). Плодовитость средневолжского леща в возрасте от 6 до 16 лет составляет от 40.5 до 653.9 тыс. икринок (73 экз.).
По наблюдениям А. В. Лукина, на ср. Волге, в засушливые годы 4-го десятилетия XX в. произошло сильное заиление тетюшского плеса, и очень возросла кормность участка. От всех этих причин создались благоприятные условия для жизни и размножения леща.
Главный ход леща (озимого?) в Куру происходит в декабре — январе. Он мечет икру в придаточной системе нижнего и среднего течения Куры вверх до Мингечаура. Созревает здешний лещ на 4-м, частью на 5-м году. В 1937 г. в Куре добыто 379 000 шт. леща весом 2.4 тыс. ц. Средняя длина куринского леща 311 мм (до основания С?), вес 633 г (Державин, 1939. Изв. Азербайдж. рыбхоз. ст., вып. 2, Баку, 1939, стр. 21.). Но в Кумбашинку лещ (яровой?) входит в больших количествах весною — во второй половине марта и в первой половине апреля, т. е. в иные сроки, чем в Куру. Средний годовой улов леща в тысячах штук в 1924—1928 гг. в пределах Азербайджана (Беляев, 1933): Кура — 303, Кумбаша и Ольховское оз. — 433, море — 350.
Средние размеры (до основания С) в Азербайджане, по данным 1931 г. (М. И. Федорова, Бюлл. Всекасп. рыбохоз. эксп., 1932, № 5—6, стр. 125.), таковы:

По данным А. М. Шуколюкова (1936), промысел, леща в юго-западном углу Каспийского моря основан как на весенних, так и на зимних подходах. При этом весенние и зимние лещи отличаются как биологически, так и морфологически: рыло у весенних длиннее, голова выше, тело ниже (его высота составляет 33.6% длины тела, тогда как у зимних 36.0%), D и А ниже, l.l. у 105 весенних (30 III 1935) в среднем 53.1, у 109 зимних (8—10 XII 1934) такой же средней величины (30 см) 52.3. Весенний лещ заходит в залив Мурдаб лишь для нереста, тогда как зимний залегает здесь на ямы. Число лучей в D и А не показывает различия между весенним и зимним: у 214 экз. D III (8) 9 (10), А III 22—28, в среднем 25.2. Данные эти весьма замечательны и их следует сопоставить с тем, что выше сообщалось о яровых и озимых формах у днепровского леща, где, однако, морфологических различий не обнаружено.
Аральский лещ мечет икру в пресноводных разливах перед устьями рек, но также, по данным П. Н. Морозовой, вдоль восточного берега Арала между дельтами Аму и Сыра и в заливах о. Возрождения. Икра откладывается на подводную растительность (где ее усиленно поедают окунь и красноперка). Перед устьями Аму-дарьи нерест происходит с середины апреля до середины июня (а по Филатову, до конца июня и даже в июле). В 1925 г. лещ метал на Урге с начала мая и до середины июня. Осенью, около времени замерзания, лещ массами начинает входить в реки, но высоко не поднимается; весною вошедший в реки лещ мечет икру по разливам. В море лещ дважды подходит к берегам: осенью — с октября по январь и весною — с марта до июня. Начало весеннего хода леща в южной части Аральского моря приходится на конец марта (перед вскрытием моря), второй, самый мощный, подход происходит в конце апреля, наконец, третий подход — в половине мая. В северо-восточной части подходы начинаются позже. Осенний подход леща к берегам выражен менее четко. Ходовой лещ представлен 4—6-летками. Половозрелости достигает почти четырех лет. Предельный: возраст 12—13 лет.
Аральские лещи весеннего улова 1925 г. на Муйнаке весили до 1310 кг в тысяче. В 1913 г. средний вес тысячи аральских мерных лещей был 573 кг, в 1899 г. 442—491 кг (в дельте Сыр-дарьи). Средняя длина лещей на Бугуни в 1939 г.: неводных 274 (5580 шт.), сетных 285 мм (2752 шт.) (до основания С). Средний вес аральского леща из неводных уловов за 1936—1942 гг. для самцов 480 г, для самок 605 г.
В дельте Сыр-дарьи массовый нерест леща происходит в мае, но отдельные нерестящиеся особи встречаются до конца августа. По описанию Дуплакова, в 1921 г. первый косяк, подходивший к предустьевому пространству Сыр-дарьи перед вскрытием, начал нерест в зал. Куйлюс 4 мая и закончил 13-го; выметав икру в тростниках, лещи тотчас ушли в открытое море, и с 14 по 16 мая как в заливе, так и в море у берегов лещей почти не было. 17 мая подошел новый косяк; он метал икру с 18 по 25 мая, а затем тотчас ушел в море. «Нерестом двух упомянутых косяков ограничивается массовое икрометание, и в дальнейшем валового подхода рыбы уже не наблюдается. Только отдельные, небольшие косячки, вероятно не превышающие 4—5 тысяч штук, время от времени появляются в заливах и, выметав икру — всегда очень быстро, в течение одного, двух дней, — тотчас уходят обратно в море. Такая картина наблюдается в июне, июле и даже в августе. В 1920 году лещи с совершенно зрелой текучей икрой последний раз были замечены, в Куйлюсе 29 августа». Молодь лещей примерно через месяц после вылупления из икры уходит от берегов в глубину. Во время нереста лещи ничем не питаются, не едят они ничего и во время осеннего хода в реку. У аральских лещей в зрелых яичниках икра, по наблюдениям П. Н. Морозовой, бывает двух размеров: крупная — диаметром 0.87 мм и мелкая: — 0.58 мм; последней иногда бывает столько же, сколько крупной. Без сомнения, аральский лещ мечет икру повторно. Сроки весеннего хода леща на Аральском море (П. Н. Морозова):

Средний вес леща в 1936 г. был 802—1310 г. Лещ в Аральском море питается преимущественно личинками хирономид (мотылем), затем бокоплавами, моллюсками (основная пища крупных лещей в прибрежной полосе Adacna и Dreissena), остракодами; из растений — диатомовыми. Мальки в течение первых 2—3 месяцев жизни питаются исключительно планктоном (Панкратова). О темпе роста леща в Аральском море см. выше. Весовой прирост, по данным П. Н. Морозовой, в Аральском море таков:

Лещ на Аральском море — одна из важнейших промысловых рыб. Уловы в самом море и в низовьях впадающих в него рек таковы (Г. В. Никольский, 1939):

Максимум уловов, совпадающий с подходом леща к берегам, приходится на севере Аральского моря на май, в южной части — на апрель.
На Аральском море рыбаки различают леща морского и камышового. Последний худой и темный; зиму и лето проводит в камышах, не уходя вглубь (Берг, 1926). Камышовый лещ из оз. Камышлы-баш описан Н. А. Гладковым (1934). Размеры и вес его (до основания С; Гладков, 1934):

Темп роста камышового леща из пресного Судочьего оз. (до основания С. Г. Никольский и В. Панкратова, 1934, стр. 173):

На р. Казак-дарье (в дельте Аму-дарьи) П. Н. Морозова (1947) встретила уродливую форму леща с высоким, как у карася, телом (53% длины тела, равной 259 мм), длина Р, V и высота D больше, чем у нормального леща, А III 27, l.l. 51, жаберных тычинок 28.
Blicca bjoerkna (L.) х Abramis brama (L.). — Помесь густеры с лещом
Abramis brama х Blicca bjoerkna? Рузский, Прил. прот. Общ. ест. Казанск. унив., № 146, 1894 (Казань; глоточные зубы 1.5—5.1).
Abramis brama var. sinegorensis Лукаш, Тр. Вятск. инст. краевед., I, 1925, стр. 29 отт. (р. Кобра, приток Вятки; глоточные зубы 1.5—5, 1.5—5.1, 5—5.1); VI, 1933, стр. 67 (низовья Вятки; глоточные зубы 1.5—5). F. Heincke среди 93 лещей из Киля нашел у одного 5—5.1 (Variabilitat und Bastardbildung bei Cypriniden. Festschrift f. Leuckart, Leipzig, 1892, p. 67, tab. VIII, fig. 5). Knauthe (Forsch.-ber. Stat. Plon, IV, 1395, p. 265) из 400 лещей из Берлина у 8 нашел 1.5—5 (и 5—5.1), у одного 1.5—5.1.
D III 8—9, А III 23—26, l.l. 48—55, жаберных тычинок 20—28, глоточные зубы обычно двурядные. Бассейн Волги, зап. Европа.
Н. И. Николюкин (Докл. Акад. Наук СССР, XXX, № 9, 1941, стр. 859.) производил скрещивание леща и густеры из бассейна верхнего Дона. У 67 экз. (обе реципрокные формы: самка леща х самец густеры и самка густеры х самец леща) преобладали формулы глоточных зубов 1.5—5.1 (у 19 экз.) и 1.5—5 (у 10 экз.), но были и другие комбинации: 5—5.1, 2.5—5.1, 2.6—5.2, 2.5—5.2 (у 2 экз.), 5—5 (у 2 экз.) и т. п.
Rutilus rutilus (L.) х Abramis brama (L.). — Помесь плотвы и леща
(На Волге и Днепре вся-рыба, всякая рыба.)
Abramis leuckarii Heckel und Kner, Süsswasserf. Oestr., 1858, p. 117, фиг. 61 (Дунай).
Abramidopsis leuckarii Siebоld, Süsswasserf. Mitteleur., 1863, p. 134, фиг. 15—16 (Дунай, Рейн, Эльба, Одер, Висла).
Abramis (Abramidopsis) leuckarii Кесслер, Тр. СПб. общ. ест., I, 1870, стр. 259 (Волга).
Rutilus rutilus х Abramis brama Берг, Фауна России, Рыбы, III, вып. 1, 1912, стр. 75, табл. III. — Белинг, Збiри. праць держ. рибн. заповiдника «Конча-Заспа», I, 1928, Киев, стр. 96—100 (бассейн р. Днепра у заповедника Конча—Заспа). — Лукаш, Тр. Вятск. инст. краевед., VI, 1933, стр. 45, 102 (р. Вятка).
D III (8) 9—10 (11), A III—IV (14) 15—20, l.l. 42—54. Глоточные зубы 6—5 или 5—5 или 5—6 или 6—6; изредка на одной стороне, еще реже на двух сторонах, зубы бывают двурядные: 1.6—5 или 6—5.1 или 1.6—6.1. (Н. И. Николюкин, Доклады Акад. Наук, XXX, № 9, 1941, стр. 858. Исследовано 173 экз. помесей (среди них и реципрокные), искусственно цолученных от особей из бассейна верхнего Дона.) Жаберных тычинок 17—20. Киль за брюшными плавниками резко выражен, но покрыт чешуей; иногда, впрочем, в задней части на незначительном протяжении свободен от чешуи. Особи этой помеси достигают половой зрелости. Во время икрометания тело и плавники у самца покрываются бугорками.
Распространена всюду, где водятся плотва и лещ. В низовьях Волги эту помесь дают вобла и лещ, на средней и нижней Волге — серушка и лещ, на Аральском море, где она обычна, — аральская плотва и аральский лещ.
Н. И. Николюкин искусственно получил в бассейне Дона помеси 1) между красноперкой и линем Tinca tinca (Бюлл. Биол. ст. Воронежск. педаг. инст., I, вып. 1,1939, стр. 5—27, рис.), причем делались скрещивания самки красноперки с самцом линя и самца красноперки с самкой линя; помеси в возрасте 5—7 лет достигали 13—18 см абс. длины; 2) между красноперкой и лещом Abramis brama, у 79 экз. зубы ясно зазубренные, обычно двурядные, изредка однорядные; 3) между красноперкой и густерой; у 100 экз. зубы двурядные, зазубренные; 4) между красноперкой и уклеей (Докл. Акад. Наук СССР, XXX, № 9, 1941, стр. 859—860).
В некоторых случаях различия между реципрокными помесями рыб имеют матроклинный характер, причем приурочены только к периоду эмбрионального развития, а затем исчезают. Это отмечено Н. И. Николюкиным (1946) для помеси между красноперкой и линем.