(Clupea Linnaeus, 1758) Род Клупеи, Сельди, Океанические сельди, Морские сельди, Genus Clupea Linnaeus, 1758 (Atlantic herrings, Pacific herrings) 2 вида
Словарь названий пресноводных рыб СССР. Г.У.Линдберг и А.С.Герд 1972 г.
3.2.0. Clupea Linnaeus, 1758 — Океанические сельди.
[1] русск. Селдь — Леке. нач. XVIII в.; Начерт. : 397; Истор. о жив. : 83; Теряев : 52. Селёдка — Росс, целл.; Нордстет; САР; Сл. 1847 г. Сельдь — Грам. тамож. Белоз., 1497 г.; Поликарпов; Торговый устав 1724 г.; Леке. 1762 г.; Росс, целл.; Нордстет; Озерецк., 1791 : 77; Блуменбах : 407; САР; Двиг. : 136; Ловецкий : 488; Сл. 1847 г.
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
Морские сельди (Clupea) населяют
умеренные воды северного полушария (бореальная область) и сопредельные моря
Ледовитого океана, а в южном
полушарии они живут у берегов Чили.
Морские сельди — стайные планктоноядные рыбы, обычно до 33—35 см длины.
Чешуя циклоидная, легко спадающая.
Килевые чешуйки слабо развиты. Бока
и брюшко серебристые, спинка
сине-зеленая или зеленая. Откладывают донную
прилипающую икру на грунт или
водоросли. Большинство морских сельдей
живут вблизи берегов, только немногие расы
в период нагула уходят за пределы
шельфа. Среди морских сельдей имеются как
совершающие дальние миграции с
пассивным расселением личинок и мальков,
возвратными миграциями подрастающих рыб
и нагульно-нерестовыми странствиями
взрослых особей, так и образующие
местные стада, приуроченные к окраинным
морям; имеются также озерные формы,
живущие в полузамкнутых или
совершенно обособленных от моря
солоноватых водоемах.
В настоящее время различают три вида
морских сельдей — атлантическую, или
многопозвонковую, восточную, или
малопозвонковую, и чилийскую сельдь.
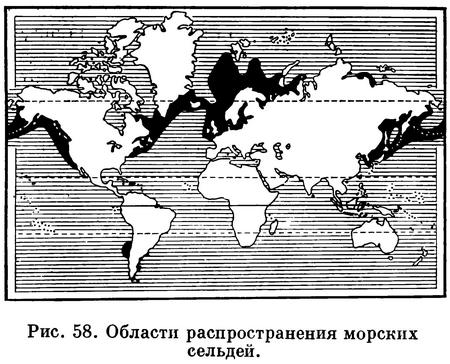
Морские сельди — важнейшая основа
мирового рыболовства: уловы их
составляли в 1960—1967 гг. около 8% общего
мирового улова рыб и беспозвоночных.
Рыбы Японского моря и сопредельных частей Охотского и Желтого морей. Часть 2. (Acipenseriformes — Polynemiformes). Г.У.Линдберг, М.И.Легеза 1965 г.
3. Род CLUPEA LINNE, 1758
Clupea Linne, Syst. Nat., ed. X, 1, 1758 : 317 (тип: Clupea harengus L.). — Световидов, Фауна СССР, Рыбы, 2, 1, 1952 : 115 (описание).
Жировые веки сравнительно хорошо развиты, прикрывают глаза сзади и немного спереди. Брюшные килевые чешуи слабые, хорошо заметные лишь позади брюшных плавников, 24—32 впереди брюшных плавников, 11—18 позади них; брюхо впереди брюшных плавников закругленное, позади них со слабым килем. Сочленение нижней челюсти с черепом и задний конец верхнечелюстной кости под вертикалью середины глаза или у мелких впереди, а у крупных позади нее; нижняя челюсть заметно выдается вперед. Сравнительно слабые зубы есть на сошнике, расположены на продольном гребне головки, несколько зубов на межчелюстной и на переднем конце нижнечелюстной кости, едва развитые на нёбных, иногда на языке, средняя часть нижнего края верхнечелюстной кости с мелкими зубчиками; на мезоптеригоиде зубов нет. Жаберные тычинки верхней части жаберной дуги налегают на тычинки нижней части дуги, тычинки довольно длинные и тонкие, 40—51 на нижней половинке дуги. Брюшные плавники расположены позади начала спинного, обычно под передней третью его, иногда немного далее середины основания; брюшные плавники обычно I 8, иногда I 7 и I 9, как редкое исключение I 6 и даже I 5. Основание спинного плавника лежит в бороздке, образованной расширенными чешуями, более крупными у начала плавника. Позвонков 45—60. Пилорических придатков 18—29. Пятен за жаберной крышкой и на боках тела нет. Икра прилипающая, откладываемая на грунт или подводную растительность, без жировой капли, с малым околожелтковым пространством. Крупного и среднего размеры рыбы, в половозрелом состоянии от 10—15, некоторые более крупные формы от 25—30 см. (Световидов, 1952а : 115).
1 вид.
Рыбы северных морей СССР. А.П.Андрияшев 1954 г.
1. Род CLUPEA LINNE — ОКЕАНИЧЕСКИЕ СЕЛЬДИ
Clupea Linne, Syst. Nat., ed. X, 1758 : 317 (тип: Clupea harengus L.). — Берг, Рыбы пресн. вод СССР, I, 1948 : 147. — Световидов, Сельдевые, Фауна СССР, Рыбы, II, 1, 1952 : 115.
Брюхо закругленное, его килевые чешуйки слабые, их число 36—48. Верхняя челюсть спереди без вырезки, ее задний конец доходит до вертикали середины глаза. Нижняя челюсть выдается вперед, сочленение ее с черепом под серединой глаза. Есть зубы на челюстях, сошнике, нёбных (слабые), иногда на языке; мезоптеригоид без зубов. Крышечная кость гладкая, без радиальных полосок. Спинной плавник закладывается в задней трети тела, но по мере роста малька сдвигается вперед на середину тела. Брюшные плавники под передней третью спинного плавника. Нет больших чешуй (alae) у основания хвостового плавника. Задние лучи анального плавника не удлинены. D (всех лучей) 16—22, А 13—20. Поперечных рядов чешуй 53—70. Позвонков (45) 47—59 (60). Жаберных тычинок около 60—70. Пилорических придатков 18—29.
Икра донная, прилипающая, без жировой капли и с малым около желтковым пространством.
Океанические сельди распадаются на две довольно хорошо обособленные группы: многопозвонковых (атлантических harengus) и малопозвонковых (тихоокеанско-арктических pallasi). В определении таксономического значения этих групп имеются разногласия: одни авторы (П. Шмидт, Берг, Рабинерсон, Световидов и др.) считают их подвидами, но некоторые (Гейнке, ихтиологи США и Канады, Аверинцев, Шнакенбек, Расс, Пономарева и др.) самостоятельными, хотя и близкими видами. Не занимаясь специально систематикой сельдевых, я не считаю возможным входить в детали этого вопроса и следую общепринятой в нашей литературе системе Л. С. Берга, с большой полнотой разработанной и изложенной А. Н. Световидовым, который в своей монографии о сельдевых рыбах критически пересмотрел как морфологию этой группы, так и обширнейшую литературу.
Основные отличия между мурманской и тихоокеанско-арктической сельдью сводятся к следующему.
1 (2). Позвонков обычно 56—58. Килевые чешуйки более или менее развиты как позади (чаще 14—15), так и впереди брюшных плавников. Зубы на сошнике сравнительно хорошо развиты. Икра откладывается на глубинах на грунт. — Баренцово море и южнее; заходит в Белое море. 1а. Clupea harengus harengus L.
2 (1). Позвонков обычно 52—55. Килевые чешуйки более или менее развиты только позади брюшных плавников (чаще их 11—12). Зубы на сошнике развиты слабее, менее многочисленные. Икра откладывается в прибрежной зоне, обычно на растительность. — Северная часть Тихого океана и арктические моря. 1б. Clupea harengus pallasi Val.
Фауна СССР. Рыбы. Том II, вып.1. Сельдевые (Clupeidae). А.Н.Световидов 1952 г.
Род 3. CLUPEA LINNE — МОРСКИЕ, ИЛИ ОКЕАНИЧЕСКИЕ СЕЛЬДИ
— Clupea Linnaeus, Syst. nat., ed. X, 1758 : 317 (тип: Clupea harengus).
— Rogenia Valenciennes, in: Cuvieret Valenciennes, Hist. nat. poiss., XX, 1847 : 340 (тип: Rogenia alba = молодь Clupea harengus).
По отсутствию заметной медиальной вырезки в верхней челюсти, удлиненных чешуй (ala) на лопастях хвостового плавника, удлиненных двух последних лучей анального плавника и кожистого двухлопастного выроста на вертикальной части ключицы и по гладкой крышечной кости без радиальных бороздок подобен Sprattus. Жировые веки сравнительно хорошо развиты, прикрывают глаза сзади и немного спереди. Брюшные килевые чешуи слабые, хорошо заметные лишь позади брюшных плавников, 24—32 впереди брюшных плавников, 11—18 позади них; брюхо впереди брюшных плавников закругленное, со слабым килем позади них. Сочленение нижней челюсти с черепом и задний конец верхнечелюстной кости под вертикалью середины глаза или у мелких впереди и у крупных позади нее, нижняя челюсть заметно выдается вперед. Сравнительно слабые зубы есть на сошнике, расположены на продольном гребне головки, несколько зубов на межчелюстной и на переднем конце нижнечелюстной кости, едва развитые на нёбных, иногда на языке, средняя часть нижнего края верхнечелюстной кости с мелкими зубчиками; на мезоптеригоиде зубов нет. Жаберные тычинки верхней части жаберной дуги налегают на тычинки нижней части дуги, тычинки довольно длинные и тонкие, 40—51 на нижней половине дуги. Отростки плавательного пузыря помещаются в prooticum и pteroticum. Брюшные плавники расположены позади начала спинного, обычно под передней третью его, иногда немного далее середины основания, V обычно I 8, иногда I 7 и I 9, как редкое исключение I 6 и даже I 5 (Иногда при нормальном числе лучей на одной стороне на другой бывает I 9, на одной стороне I 5—на другой I 6 (Heincke, 1898 : 99, Taf. VIII, figs. 1—4).). Основание спинного плавника лежит в бороздке, образованной расширенными чешуями, более крупными у начала плавника. Позвонков 45—60. Пилорических придатков 18—29 (Как очень редкое исключение бывает 6 и даже 4 (Heincke, 1898, Taf. VIII, figs. 13—14).). Пятен за жаберной крышкой и на боках тела нет. Икра прилипающая, откладываемая на грунт или подводную растительность, без жировой капли, с малым околожелтковым пространством. Крупного и среднего размера рыбы, в половозрелом состоянии от 10—15, некоторые более крупные формы от 25—30 см.
Скелет. (Из работ, касающихся описания скелета головы, следует отметить работы Мэтьюза (Matthews, 1887 : 274—287, pis. XVII—XVIII, figs. 18—25), Смитта (Smitt, 1895 : 947—951, figs. 237—238), Райдвуда (Ridewood, 1904a : 453—459, fig. 125).) Bulla prootica немного крупнее, чем bulla pterotica. Bulla prootica образует на черепе сбоку большое и резко выпуклое вздутие, вздутие bulla pterotica сбоку небольшое, несколько большее сверху под fossa praeepiotica. Отростки плавательного пузыря на occipitalia lateralia помещаются в костном канале, образующем с каждой стороны черепа небольшое продолговатое вздутие между fenestra auditiva и отверстием n. vagus; отверстие, через которое отростки плавательного пузыря заходят в череп, расположены у нижнего края occipitalia lateralia. Fenestra auditiva небольшая, несколько продолговатая, немного более широкая сзади, чем спереди, передний край ее отделен значительным промежутком от bulla prootica, задний — от отверстия n. vagus. Fossa praeepiotica большая и высокая, края ее образуют равнобедренный, почти равносторонний треугольник, обращенный основанием вверх, углом вниз. Foramen temporale сравнительно небольшое, удлиненное, по длине почти равное длине fossa praeepiotica, но значительно менее высокое. Сочленовная поверхность для hyomandibulare на sphenoticum и prooticum сплошная, почти не разделенная на две части. Парасфеноид слегка изогнутый, крылья его не выдаются вниз под миодомом, широкие как с боков миодома, так и за задним отверстием его; передняя часть парасфеноида сверху с килем, более высокая спереди. Нисходящий отросток базисфеноида выдается вперед из-под prooticum. Задний край supraoccipitale на черепе сверху (Б) расходится от середины под тупым углом, задний край epioticum заостренный или притуплённый. Pteroticum сзади оканчивается небольшим, загнутым немного вниз шипом. Передний конец каждого sphenoticum отходит вперед и в сторону, заострен на конце. Ethmoidalia lateralia слегка выдаются из-под переднего края лобных костей, направлены в стороны и вперед, варьируя в этом отношении. Поперечные отростки мезэтмоида направлены несколько вперед, не заострены на концах. Череп довольно широкий, ширина его между краями pteroticum около 2 раз в длине его, наибольшая ширина между краями лобных костей от 2.5 до 3.5 раз. Сошник на черепе снизу (В) слегка закругленный, с более или менее широким и коротким, равным около 1/3 длины его, гребнем; передняя часть парасфеноида плоская, слегка суживающаяся назад; крылья парасфеноида разделяются и сразу широко расходятся под передним концом prooticum. Opisthoticum сзади оканчивается коротким и тупым отростком. Углубления fossae temporales posteriores на черепе сзади (Г) слабые. Fossae subtemporales глубокие и значительных размеров, овальной формы, вытянуты в горизонтальном направлении, каждая располагается над отростком opisthoticum в pteroticum и occipitale laterale. Головка для сочленения с черепом сверху hyomandibulare одна, почти одинаковой ширины на всем протяжении. Quadratum без выемки в верхнем крае. Нижнечелюстная кость широкая, сочленовная также высокая, короче нижнечелюстной. Описание сделано по многим скелетам Clupea harengus harengus, Clupea harengus pallasi и нескольким Clupea harengus harengus n. membras, Clupea harengus pallasi n. suworowi, Clupea harengus pallasi n. maris-albi.
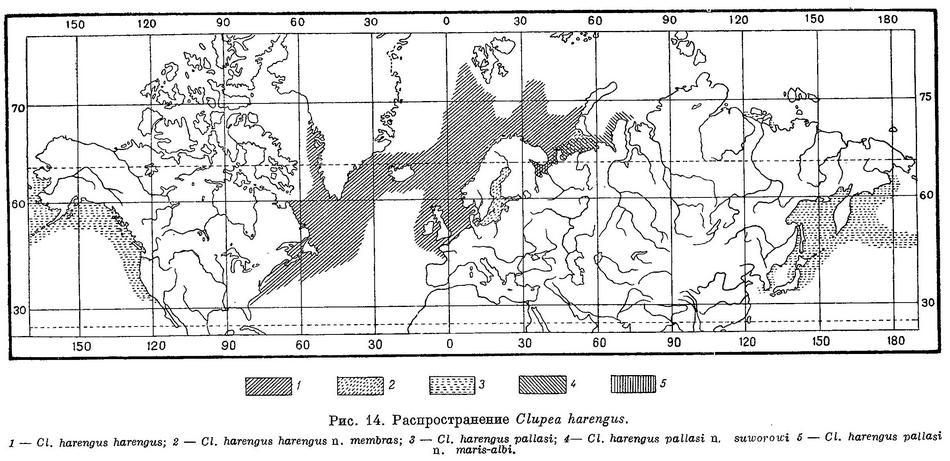
Морские рыбы. Один вид в умеренных частях северного Атлантического и Тихого океанов и в прилегающих как умеренных, так отчасти и в арктических морях (рис. 14). Кроме того, к этому или, возможно, особому, новому роду относится Clupea bentincki Norman, 1936 (размеры 110—115 мм, берега Чили от 37° ю. ш. до Магелланова прол., Гальегос к востоку от входа в этот пролив) и Clupea fuegensis Jenyns, 1842 (размеры 140—170 мм, Магелланов прол., Патагония — Фольклэндские о-ва), предположительно относимые автором (1949 : 47, рис. 4) к роду Sprattus. У Clupea bentincki, в отличие от рода Sprattus, отростки плавательного пузыря помещаются как в prooticum, так и в pteroticum. Кроме того, у этого вида килевые чешуи слабо развиты и брюхо впереди брюшных плавников без острого киля, закругленное; брюшные плавники расположены под или позади вертикали начала спинного; сочленение нижней челюсти с черепом под вертикалью середины глаза. По всем этим признакам Clupea bentincki относится к роду Clupea, но зубов на сошнике у него нет, что однако, не является существенным, так как у одних видов Sprattus зубы на сошнике есть, у других отсутствуют. Наличие или отсутствие зубов на некоторых костях не является существенным и у рода Alosa. От Clupea harengus и всех видов рода Sprattus этот вид отличается большим количеством жаберных тычинок (75—95 на нижней части жаберной дуги, у Clupea harengus — 40—51, у видов Sprattus — 33—41), и, что самое существенное, жаберные тычинки нижней части 1-й жаберной дуги у исследованного экземпляра налегают снаружи на тычинки верхней части дуги, между тем как у Clupea harengus жаберные тычинки верхней части 1-й жаберной дуги налегают на тычинки нижней части дуги, а у Sprattus (Sprattus sprattus, Sprattus arcuatus, Sprattus bassensis, у других видов не установлено) налегания жаберных тычинок одной части жаберной дуги на тычинки другой части дуги нет. По количеству килевых чешуй (18—20 + 10—12) этот вид ближе к Sprattus (19—24 + 8—13), чем к Clupea harengus (24—32 + 11—18). Таким образом, этот вид ничего общего, кроме последнего признака, с родом Sprattus не имеет, существенно отличаясь налеганием жаберных тычинок и от рода Clupea. У Clupea fuegensis, отсутствующего в наших коллекциях, наличие или отсутствие bulla pterotica установить не представилось возможным, равным образом также и характера налегания жаберных тычинок одной части жаберной дуги на тычинки другой части ее. Килевые чешуи у этого вида также слабо развиты и не образуют впереди брюшных плавников острого киля; брюшные плавники расположены под или позади вертикали переднего конца спинного плавника, т. е. по всем этим признакам он подобен предыдущему виду и Clupea harengus, однако зубы на сошнике, как и у Clupea bentincki, отсутствуют, по числу килевых чешуй (22—25 + 10—13) он ближе к Sprattus, жаберных тычинок на нижней части жаберной дуги меньше (38—42), чем у Clupea bentincki. До более детального изучения строения скелета и других особенностей, в частности икринок (пелагические они или донные, приклеивающиеся) и числа пилорических придатков, трудно сказать, относятся ли оба эти вида к роду Clupea или к особому роду.
Clupea совершает в общем меньшие по протяженности миграции, чем Sardinops, Sardina и другие роды, расположенные в середине ряда. Clupea harengus pallasi, Clupea harengus pallasi n. suworowi, Clupea harengus pallasi n. maris-albi и Clupea harengus membras живут и размножаются ближе к берегам, чем Clupea harengus harengus. Последний размножается на больших глубинах, часто в отдалении от берегов, на банках среди моря. Среди локальных форм Clupea harengus harengus есть такие, которые совершают значительные миграции (например мурманская сельдь), но у многих форм, в частности у некоторых в Северном море, миграции ограниченные.
Однако в пределах наиболее изученных родов и видов у более примитивных по рассматриваемым признакам зубы развиты сильнее. В частности, из подвидов Clupea harengus зубы на сошнике сильнее всего развиты у Clupea harengus harengus, более примитивного по положению плавников и по другим признакам, чем Clupea harengus pallasi и его формы (Clupea harengus pallasi n. maris-albi, Clupea harengus pallasi n. suworowi), у которых зубы на сошнике развиты менее.
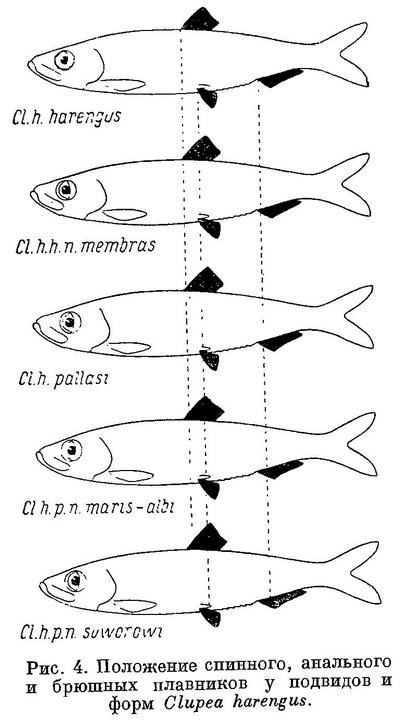
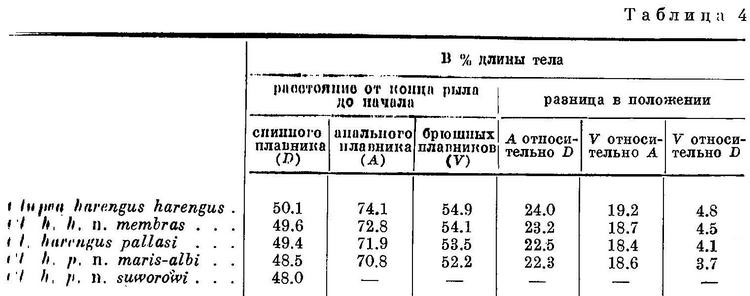
Изменение положения плавников у Clupea harengus, имеющего наибольшее количество подвидов, представлено в табл. 4 и на рис. 4. Следует отметить, что положение анального плавника у подвидов Clupea harengus изменяется в том же направлении, что и спинного и брюшных. Однако вследствие того, что на протяжении ряда положение анального плавника относительно переднего конца тела изменяется в большей степени, чем спинного и брюшных, разница в положении анального плавника относительно спинного также уменьшается, равно как и брюшных относительно анального плавника.
*1. Clupea harengus L. — Морская, или океаническая сельдь.
Литературу см. при подвидах.
Зубы на сошнике есть. Sp. br. 58—73, на нижней части дуги 40—51.
Морской вид, но некоторые формы размножаются и отчасти живут в солоноватой воде.
Разделяется на два подвида, которые в свою очередь распадаются на ряд форм, причем подвиды и их формы по количеству позвонков, степени развития зубов и ряду биологических особенностей разделяются на две группы — многопозвонковых и малопозвонковых сельдей. К многопозвонковым сельдям относятся атлантическая (Clupea harengus harengus) и балтийская (Clupea harengus harengus n. membras) сельди, к малопозвонковым — тихоокеанская (Clupea harengus pallasi), чёшско-печорская (Clupea harengus pallasi n. suworowi) и беломорская (Clupea harengus pallasi n. maris-albi) сельди. Биологические различия обеих групп сводятся к следующему. 1) У многопозвонковых сельдей икрометание происходит на протяжении всего года, в связи с чем среди них, в частности у атлантической сельди, по времени нереста различают весенних, осенних, летних и зимних сельдей. Малопозвонковые сельди размножаются весной, и поэтому у них по времени икрометания есть только весенние сельди. 2) Икрометание у многопозвонковых сельдей происходит в общем при несколько более высокой температуре, чем у малопозвонковых. 3) Нерестилища многопозвонковых сельдей располагаются наичаще на значительном отдалении от берегов, иногда на банках среди моря, и на больших глубинах, чем у малопозвонковых, у некоторых форм атлантической сельди на глубине до 200 м. Нерест малопозвонковых сельдей происходит в узко прибрежной зоне на небольших глубинах, максимально до 10—15 м. 4) В связи с этим икра малопозвонковых сельдей, как правило, откладывается на подводную растительность, между тем как у многопозвонковых сельдей икра откладывается на грунт, так как на таких глубинах, где они размножаются, подводной растительности обычно нет. 5) Малопозвонковым сельдям свойственна большая по сравнению с многопозвонковыми приуроченность в период икрометания к опреснениям. 6) Формам малопозвонковых сельдей свойственна большая локальность и привязанность к узким районам распространения и в общем меньшие по размерам миграции, чем многопозвонковым, причем у последних миграции происходят и в личиночной стадии в виде пассивного дрейфа их с течениями (см. об этом «Биология»). Эти биологические различия обеих групп сельдей обусловлены различным происхождением их. Биологические особенности малопозвонковых сельдей выработались в условиях арктических морей, где большинство их (Clupea harengus pallasi n. maris-albi и Clupea harengus pallasi n. suworowi) обитает и в настоящее время. В арктических морях температурные условия, при которых происходит икрометание и развитие икры и личинок Clupea harengus, кратковременны, и, чтобы иметь, соответствующую температуру для развития икры и личинок, размножаться эта сельдь здесь может лишь весной. Поэтому в арктических морях и имеются лишь весенние сельди. По этой же причине и у Clupea harengus harengus в северных частях ареала (у Норвегии, Исландии, склонов континентального плато Северного моря и др.) имеются лишь весенние сельди, размножающиеся весной и отчасти в конце зимы и летом, а осенние свойственны южным частям ареала (Северное море, Ламанш и др.). Температурными условиями арктических морей обусловлено и икрометание малопозвонковых сельдей на небольшой глубине у самых берегов, так как вода только здесь и прогревается до той температуры, при которой возможны икрометание и развитие икры и личинок. Склонность малопозвонковых сельдей к опреснениям в период икрометания обусловлена тем, что более теплая вода рек и поверхностного стока оказывает отепляющее влияние на прибрежные воды, вследствие чего в арктических морях икрометание частично и происходит вблизиустьев рек. Биологические особенности Clupea harengus pallasi, аналогичные биологическим особенностям других малопозвонковых сельдей, выработались несомненно также в арктических морях. Попав из арктических морей в умеренные части Тихого океана, малопозвонковая сельдь образовала здесь особый подвид, широко расселившийся здесь, но сохранивший все биологические особенности этих сельдей арктических морей. В силу консерватизма своей наследственности этот подвид оказался неспособным дать формы, размножающиеся на больших глубинах в отдалении от берегов и в другое время кроме весны, вследствие чего у него нет и осенних форм (подробнее об этом см.: Световодов, 1949б).
Беломорская и чёшско-печорская сельди приняты Л. С. Бергом за nationes Clupea harengus pallasi, что безусловно правильно отражает их родственное и подчиненное отношение к последней. Балтийская сельдь, имеющая от Clupea harengus harengus такие же отличия, как формы малопозвонковых сельдей от Clupea harengus pallasi, и являющаяся в сущности одной из местных форм атлантической сельди, принята им в качестве подвида; следует отметить, что некоторые из подвидов других видов семейства (например Sprattus sprattus, Sardinops sagax, отчасти Sardina pilchardus и др.) имеют такую же степень различия, что и формы многопозвонковых и малопозвонковых сельдей и поэтому следовало бы считать и формы обеих групп также подвидами, подобно тому как это принято для Gadus morhua, который также разделяется на две группы подвидов, аналогичных группам и формам Clupea harengus и имеющих сходные и биологические особенности, обусловленные их происхождением (Световидов, 1944, 1948). Однако при таком наименовании разделения Clupea harengus на две группы подчиненного значения форм каждой из этих групп не видно. Кроме того, и степень различия форм обоих подвидов Clupea harengus значительно меньше, чем подвидов обоих групп Gadus morhua. Поэтому в данном случае удобнее, несмотря на громоздкость наименования, принять разделение Clupea harengus на два подвида — многопозвонковых (Clupea harengus harengus) и малопозвонковых (Clupea harengus pallasi) сельдей, а формы их за nationes. При этом балтийскую сельдь, принимая во внимание то, что она является такой же формой Clupea harengus harengus, как, например, атлантическо-скандинавские сельди или близкие к ней сельди датских проливов, следует считать за natio атлантической сельди. Резких различий между этими подвидами в основных систематических признаках, так же как и в других, приводимых ниже в диагнозах, нет. Развитие зубов, на сошнике, более сильных и многочисленных у атлантической сельди, имеет также переходный характер, так как одинаковая степень развития встречается у той и другой. Наблюдаемые между многопозвонковыми и малопозвонковыми группами сельдей резкие различия личинок в положении анального отверстия (на 47—49-м сегменте и под 82—83-й клеткой хорды у Clupea harengus harengus и на 42—45-м сегменте и под 62—67-й клеткой хорды у Clupea harengus pallasi n. maris-albi) и в числе клеток хорды (более 100 у Clupea harengus harengus и приблизительно 90 у Clupea harengus pallasi n. maris-albi) обусловлено малым числом исследованных личинок. При исследовании большего числа их и при этом у всех форм как Clupea harengus harengus, так и Clupea harengus pallasi, эти, несомненно варьирующие в значительно более широких пределах признаки, в особенности число клеток хорды, будут иметь заходящий характер. Наоборот, в нижеследующем описании подвидов hiatus имеется в некоторых признаках между атлантической и балтийской сельдями, с одной стороны, и тихоокеанской и чёшско-печорской и беломорской, — с другой. Различия эти обусловлены тем, что характеристика атлантической сельди дана лишь по одной весенней форме (атлантическо-скандинавской), так же как и балтийской (весенняя сельдь). При дальнейшем исследовании обеих этих сельдей, основанном на изучении и других форм их, в частности и осенних той и другой и более мелких по размерам форм атлантической, имеющиеся резкие различия в некоторых признаках (длина головы, размеры глаз) несомненно сгладятся. Равным образом и описание подвидов и малопозвонковых сельдей дано лишь по одной из форм каждой из них: тихоокеанской— по южносахалинско-хоккайдской, беломорской — по мелкой сельди Онежского зал., а чёшско-печорской — вообще по недостаточному числу экземпляров.Резкие различия и между этими подвидами наблюдаются в тех же признаках, что и между атлантической и балтийской сельдями, и в обоих случаях обусловлены, помимо указанных причин, в значительной степени и разными размерами их (более крупными атлантической и тихоокеанской сельдей и более мелкими остальных).
Когда работа была закончена и сдана в Издательство, автор познакомился со статьей Пономаревой (1951), цитируемой при описании Clupea harengus harengus и Clupea harengus pallasi. В этой статье Л. А. Пономарева рассматривает многопозвонковых и малопозвонковых сельдей за самостоятельные виды — Clupea harengus и Clupea pallasi, а все формы — за подвиды: Clupea harengus membras, Clupea pallasi maris-albi и Clupea pallasi suworowi. Однако все приводимые ею в пользу этого доводы основаны на малой компетенции в рассматриваемом вопросе и на недостаточном знакомстве с литературой. Л. А. Пономарева, считая число позвонков важнейшим признаком, положенным в основу систематики рода Clupea, утверждает, что «по этому признаку многопозвонковые или океанические и малопозвонковые или восточные сельди всегда резко различны» и в качестве «наглядной иллюстрации» приводит кривые числа позвонков (рис. 3). Однако даже на этом рисунке кривые числа позвонков атлантической сельди и малопозвонковых — тихоокеанской и ее форм явно трансгрессивны. Нельзя не отметить несоответствия между количеством позвонков на этих кривых и в описаниях. Так, у атлантической сельди по кривой позвонков 55—59, а в описании 53—55, у чёшской сельди по кривой позвонков 52—56, в описании 54, в среднем: 53.5 и т. д. Если же принять во внимание пределы колебаний количества позвонков, известное по литературным данным, то в действительности мы имеем дело не с трансгрессией, а почти с полным совпадением, так как у тихоокеанской сельди количество позвонков колеблется от 47 до 57, а у атлантической сельди, если принять во внимание все ее формы, а не одну лишь мурманскую, — от 51 до 60. У атлантической сельди даже средние числа позвонков у разных рас колеблются от 54.7 до 57.5, между тем как на кривой пределы колебаний указаны от 55 до 59, что свойственно лишь мурманской сельди. Другим признаком, по которому многопозвонковые и малопозвонковые сельди резко различаются, Л. А. Пономарева считает развитие зубов на сошнике. В подтверждение Л. А. Пономарева ссылается на П. Ю. Шмидта и Л. С. Берга, которые однако считают обеих этих сельдей лишь за подвиды. Со ссылкой на стр. 182 на рисунок «сошников сельдей группы harengus и группы pallasi, наглядно подтверждающий справедливость указаний, упомянутых автором», получилось явное недоразумение, так как этого рисунка в статье нет. На этом рисунке изображена карта распространения атлантической сельди. Что касается различий в числе поперечных рядов чешуй (перед спинным плавником), по которому «всегда легко отличить группу harengus от группы pallasi», то число их, как видно из табл. 3, также трансгрессивно: у форм первой колеблется от 13 до 15, у второй — от 9 до 13 (на стр. 189 от 10 до 13). Относительно различий в числе сегментов у личинок сказано выше. Следует лишь отметить, что в этой части статьи имеется также ряд ошибочных утверждений. Так, личинки беломорской сельди, по данным Каринского (1938), имеют не 53 сегмента, а 53 и более, анальное отверстие у них находится не на 43—44 сегменте, а на 42—45 и т. д.
Распространение. Северные части Атлантического и Тихого океанов и прилегающие моря. В Атлантическом океане у берегов Европы от Бискайского зал. до Шпицбергена, Новой Земли, в Балтийском, Белом и Карском морях, у берегов Сев. Америки от м. Гаттераса до Девисова прол. и Гренландии. В Тихом океане у берегов Азии от западного побережья Кореи в Желтом море и зал. Тояма в Японском, на север до Анадырского зал., в бухте Тикси и Оленёкском зал. у берегов Сибири, у берегов Сев. Америки от Калифорнии до Аляски (рис. 14).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА
1 (4). Число позвонков 51—60, наичаще 55—57. Килевые чешуи более или менее развиты как позади, так и впереди основания брюшных плавников. Между брюшными и анальным плавниками 12—16 килевых чешуй. Зубы на сошнике сравнительно хорошо развиты. Северная часть Атлантического океана и прилегающие моря.
2 (3). Позвонков в среднем от 54.8 до 57.5. Грудные плавники короткие, менее 14% длины тела. Голова короткая и широкая, длина ее менее 23% длины тела, межглазничный промежуток более 16.5% длины головы. Глаза небольшие, менее 24% длины головы. Северная часть Атлантического океана и прилегающие моря — Северное, Баренцово и отчасти Белое. *la. Clupea harengus harengus L.
3 (2). Позвонков меньше, в среднем от 55.2 до 55.7. Грудные плавники
длиннее, не менее 13.5% длины тела. Голова длиннее и уже, длина ее более 23% длины тела, межглазничный промежуток менее 17.5% длины головы. Балтийское море. *lb. Clupea harengus harengus n. membras L.
4 (1). Число позвонков 49—57, наичаще 52—55. Килевые чешуи более или менее развиты лишь позади брюшных плавников. Между брюшными и анальным плавниками 10—14 килевых чешуй. Зубы на сошнике менее развиты. Северная часть Тихого океана и арктические моря.
5 (6). Голова короткая, менее 21% длины тела. Грудные плавники менее 14.5% длины тела. Глаза менее 23% длины головы. Крупные рыбы, в половозрелом состоянии длиной от 20 до 40 см и более. Северная часть Тихого океана и прилегающие моря. *lc. Clupea harengus pallasi Val.
6 (5). Голова длиннее, более 21% длины тела. Грудные плавники более 13% длины тела. Глаза более 23% длины головы. Размеры меньше, до 30—34 см, в половозрелом состоянии — от 10—20 см. Белое море, Мезенский зал., Чешская губа, Карское море на восток до Обской губы.
7 (8). Высота головы более 13% длины тела. Берг (1948 : 148) в определительной таблице в качестве отличительного признака между Clupea harengus pallasi n. maris-albi и Clupea harengus pallasi n. suworowi указывает длину основания анального плавника, меньшую у первого (около 10.5% длины тела), чем у второго (более 11%). Данные эти взяты из работ разных авторов. По нашим данным, длина основания анального плавника больше у Clupea harengus pallasi n. maris-albi (10.9%), чем у Clupea harengus pallasi n. suworowi (10.4%). Белое море. *ld. Clupea harengus pallasi n. maris-albi Berg
8 (7). Высота головы менее 14% длины тела. Мезенский зал., Чёшская губа, Карское море. *1е. Clupea harengus pallasi n. suworowi Rabinerson.
Рыбы пресных вод СССР и сопредельных стран. Л.С.Берг 1948 г.
Род CLUPEA LINNS. — МОРСКАЯ (ОКЕАНИЧЕСКАЯ) СЕЛЬДЬ
Clupea Linne, Syst. nat. ed. X, 1758, p 317 (тип: Clupea harengus). — Берг, Мат. к позн. русск. рыбол., II, вып. 3, 1913, стр. 4.
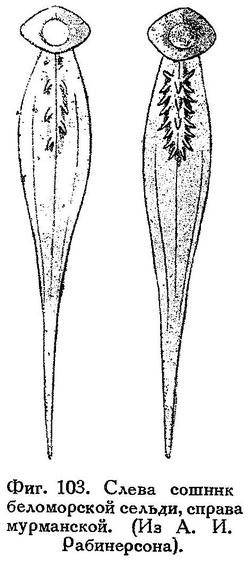
Зубы на сошнике (слабые) есть (фиг. 103), иногда на языке, на praemaxillare, на нёбных костях (едва развитые), на нижней челюсти спереди. На мезоптеригоиде зубов нет.
Веки зачаточные. Сочленение нижней челюсти с черепом под серединой глаза. Нижняя челюсть заметно выдается вперед. Верхняя челюсть хватает кзади до вертикали середины глаза. Вырезки на praemaxillare нет. Длина щек более их высоты. Крышечная кость (operculum) гладкая, без полосок. Брюшные плавники под передней третью спинного. Начало спинного плавника немного ближе к концу рыла, чем к основанию хвостового плавника. Последние два луча анального плавника не увеличены. Брюшные шипы (килевые чешуи) слабые, едва заметные, числом 36—48; брюхо закруглено. D III—V 13—17, всего 16—22 (У берегов Америки бывает и 15 (Rounsefell, 1930, р. 263).), А II—IV 12—17 (всего 13—20), Р I 15—17, V I 7—8 (9) (Обычно I 8; как очень редкое исключение бывает I 6 и даже I 5; см. Неinckе, 1898, р. 99. tab. VIII, fig. 2, 4.). Жаберных тычинок около 65—70. Поперечных рядов чешуй 53—70. Больших чешуй (alae) на основании хвостового плавника нет. Позвонков (45) 48—60. Пилорических придатков 18—29. Слепые отростки плавательного пузыря помещаются как в prooticum, так и в pteroticum (W. Ridewood, Proc. Zool. Soc. London, 1904, II, p. 490.). Икра прилипающая (не пелагическая), без жировой капли, с малым околожелтковым пространством.
Северные части Атлантического и Тихого океанов. Один вид Clupea harengus, образующий множество местных форм. У берегов Новой Зеландии водятся мелкие селедочки, очень близкие к роду Clupea, но отличающиеся отодвинутым кзади спннным плавником и брюшными плавниками, начинающимися под началом спинного или несколько впереди начала спинного. Позвонков 42—46. Перечисленными признаками эти селедки, относимые Ригэном к роду Clupea (Clupea antipodum Hector и Clupea muelleri Klunz.; см.: С. Т. Regan, Ann. Mag. Nat. Hist. (8), XIX, 1917, p. 227—228), приближаются к килькам, или шпротам, p. Sprattus Girg. (тип: Clupea sprattus L.), но у последнего обычно нет зубов на сошнике. По всей видимости, названные новозеландские селедочки составляют особый род. Берега Азии и Европы от Обской губы и Белого моря до Бискайского залива (Аркашон), зап. Шпицберген, Медвежий остров, берега Исландии, южн. Гренландии, Ньюфаундленда, по американскому побережью на юг до м. Гаттерас (35° с. ш.). В Тихом океане распространен от Берингова пролива на юг по азиатскому побережью до Хондо, Кореи (Фузан, Цзын-нам-пхо), Шань-дуна (Желтое море), а по американскому — до Калифорнии (Сан-Диего, 32.5° с. ш.). Это — морская сельдь, обычно не входящая в реки. Но она не избегает и очень опресненной воды и иногда, как исключение, попадается и в совершенно пресной воде. В рассматриваемых пределах образует формы:
1 (4). Число позвонков наичаще 57—55. Килевые чешуи более или менее развиты как позади, так и впереди основания брюшных плавников. На сошнике сравнительно крупные зубы. — Атлантический океан, Сев. Ледовитое море.
2 (3). Позвонков наичаще 57. — Мурман, Белое море. 1. Clupea harengus harengus L.
3 (2). Позвонков наичаще 55—56. — Балтийское море. 1а. Clupea harengus membras L.
4 (1). Число позвонков наичаще 53—54. Килевые чешуи более или менее развиты лишь позади основания брюшных плавников. На сошнике слабые зубы. — Тихий океан, Сев. Ледовитое море (и Белое).
5 (8). Половозрелые длиной до 30 см, изредка до 34 см.
6 (7). Длина анального плавника в среднем около 10.5 длины всего тела (При описании Clupea harengus все пропорции относятся к абсолютной длине тела (до конца крайних лучей С).). — Белое море. lb. Clupea harengus pallasi n. maris-albi Berg
7 (6). Длина анального плавника в среднем более 11% длины всего тела. — Чошский залив, Печорский залив, Карская губа. lb. Clupea harengus pallasi n. suworowi Rabinerson
8 (5). Половозрелые длиной от 25 до 40 см и даже до 50 см. — Дальний Восток. lb. Clupea harengus n. pallasi Valenciennes