(Anarhichadidae Bonaparte, 1835) Семейство Анарихадиды, Зубатковые, Зубатки, Зубаток, Family Anarhichadidae Bonaparte, 1835 (Wolffishes) 2 рода 5 видов
Материал подготовил Фактистов И. Е., 2020 г. Сайт www.fishbiosystem.ru
Семейство Anarhichadidae впервые описал в 1835 году французский натуралист Шарль Бонапарт (Шарль Люсьен Бонапарт — Charles Lucien Bonaparte; Шарль Люсьен Жюль Лоран Бонапарт или Карло Лучано Джулио Лоренцо Бонапарте — фр. Charles Lucien Jules Laurent Bonaparte или итал. Carlo Luciano Giulio Lorenzo Bonaparte, 1803 — 1857, французский биолог и орнитолог, племянник императора Наполеона, посещал различные итальянские университеты, затем отправился в США, где посвятил себя изучению природы, издал фундаментальное сочинение «Американская орнитология» (3 тома, Филадельфия, 1825), составившее ему репутацию в научном мире, по возвращении из Америки поселился в Риме, где издал труд «Иконография итальянской фауны» (Позвоночные, 3 т., Рим, 1833—41, большой фолиант) и ещё несколько научных трудов, в 1845 году избран членом Королевской академии наук, литературы и изящных искусств Бельгии (Royal Academy of Science, Letters and Fine Arts of Belgium), в 1855 году — иностранным членом Шведской королевской академии наук (Royal Swedish Academy of Sciences), автор описаний 21 нового вида рыб, в его честь назван 1 вид рыб).
Анарихадиды, Зубатковые, Зубатки (лат. Anarhichadidae) — семейство морских лучепёрых рыб из отряда окунеобразных (Perciformes Bleeker, 1859), обитающих в умеренных и холодных зонах морей северного полушария.
Название семейства Anarhichadidae происходит от греческого слова αναρριχηση (anarrhichesis, anarrichisi) — «скалолазание», «лазание», вероятно, имеется в виду тяга зубаток к каменистому дну и скалистым берегам. По другим данным, название семейства происходит от древнегреческого слова αυαρριχαομαι (avarrichaomai) — «взлезать», «вскарабкиваться». Происхождение этого названия таково. Рыбаки Каттегата и Скагеррака прежде называли полосатую зубатку «Klippfisch» за то, что она обитает на дне между камней. Геснер (Gesner, 1598 : 63), кроме правильного толкования народного названия, предположил, что зубатка вылезает из воды на скалы, а поэтому дал ей греческое название Anarrhichas, принятое позднее Артеди (Artedi, 1793 : 38; Линней писал это слово через одно r). Возможно, на самом деле в те времена ходила легенда, будто бы зубатка вылезает на скалы греться на солнце, как сообщают об этом последующие авторы; легенд про нее и позднее писалось много: будто грызет якорные цепи, оставляя на них следы зубов (Schonevelde, 1624 : 45). [Фауна СССР. Рыбы. Том V, вып. 5. Семейство Зубаток (Anarhichadidae). В.В.Барсуков 1959 г.]
Английское название «wolffishes», в дословном переводе — «рыбы-волки», указывает на мощные клыки и крепкие молярные зубы у этих рыб.
Время появления современных родов семейства Зубатковые: Anarhichas — в начале плиоцена; Anarrhichthys — в начале плейстоцена. [Sepkoski, Jack. A compendium of fossil marine animal genera // Bulletins of American Paleontology — Paleontological Research Institution, 2002. — Vol. 364. — P. 560.]
У зубаток тело умеренно удлинённое, в большей своей части сжатое, за исключением одного вида Anarrhichthys ocellatus с чрезвычайно удлинённым телом, за что получил название «wolf-eel» — «волчий угорь» (см. Фото). У анарихадид спинной плавник длинный, начинается от головы и состоит из 69—88 гибких шипиков у Anarhichas и 218—250 у Anarrhichthys. Анальный плавник с 42—55 мягкими лучами у Anarhichas и 0—1 шипиком и 180—233 мягкими лучами у Anarrhichthys. Хвостовой плавник отделен от спинного и анального плавников сильно укороченными лучами у Anarhichas, а у Anarrhichthys срединные (непарные — спинной, анальный и хвостовой) плавники сливаются, сужаясь к точке. Грудные плавники крупные и округлые, с 18—24 лучами. Брюшные плавники отсутствуют. Одна пара ноздрей. Чешуйки циклоидные, мелкие и неперекрывающиеся или отсутствуют. Механосенсорные каналы головы хорошо развиты, поры с возрастом зарастают: носовые 2, затылочные 3—5, межорбитальные 1—2, посторбитальные 4—5, суборбитальные 7—9, преоперкулярные 4, нижнечелюстные 3—4 (см. Фото). Одна или две боковые линии туловища поверхностных невромастов (superficial neuromasts), которые трудно различимы в консервированном материале. Зубатки обладают мощными широкими челюстями. У большинства разновидностей есть сильные клыки, похожие на собачьи, и дробящие зубы, расположенные в задней части челюстей и на нёбе (см. Фото), чтобы выкапывать и разгрызать моллюсков и другую добычу с твердой оболочкой. Зубы меняются каждый год. Жаберные перепонки прикреплены к перешейку, жаберные отверстия широко разделены. Лучей жаберной перепонки — бранхиостегальных лучей (branchiostegal rays) 6—8. Плавательный пузырь отсутствует. В позвоночнике от 72—89 до 251 позвонков. Размеры зубаток относительно крупные, взрослые особи достигают длины 110—240 см и массы 20—32 кг. Максимальная длина около 2,5 м.
Распространение (карта): обитают в умеренных и холодных зонах морей северного полушария, Северная Атлантика и Северная часть Тихого океана. Зубатки преимущественно демерсальные (донние) рыбы, населяющие мелкие или умеренно глубокие холодные воды. Держатся недалеко от берега на глубинах до 300—1700 м.
Питаются зубатки моллюсками, иглокожими, высшими ракообразными, медузами и рыбой. Обладая мощными широкими челюстями с бугорковидными зубами (см. Фото), зубатки способны раздавливать толстые стенки раковин.
Некоторые виды являются объектами промысла и спортивной рыбалки.
Рыбы открытого океана. Н.В.Парин 1998 г.
Зубатки (два рода семейства Anarhichadidae), распространенные в северных частях Атлантического и Тихого океанов, живут в основном в шельфовой зоне, но в зимнее время мигрируют на глубины (эта группа всесторонне изучена В. В. Барсуковым). Наиболее глубоководный вид - синяя зубатка (Anarhichas denticulatus), достигающая 1,4 м в длину, отмечена даже на глубине более 900 м. Все зубатки откладывают очень крупную донную икру, но их личинки поднимаются в толщу воды, где проходят весь мальковый период жизни (его продолжительность у разных видов сильно разнится).
Атлас-определитель рыб: Книга для учащихся. Н.А.Мягков 1994 г.
Семейство Зубатковые (Anarhichadidae)
Тело удлиненное, в поперечном сечении округлое. Чешуя мелкая, глубоко погружена в плотную кожу. Голова голая, рыло тупое с очень большим ртом. Зубы крупные, явно дифференцированные на три типа: клыковидные, конические и бугорковые (приспособлены для перетирания пищи — организмов бентоса). Боковая линия двухветвистая: верхняя ветвь идет по туловищу, нижняя — по хвостовому стеблю. Морские и реже солоноватоводные рыбы. Ценные объекты промысла. В наших водах несколько видов, но в прибрежной зоне обычны два.
Рыбы Японского моря и сопредельных частей Охотского и Желтого морей. Часть 4. (Perciformes). Г.У.Линдберг, З.В.Красюкова 1975 г.
НАДСЕМ. STICHAEOIDEA
В. М. Макушок (1958 : 63) сближает семейство Stichaeidae с семействами Ptilichthyidae, Pholididae и Anarhichadidae и, объединяя их в Stichaeoidea, дает этому надсемейству следующую характеристику.
Тело умеренной длины или сильно удлиненное, покрыто мелкой циклоидной чешуей, которая может заходить на голову и плавники или в различной степени редуцироваться (если чешуя не заходит на голову, то сверху она ограничивается затылочным каналом). С каждой стороны по одному небольшому непарному ноздревому отверстию, которым оканчивается удлиненная обонятельная трубочка. Жаберных дуг 4, за последней дугой щель не зарастает. Дыхательные перепонки (нёбная и нижнечелюстная) обычно имеются (полностью редуцируются лишь у родов Anisarchus и Lumpenella). Плавательный пузырь отсутствует. Сейсмосенсорные каналы головы открываются наружу посредством пор. У преобладающего большинства форм туловищные сейсмосенсорные каналы отсутствуют (имеются лишь у некоторых групп Stichaeidae). Количество позвонков варьирует от 43 (Stichaeopsis hopkinsi, известен у восточного берега Камчатки и у о. Ванкувер.) до 240 (Ptilichthyidae, распространен в северной части Тихого океана.) и более (Anarhichthys). Спинной и анальный плавники длинные, сплошные, их перепонки доходят до основания хвостового плавника и в ряде случаев в большей или меньшей степени сливаются с последним. Спинной плавник обычно содержит одни колючие лучи, лишь у некоторых Stichaeidae и у Ptilichthys в его задней части имеются мягкие лучи. Анальный плавник начинается непосредственно за анальным отверстием, его передний луч, как правило, нечленистый, в виде колючки или шипа. Антеанальное расстояние обычно менее половины абсолютной длины тела. Хвостовой плавник никогда не бывает вильчатым (обычно овально-округлый), основных лучей 12—15 (обычно 13—14). Грудные плавники, если имеются, содержат от 8 до 21 луча. Брюшные плавники торакальные по положению, содержат 1 колючий и максимум 4 мягких луча; в большинстве групп полностью редуцируются. Мягкие лучи плавников обычно ветвятся 2—3 раза (Макушок, 1958 : 63).
Донные, преимущественно прибрежные рыбы; отдельные представители различных групп опускаются на значительную глубину (Anarhichas latifrons и Lumpenella). Многие виды проявляют заботу о потомстве, охраняя кладку икры. Икра донная, обычно крупная. Мальки прибрежно-пелагические. Преимущественно мелкие рыбы (исключая зубаток). Питаются обычно мелкими донными беспозвоночными. Как исключение имеются хищники (например, Stichaeus grigorjewi). Типично растительноядных форм, по-видимому, нет (Макушок, 1958 : 65).
Известен из северных частей Тихого (преобладающее большинство форм) и Атлантического океанов, а также из Северного Ледовитого океана. Южная граница ареала проходит через южную часть Японского моря, центральную Калифорнию, мыс Код и Ла-Манш, совпадая с северной границей ареала тропических бленниид.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА СЕМЕЙСТВ НАДСЕМ. STICHAEOIDEA
(По В. М. Макушку (1958 : 66), с изменениями.)
1 (6). Зубы не дифференцированы по форме. Сошниковая головка не удлинена; сошниковые зубы, если имеются, конические.
2 (5). В хвостовом плавнике ветвятся только основные лучи (рис. 26, А).
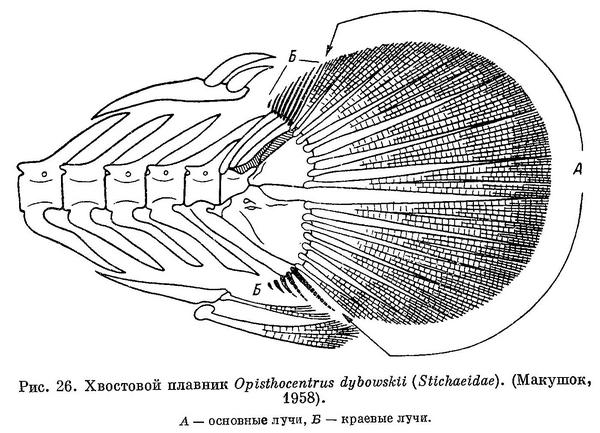
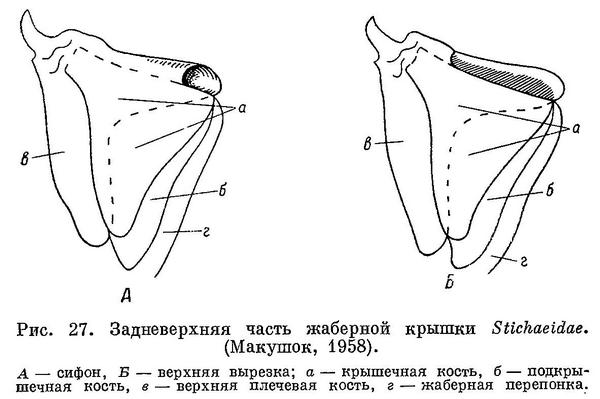
3 (4). Тело не очень сильно удлинено (не более 134 позвонков). Жаберные отверстия снизу заходят значительно выше верхнего края основания грудных плавников; сифон или верхняя вырезка жаберной крышки имеется (рис. 27, А, В). Лучей жаберной перепонки 5 или 6. Колючие лучи спинного плавника соединены друг с другом перепонкой. Кожистый подбородочный выступ отсутствует. 1. Stichaeidae.
4 (3). Тело чрезвычайно сильно вытянутое (238—240 позвонков). Жаберные отверстия снизу едва доходят до нижнего края основания грудных плавников; сифон или верхняя вырезка жаберной крышки отсутствуют. Лучей жаберной перепонки 3. Мак-Аллистер (McAllister, 1968 : 145) указывает 6. Колючие лучи спинного плавника не соединены перепонкой. Кожистый подбородочный выступ имеется. Один род и 1 вид (Ptilichthys goodei Bean, 1882). Мало находок. Восточное побережье Камчатки, р-н о. Уналашка и о. Ванкувер, зал. Пьюджет-Саунд (Макунгок, 1958 : 117). [Ptilichthyidae].
5 (2). В хвостовом плавнике ветвятся не только основные лучи, но и ближайшие к ним краевые лучи (рис. 26, Б). 2. Pholididae.
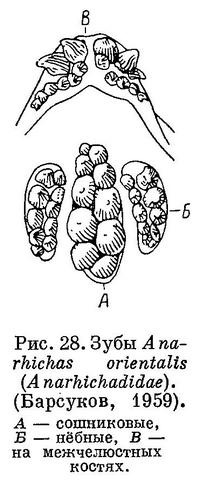
6 (1). Зубы резко дифференцированы по форме. Головка сошника сильно удлинена, несет массивно дробящие зубы (рис. 28, А—В). 3. Anarhichadidae.
CXLV. Сем. ANARHICHADIDAE — ЗУБАТКОВЫЕ
(Описание семейства частично заимствовано из монографии В. В. Барсукова (1959) и дополнено сведениями из монографии В. М. Макушка (1958).)
Тело умеренно или сильно удлиненное, сжатое с боков, покрыто очень мелкой, тонкой, погруженной в кожу чешуей, не налегающей друг на друга. Голова не покрыта чешуей. Усиков нет. Рыло тупое, рот большой. Верхняя челюсть слабо выдвижная. Зубы крупные, дифференцированы на три основных типа (клыкообразные, конические и бугорковидные); наиболее мощные зубы расположены на сошнике и противостоящих ему частях нижнечелюстных костей (см. рис. 28). Верхнеглоточные зубы расположены на трех площадках. Зубы челюстей и нёба приспособлены к разрыванию и раздавливанию организмов с твердым панцирем (моллюски, ракообразные, иглокожие). Жаберные отверстия широко разделены межжаберным промежутком. Жаберная щель широкая. Имеется сифон жаберной крышки. Лучей жаберной перепонки 6—8 (чаще 7). Спинной плавник один, простирается от затылка до хвостового плавника или сливается с ним (у Anarhichthys, который распространен по тихоокеанскому побережью Северной Америки от о. Кадьяк (Аляска) до г. Сан-Диего (Барсуков, 1959).). Лучи спинного плавника все колючие, они либо гибкие на протяжении всего плавника, либо утолщающиеся и отвердевающие по направлению к заднему концу плавника. Грудные плавники большие, округлые, расположены низко. Брюшных плавников нет. Лучи грудного и хвостового плавников дважды-трижды ветвятся, как и несколько самых задних лучей анального плавника. Остальные лучи анального плавника не ветвятся. В начале анального плавника 1 недоразвитая колючка. Число пор каналов боковой линии головы варьирует. Каналы боковой линии на теле отсутствуют: сейсмосенсорные почки сидят открыто в виде верхней и средней ветвей.
Два рода. Один из них (Anarhichas) известен в Японском море.
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
Семейство Зубатковые (Anarhichadidae)
Этих рыб недаром назвал народ
зубатками — их мощные зубы, и прежде всего
торчащие изо рта клыки, бросаются в
глаза. А проглядишь — зубатка может
напомнить: цапнув за ногу зазевавшегося
рыбака, она способна прокусить сапог,
а то и достать до тела. Все это породило
немало легенд о весьма хищном нраве
зубаток, вплоть до ходившего в средние века
поверья, будто зубатки заранее
собираются на том месте, где должно произойти
кораблекрушение, чтобы полакомиться
редкой добычей.
Конечно, зубатки пускают свои клыки
в ход, не только обороняясь, но и нападая
на других рыб, иногда и на других
зубаток. Однако это случается не так уж
часто, клыки обычно употребляются для
другой цели, а именно для отрывания от
дна цепляющихся за него моллюсков,
иглокожих и ракообразных, раковины
и панцири которых зубатки дробят или
разрывают бугорковидными и
коническими зубами, сидящими на нёбе и на нижней
челюсти позади клыков. От такой работы
зубы быстро снашиваются, но ведь срок
их службы невелик: каждый год, в те
месяцы, когда воды в местах обитания
зубаток охлаждаются, все старые зубы
вываливаются, а из-под них поднимаются новые
зубы на новых, пока еще мягких цоколях.
В это время зубатки хватают лишь
добычу, не защищенную панцирем, или совсем
не питаются. Месяца через полтора
цоколи окостеневают, а в их основаниях снова
появляются зародыши зубов, которые
растут внутри слившихся друг с другом
цоколей, постепенно разрушая их вплоть до
новой смены.
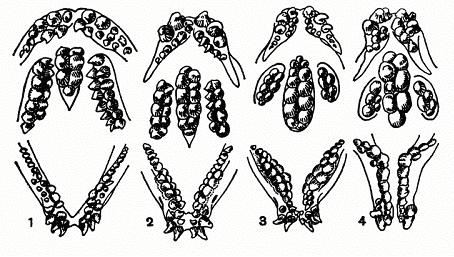
Рис. 193. Зубы взрослых зубаток:
1 — синей (Anarhichas latifrons); 2 — пятнистой (Anarhichas minor); 3 — обыкновенной (Anarhichas lupus); 4 — угревидной (Anarrhichthys ocellatus).
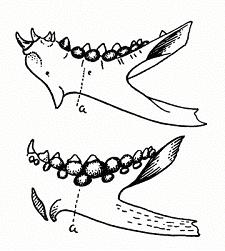
Рис. 194. Смена зубов
у зубатки.
Как и у других животных с крепкой
хваткой, челюсти зубаток спереди
укорочены. Сильно развитые «жевательные»
мышцы выпирают в виде желваков.
Круглая голова с выступающими изо рта
клыками немного напоминает кошачью,
отчего англичане и немцы иногда называют
зубаток «морскими кошками». Эта
необычная для рыбы форма головы издавна
привлекала внимание, и в старину
архангельские поморы, а также исландские
рыбаки любили украшать свои жилища,
привешивая к потолку сушеную голову
зубатки.

Тело зубатки довольно длинное; когда
зубатка плывет, по нему проходят
сильные изгибы, подобные изгибам
плывущего тритона. Это чисто поверхностное
сходство усиливают длинные непарные
плавники — спинной и анальный, но хвостовой
плавник от них обособлен, он имеет
округлую или усеченную форму, обычную для
рыб, плавающих не быстро. Грудные
плавники большие, веерообразные. Брюшных
плавников нет, но у дальних предков
зубаток они когда-то были, что видно по
тазовым костям современных зубаток,
прикрепленным к плечевому поясу, как и у
большинства других рыб из отряда
окунеобразных. Уменьшение или полная
утрата брюшных плавников, связанная с
удлинением тела и увеличением его гибкости,
наблюдается и у многих других рыб из
подотряда собачковидных, к которому,
помимо зубаток, принадлежат морские
собачки, бельдюга и другие мелкие
морские рыбы, среди которых зубатки
выглядят гигантами.
Семейство зубаток содержит 5 видов,
из которых три (полосатая, пятнистая и
синяя) обитают в северной части
Атлантического океана и два (дальневосточная и
угревидная) в северной части Тихого.
В мясе много витамина А, возможно, в
связи с быстрым ростом; вообще, чем больше
глубины, на которых предпочитают
держаться атлантические зубатки, тем
быстрее они растут: например, в возрасте 7 лет
беломорская зубатка достигает в среднем
37 см, баренцевоморская полосатая 54 см,
пятнистая 63 см и синяя 92 см.
Фауна СССР. Рыбы. Том V, вып. 5. Семейство Зубаток (Anarhichadidae). В.В.Барсуков 1959 г.
ПРЕДИСЛОВИЕ
Представители небольшого семейства зубаток, принадлежащего к подотряду Blennioidei из отряда окунеобразных, населяют северные части Атлантического и Тихого океанов. В Тихом океане обитают 2 вида зубаток — Anarhichthys ocellatus и Anarhichas orientalis, в Атлантическом 3 — Anarhichas lupus, Anarhichas minor и Anarhichas latifrons. Атлантические зубатки являются объектом промысла, в том числе и отечественного. Правда, зубатки имеют лишь второстепенное промысловое значение, несмотря на то, что их добывается в настоящее время примерно в семь раз больше, чем в начале текущего столетия. Однако с практической точки зрения исследование непромысловых и малопромысловых рыб не менее важно, чем изучение рыб промысловых. Вряд ли изменения численности последних могут быть достаточно хорошо поняты без представления о характере и степени влияния на них численности первых. К тому же промысловое значение каждого вида рыб изменяется в связи с характером развития промысла, в частности, с усовершенствованием орудий лова: исследование рыб, в настоящее время не используемых или слабо используемых промыслом, дает возможность в какой-то степени представить их будущее промысловое значение. Изучение зубаток, отличающихся ярко выраженной специализацией к питанию организмами с прочным наружным скелетом, представляет также немалый теоретический интерес.
МОРФОЛОГИЯ
При изучении морфологии зубаток основное внимание обращалось, как указано выше, на зависимость строения этих рыб от их образа жизни. Тем не менее в текст включены и такие морфологические данные, которым пока еще не найдено функционального объяснения: они могут оказаться интересными при дальнейшем изучении зубаток.
Изложение материала следует начать с органов, непосредственно связанных с наиболее заметной биологической особенностью зубаток — их специфическим питанием.
Зубы
Зубатки питаются по преимуществу организмами с твердым панцирем: моллюсками, иглокожими и высшими ракообразными. Приспособление к питанию такого рода прежде всего сказывается на строении зубов, своеобразие которых у зубаток настолько бросается в глаза, что находит отражение в названиях этих рыб на языках всех народов, знакомых с зубатками. Вполне понятно, что ни один из ихтиологов при описании видов зубаток, а описания эти довольно многочисленны, не проходит мимо описания их зубов. Тем не менее известные нам авторы отмечают связь между формой зубов и их функцией обычно лишь мимоходом, редко вдаваясь в детали.
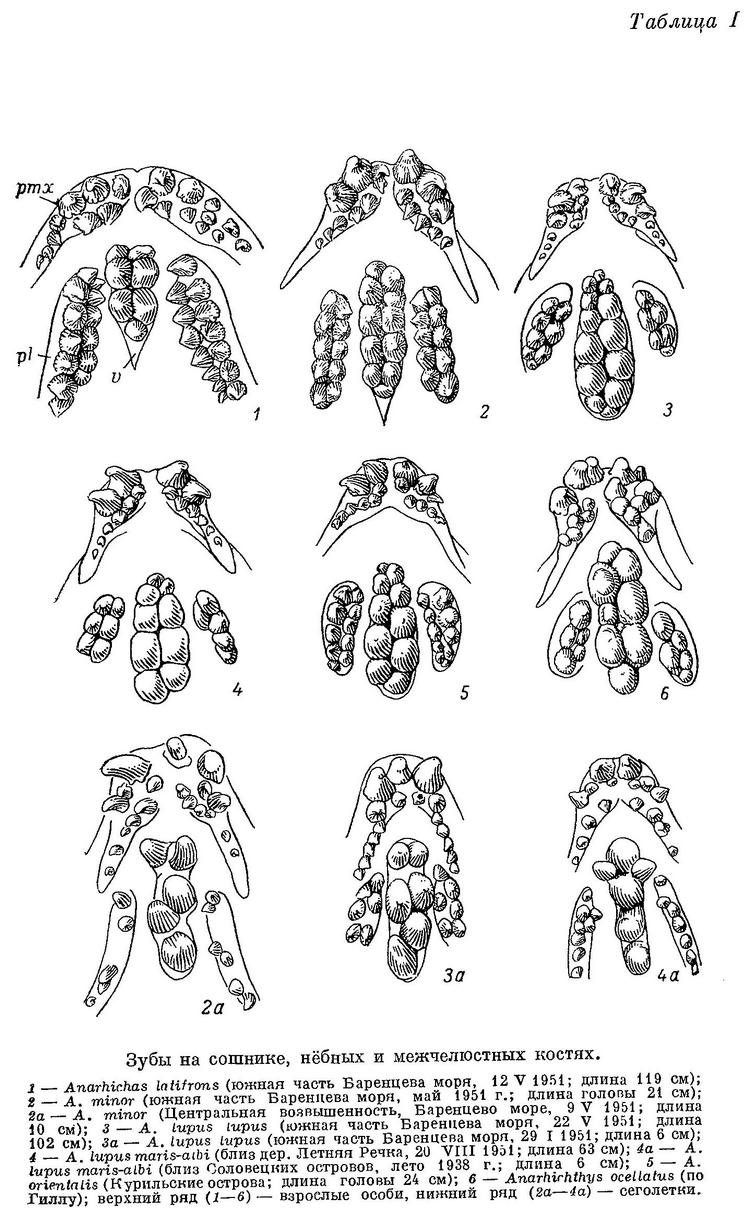
Все виды зубаток имеют очень много общих черт в строении зубной системы. У всех зубы дифференцированы, правда без резких границ, на 3 основных типа: крючковидные (схватывающие), конические (рвущие) и бугорковидные (давящие). Крючковидные зубы принято называть «клыками» в силу поверхностного сходства с клыками млекопитающих; по той же причине бугорковидные зубы называют иногда «коренными». Костями, несущими зубы, являются межчелюстные, нёбные, сошник и нижнечелюстные (не считая глоточных). У одной из 80 Anarhichas lupus maris-albi, черепа которых были отпрепарированы, обнаружен зуб на эктоптеригоиде. Более того, расположение зубов на этих костях в общем сходно (табл. I—II). На каждой из названных выше костей обычно имеется два продольных ряда крупных зубов (между ними изредка может вклиниваться еще один ряд). Зубы двух соседних рядов располагаются в шахматном порядке по отношению друг к другу, часто в той или иной степени нарушенном. Основания зубов обычно соприкасаются. Все зубы сидят на цоколях, почти всегда не только соприкасающихся, но и сливающихся в одну костную массу. Высота цоколей соответствует высоте сидящих на них зубов. Основание каждого зуба изрезано складками, входящими в соответствующие складки на цоколе, с помощью чего достигается прочное соединение зуба с цоколем (легче сломать зуб или выломать его вместе с цоколем, чем оторвать от последнего). Между основаниями двух соседних цоколей обычно имеется углубление для замещающего зуба. Зубы меняются ежегодно и почти одновременно, но у молоди, по-видимому, нерегулярно и поочередно.
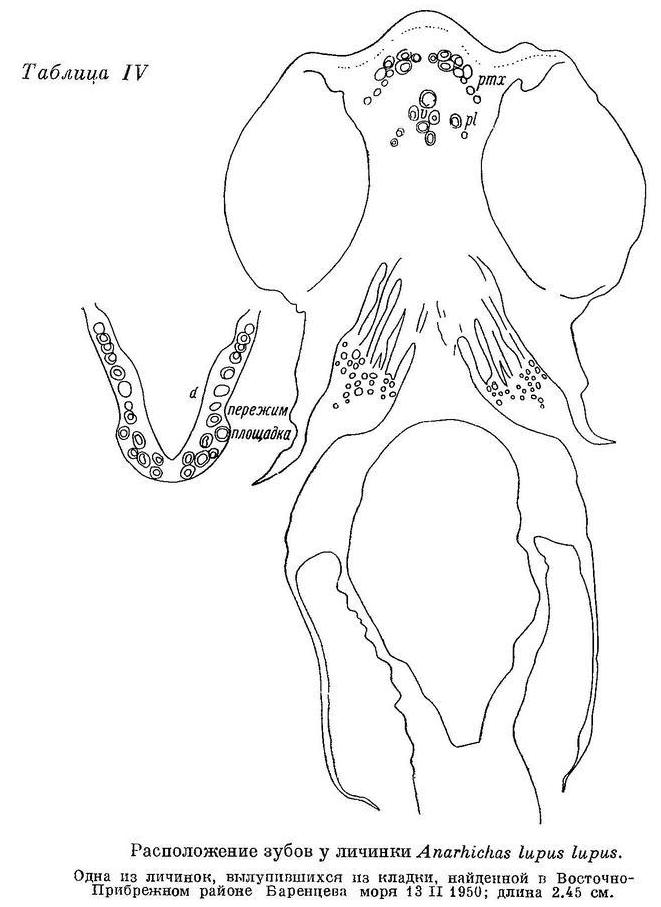
На межчелюстной кости внешний ряд зубов составляют большие клыки. Их размеры обычно уменьшаются спереди назад, а изогнутость назад возрастает. Внутренний ряд зубов состоит из мелких клыков, обычно загнутых внутрь и назад. Оба ряда располагаются под мощным вертикальным отростком межчелюстной кости. Сзади к ним примыкает ряд мелких клыков, расположенных на горизонтальной ветви кости, являющийся, очевидно, продолжением внутреннего ряда зубов. На нёбной кости зубы внешнего ряда почти всегда выше зубов внутреннего и обычно конические. Во внутреннем ряду все или часть зубов могут быть давящими. На сошнике два ряда давящих округлых зубов; передние часто бывают заострены. Макушок (1958 : 67) утверждает, что на сошнике зубаток поверхность, несущая зубы, представляет собою удлиненную головку этой кости, что вероятно, поскольку у личинок (Anarhichas lupus lupus; табл. IV) указанная поверхность коротка и расположена впереди таковых же поверхностей на нёбных костях. Тем не менее Макушок (1958 : 17) высказывает и противоположное мнение, возможно, ошибочное.
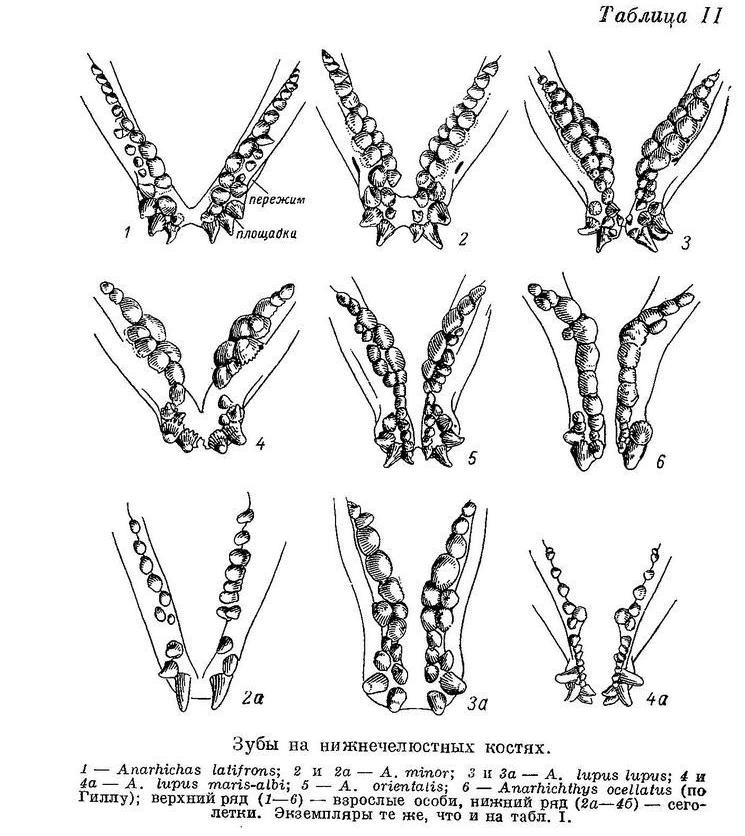
Передний конец нижнечелюстной кости, изогнутый наружу, занят клыками. Этот участок кости мы будем в дальнейшем для краткости называть «площадкой», а условную границу между площадкой и задним участком кости, прогнутым внутрь и вниз, — «пережимов» (табл. II, 1). Внешний ряд зубов начинается на площадке большими клыками. Как и на верхнечелюстной кости, передние клыки крупнее задних, но менее изогнуты назад. За клыками, уже на пережиме и далее, следуют конические зубы. Внутренний ряд зубов начинается на площадке мелкими клыками, изогнутыми назад и внутрь. За ними идут конические, затем бугорковидные зубы, сидящие на участке кости, прогнутом внутрь и вниз. Наибольшие из этих зубов находятся в самой низкой точке прогиба и более всего наклонены внутрь. Далее размеры зубов постепенно уменьшаются. Внутренний ряд зубов продолжается назад дальше внешнего. Внешние клыки нижнечелюстных костей находятся под внешними и задними клыками межчелюстных и при закрытом рте входят в промежутки между последними (у всех зубаток, исключая Anarhichas latifrons). Зубы сошника при этом приходятся посередине между внутренними рядами зубов нижнечелюстных костей, тогда как зубы каждой из нёбных костей располагаются над внешними, а частью и над внутренними зубами соответствующей нижнечелюстной кости.
Судя по всему, клыки зубаток служат главным образом для отрывания от субстрата прикрепляющихся и цепляющихся за него организмов, особенно моллюсков (Nilsson, 1855 : 210; Kent, 1883 : 57, и др.). Доказательством служит быстрая затупляемость клыков вскоре после их смены у всех зубаток, кроме Anarhichas latifrons. Такова основная функция клыков. Однако даже затупленные клыки служат зубаткам для обороны и нападения. Агрессивность зубаток часто отмечается рыбаками и водолазами с Белого и Баренцева морей, а также в литературе (Даллас, 1811 : 77; Gill, 1911 : 177; Виноградов, 1947 : 424, и др.). Случай нападения Anarhichthys ocellatus на Ophiodon elongatus почти таких же размеров описан Клеменсом и Вилби (Clemens a. Wilby, 1946 : 172). Одна из пятнистых зубаток (Anarhichas minor), посаженных мною в небольшой бассейн с водой, вскоре перекусила у другой верхнечелюстную кость. В ставной ловушке одна из беломорских зубаток (Anarhichas lupus maris-albi) вырвала у другой весь бок так, что печень вывалилась наружу. На Баренцевом и Белом морях мне нередко встречались зубатки с вырванными кусками плавников, чаще спинного. Подобные же изъяны в плавниках встречаются на рисунках зубаток и отмечаются в описаниях этих рыб (Krøyer, 1838 : 381; Книпович, 1902, фиг. 4; Bigelow a. Schroeder, 1935, fig. 3, и др.). По всей вероятности, это следы драк между самими же зубатками. В небольшом бассейне с морской водой, по стенкам которого была натянута сетка, зубатки часто лежали неподвижно, зацепившись клыками за эту сетку. Как было видно из вышеизложенного, наиболее крупные клыки расположены на верхнечелюстных костях по их внешнему краю, образуя полукруг; то же самое наблюдается на площадках нижнечелюстных костей. Подобное расположение клыков не позволяет моллюску или морскому ежу выскальзывать при отрывании. Конические зубы нижнечелюстных и нёбных костей рвут на части концы лучей крупных офиур и морских звезд, перед панцирем которых давящие зубы бессильны, так как он не ломается, а рвется. Наконец, бугорковидные зубы служат главным образом для раздавливания раковин моллюсков. При этом моллюск попадает на обе нижнечелюстные кости и раздавливается между их зубами, зубами сошника и отчасти нёбных костей. Выскальзыванию моллюска из-под давящих зубов препятствуют спереди клыки обеих челюстей, с боков — конические зубы нижнечелюстных и нёбных костей, сзади — задние зубы сошника (сошник сидит на парасфеноиде под углом к продольной оси тела так, что зубы по направлению назад опускаются все ниже, притом назад они простираются дальше, чем основные давящие нижнечелюстных). Кроме того, наклон давящих зубов на нижнечелюстных костях внутрь и опускание их вниз по сравнению с другими зубами само по себе ограничивает выскальзывание, так как общая поверхность зубов (если можно так выразиться) образует при этом нечто вроде выемки, в которую попадает моллюск при раздавливании. В связи с этим интересен рефлекс, который наблюдался у только что пойманных Anarhichas lupus maris-albi. Если притрагиваться к нижней челюсти зубаток сверху, над давящими зубами, то они обычно закусывают палочку лишь тогда, когда она касается обеих сторон челюсти. С помощью давящих зубов зубатки ломают также панцири морских ежей и крупных ракообразных. Давящие зубы имеют тенденцию соприкасаться друг с другом не только в основаниях, по и всей боковой поверхностью, функционируя как одно целое, что укрепляет всю их систему, а также предупреждает заклинивание между зубами обломков панцирей пищевых организмов (Lühmann, 1954а : 473).
Зубатки только дробят панцири пищевых организмов, но не перетирают их. Передние клыки даже при полузакрытом рте так заходят друг за друга, что боковое и продольное смещение нижней челюсти относительно верхней исключается. В кишечниках зубаток обломки раковин моллюсков, панцирей морских ежей, крабов и т. д. бывают обычно очень крупными; морские звезды и офиуры часто встречаются с целым диском, но с разорванными лучами.
Слизистая рта очень толстая, прочная и эластичная, что устраняет повреждаемость ее обломками.
До сих пор речь шла об общих чертах в строении зубной системы зубаток. Рассмотрим теперь различие между взрослыми особями отдельных видов.
У Anarhichas latifrons, в отличие от других зубаток, нижняя челюсть несколько укорочена. Когда рот закрыт, передние большие клыки нижней челюсти не вклиниваются между такими же клыками верхней, а располагаются позади них (табл. VII).
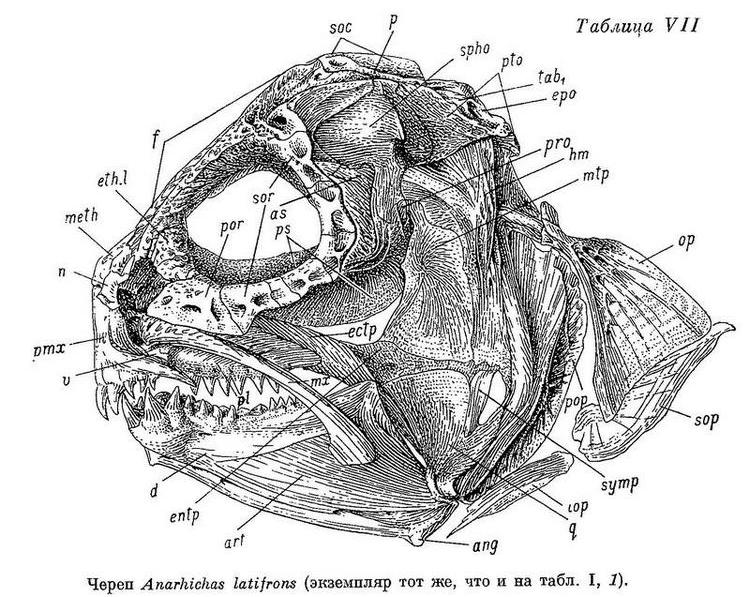
У Anarhichas minor передние клыки нижней челюсти входят в промежутки между передними клыками верхней. Клыки, как и все другие зубы, крупнее, чем у Anarhichas latifrons; сильнее стираются к концу лета; соединение их с цоколями прочнее ввиду большего развития складчатости оснований и уменьшения полости пульпы (табл. VI, 2).

У Anarhichas lupus lupus передние клыки еще длиннее, а разница между ними и малыми клыками еще больше; площадки сближены сильнее. Давящие крупнее, еще сильнее стираются к концу лета, как и другие зубы. Складчатость их оснований увеличивается, а полость пульпы уменьшается по сравнению с Anarhichas minor (табл. VI, 3).
У Anarhichas lupus maris-albi расположение зубов в общем сходно с таковым у Anarhichas lupus lupus. Однако заметно дальнейшее возрастание прогиба внутреннего ряда нижнечелюстных давящих вниз и внутрь и наклона этих зубов внутрь (табл. III, 4). Большие клыки и давящие заметно шире (табл. II, 4). Складчатость на основаниях давящих усиливается, полость пульпы уменьшается (табл. VI, 4). Зубы на нёбных отодвигаются еще дальше от клыков. Специализация к питанию моллюсками у этого подвида выражена еще сильнее. Передние клыки на нижней челюсти меньше наклонены вперед, чем у Anarhichas lupus lupus: вероятно, беломорской зубатке чаще приходится брать пищу не со дна, а с боковой поверхности камней.
У Anarhichas orientalis разница между большими и малыми клыками еще больше, чем у Anarhichas lupus, и все клыки шире; у них вновь уменьшается полость пульпы, а складчатость оснований, как и у других зубов, возрастает (табл. VI, 5).
У Anarhichthys ocellatus наибольшие нижнечелюстные давящие зубы оттесняются есще дальше назад (табл. II, 6), а ряды зубов на сошнике удлиняются (табл. I, 6). Складчатость оснований зубов несколько увеличивается (табл. VI, 6). Как и у Anarhichas lupus maris-albi, клыки на нижней челюсти меньше наклонены вперед и сама челюсть спереди массивнее, чем у других зубаток (табл. X, 2).
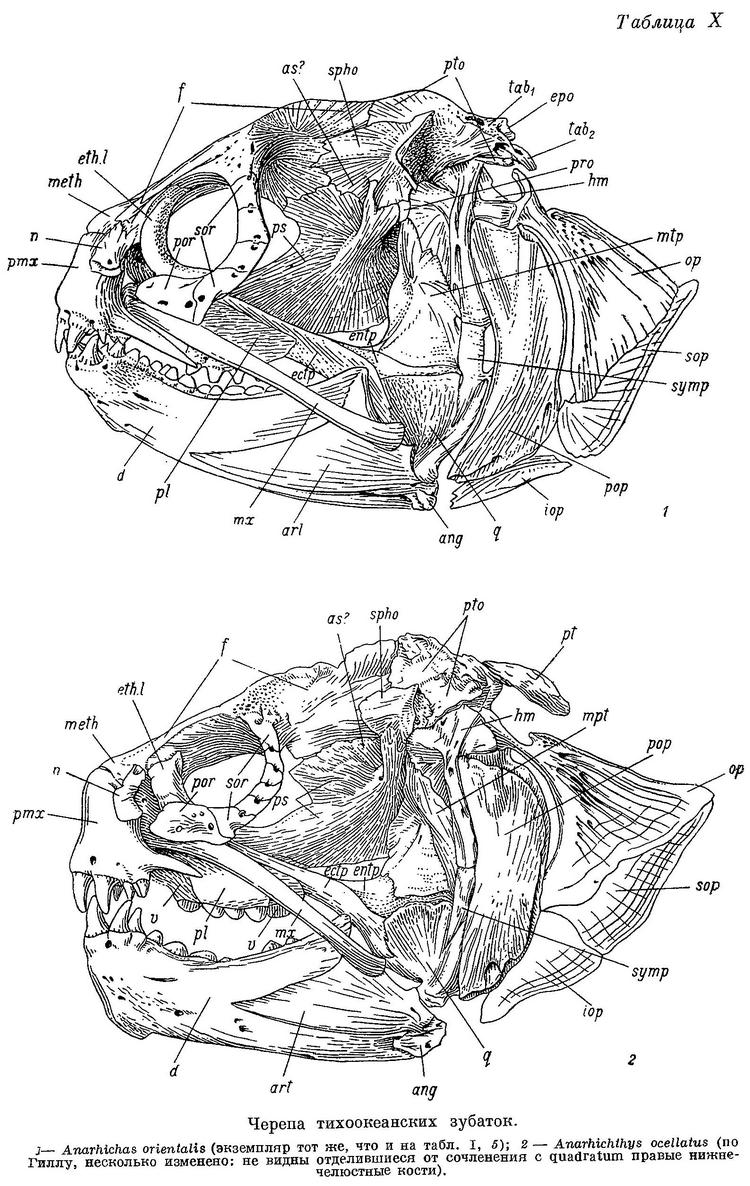
Как видно из вышеизложенного, в ряду взрослых особей Anarhichas latifrons — Anarhichas minor — Anarhichas lupus lupus — Anarhichas lupus maris-albi — Anarhichas orientalis — Anarhichthys ocellatus возрастает специализация зубной системы к питанию организмами с более прочным панцирем (в основном моллюсками).
Возрастание специализации происходит особенно быстро от Anarhichas latifrons к Anarhichas lupus, в дальнейшем замедляясь. По-видимому, различия в составе пищевых организмов (по форме и прочности их панцирей) между Anarhichas latifrons, Anarhichas minor и Anarhichas lupus больше, чем между Anarhichas lupus, Anarhichas orientalis и Anarhichthys ocellatus.
Не менее интересна возрастная изменчивость в строении зубной системы.
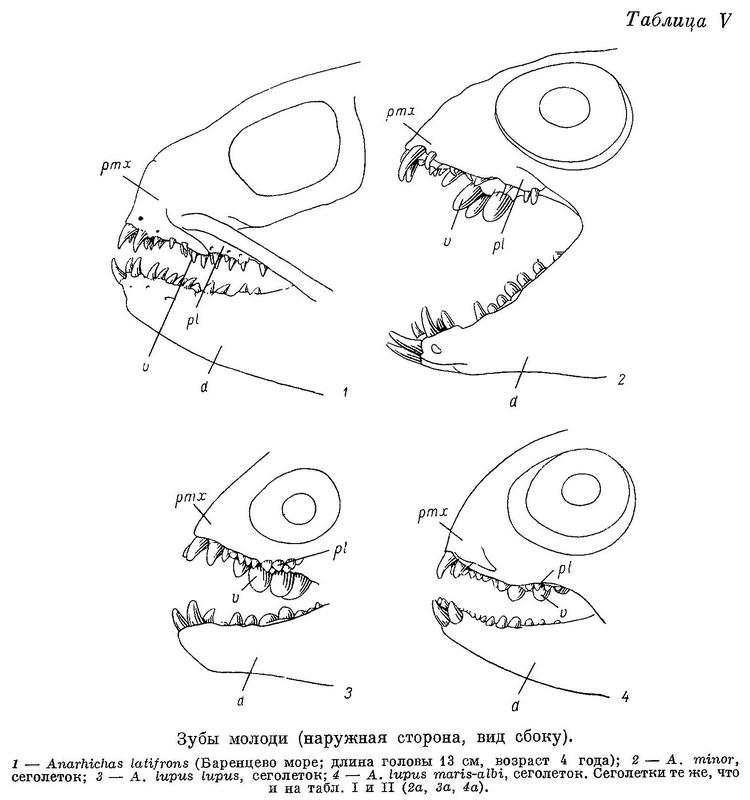
К сожалению, в моем распоряжении не было Anarhichas latifrons длиной менее 60 см. (Здесь и всюду далее, где упоминается длина рыбы, речь идет об абсолютной ее длине.) У молодых особей этого вида длиной 60—70 см малые клыки по величине лишь немного отличаются от больших. Судя по строению зубов, у молоди Anarhichas latifrons основную роль в питании играют подвижные организмы, лишенные панциря, что, вероятно, еще резче выражено у молоди меньших размеров. Переход к питанию крупными иглокожими и мелкими моллюсками, очевидно, происходит поздно, при длине не менее 70—80 см, т. е. в возрасте не менее 4 лет.
У сеголеток Anarhichas minor длиной 10—12 см большие клыки на межчелюстных костях по размерам сильно превосходят малые; малых клыков на нижнечелюстных часто вообще не бывает. Судя по строению зубов, у Anarhichas minor уже сеголетки питаются в основном иглокожими, а в следующие годы в рацион все больше включаются моллюски.
Личинки Anarhichas lupus lupus, выходящие из икры, имеют уже около 50 зубов (не считая глоточных). Все зубы острые. Судя по строению зубов, у Anarhichas lupus lupus уже сеголетки питаются в основном моллюсками.
По существу то же самое можно сказать о сеголетках Anarhichas lupus maris-albi длиной 6—7 см (с той разницей, что по расположению зубов они стоят ближе к личинкам) и сеголетках Anarhichas orientalis длиной 15 см.
У молоди Anarhichthys ocellatus все зубы, за исключением клыков, конические и становятся давящими лишь при достижении молодью длины 50—60 см (Kanazawa, 1952 : 572). Судя по строению зубов, молодь Anarhichthys ocellatus питается в основном иглокожими и к питанию моллюсками переходит довольно поздно.
Из вышеизложенного следует, что уменьшение роли подвижных организмов, лишенных панциря, и увеличение роли сначала иглокожих, а потом моллюсков в питании зубаток, наблюдающееся в ряду взрослых особей. Anarhichas latifrons — Anarhichas minor — Anarhichas lupus lupus — Anarhichas lupus maris-albi — Anarhichas orientalis — Anarhichthys ocellatus, намечается и в онтогенезе каждого из видов. При этом, как правило, чем сильнее выражена специализация у взрослых особей, тем раньше она наступает у молоди.
Функцию, требующую наибольшего приложения силы, сначала рвущую, а потом давящую, у всех зубаток выполняют зубы сошника и внутренний ряд нижнечелюстных зубов таким образом, что зубы сошника приходятся между нижнечелюстными костями, между внутренними рядами зубов на этих костях, а не напротив них. Эта самая существенная, черта в строении зубного аппарата зубаток, общая для всех видов и возрастов, оказывает весьма сильное влияние на расположение жевательных мышц, а через них и на строение черепа, как будет видно в дальнейшем.
В указанном выше ряду взрослых особей различных видов в помощь зубам сошника и внутренним нижнечелюстным все больше включаются зубы нёбных костей и внешние зубы нижнечелюстных. По существу то же самое наблюдается и в онтогенезе каждого из видов. Это подтверждает мнение Ван-Доббена (Van Dobben, 1937 : 40), считающего совместную работу нижнечелюстных костей с нёбными не примитивной, а вторично приобретенной чертой зубаток. Взаимодействие между нижнечелюстными костями и сошником у зубаток, судя по всему, было выработано еще раньше, чем взаимодействие между нижнечелюстными и нёбными костями.
Наблюдения над содержимым кишечников зубаток в основном подтверждают выводы о питании их, сделанные выше на основании изучения зубного аппарата. Для полноты этих данных требуется не только качественный, но и количественный анализ питания молоди и взрослых особей и также наблюдения над размером, формой и мощностью панцирей у организмов, входящих в его состав. Наблюдения эти должны охватить весь ареал каждого из видов и все сезоны. Такого рода наблюдения, надо думать, еще больше подтвердили бы вышеуказанные основные выводы о характере питания зубаток. Выводы эти были бы гораздо точнее и полнее, если бы имелась возможность работать не только с мертвыми экземплярами, но и с живыми в аквариуме. Целый ряд вопросов не может быть решен без непосредственного наблюдения над питающимися рыбами. Примером может служить, в частности, следующий: происходит ли раздавливание пищевого организма одновременно с его отрыванием от субстрата или это два различных акта? Можно предполагать, что в ряду Anarhichas latifrons — Anarhichas minor — Anarhichas lupus — Anarhichas orientalis — Anarhichthys ocellatus нарастет разрыв между актом отрывания моллюска от субстрата и актом его раздавливания, но без непосредственного наблюдения над процессом питания этих видов подобные предположения останутся не в меру смелыми.
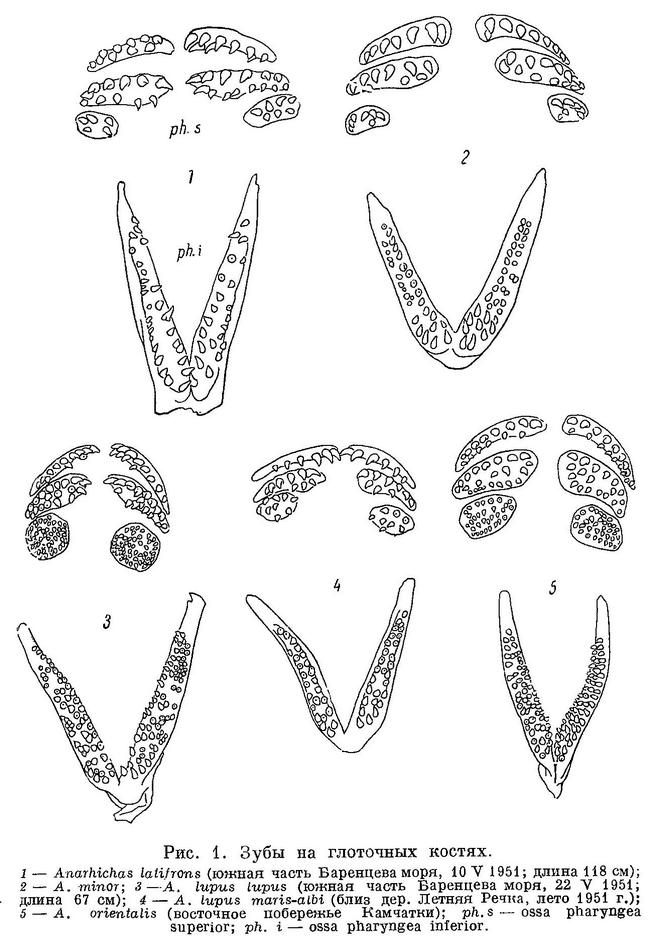
Глоточные зубы у всех зубаток устроены очень сходно (рис. 1). Мы не имели возможности наблюдать глоточные зубы у Anarhichthys ocellatus. Судя по краткому описанию Каназава (Kanazawa, 1952 : 572) и рентгеноснимку с молодого экземпляра, строение глоточных зубов у данного вида не отличается значительными особенностями, о деталях же по этим материалам сказать ничего нельзя. На каждой стороне глотки расположены 3 верхнеглоточных кости и одна нижнеглоточная, несущие зубы. Каждая верхнеглоточная вооружена, как правило, двумя рядами зубов. В переднем ряду зубы всегда многочисленнее и крупнее, чем в заднем. Внутри переднего ряда более крупные зубы располагаются обычно ближе к его внутреннему краю. На передней верхнеглоточной иногда бывает лишь один ряд зубов, реже это наблюдается на средней и никогда — на задней. Между двумя обычными рядами зубов иногда включаются беспорядочно сидящие зубы: на передней верхнеглоточной этого не бывает, на средней — бывает изредка, на задней — обычно. Задняя верхнеглоточная всегда меньше передних и более округла. На нижнеглоточных костях зубы расположены длинной полосой, состоящей из нескольких, реже только из двух, неправильных рядов. Внутренние и передние зубы обычно крупнее остальных. Передние концы обеих нижнеглоточных соединены крепкой связкой, задние расходятся. Соответственно ближе всего сходятся передние верхнеглоточные, а задние расходятся далее всего. Все глоточные зубы имеют одинаковую форму — похожи на малые клыки челюстей. Точно так же как и зубы на челюстях, они сидят на цоколях, между которыми с боков или сверху (когда зубы расположены между боковыми рядами) внедряются закладки замещающих зубов.
Видовые различия невелики. В ряду взрослых особей Anarhichas latifrons — Anarhichas minor — Anarhichas lupus lupus — Anarhichas orientalis высота глоточных зубов постепенно уменьшается, а количество их возрастает. У Anarhichas lupus lupus оно сильно возрастает на задней верхнеглоточной: участок, несущий зубы, становится почти круглым. У Anarhichas orientalis это выражено менее резко, но сильно увеличивается число зубов на средней верхнеглоточной. Anarhichas lupus maris-albi имеет наименьшее число глоточных зубов — внутривидовые отличия в этом признаке превосходят видовые. Молодь имеет меньшее число глоточных зубов, чем взрослые особи, — явление, по-видимому, общее для многих костистых рыб (Еремеева, 1950 : 30).

Движение глоточных зубов наблюдалось нами лишь у Anarhichas lupus maris-albi. Наблюдение производилось по методу А. П. Андрияшева (1944) — раздражалось глоточное кольцо и глоточные зубы у только что пойманных зубаток, до перерезки позвоночника за черепом и после нее. В обоих случаях было трудно добиться рефлекторного движения глоточных зубов. Ни разу не наблюдалось более одного цикла; последний состоял всего из 3 фаз (рис. 2).
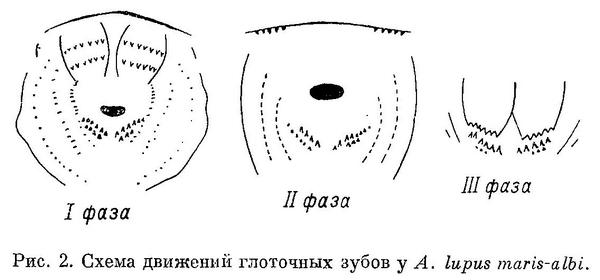
I фаза (состояние покоя) — верхнеглоточные кости обеих сторон сближены; их поверхности, несущие зубы, обращены вперед и вниз; задние концы нижнеглоточных костей расположены несколько выше передних соединенных концов.
II фаза (охват пищи) — верхнеглоточные кости сильно расходятся в стороны, втягиваясь в нёбо; их поверхности, несущие зубы, обращены вниз; движения нижнеглоточных костей не замечено.
III фаза (основное движение) — верхнеглоточные кости идут вниз и назад, к глотке; их зубы почти соприкасаются с зубами нижнеглоточных; движения последних опять-таки не замечено.
IV фаза (контрактурная пауза) отсутствует (свойствена хищным рыбам).
Нижнеглоточные кости изогнуты так, что при прохождении над ними верхнеглоточных все зубы последних почти соприкасаются с зубами первых. Не исключена возможность, что они не только проталкивают добычу в глотку, но выскребают мягкие ткани из обломков панцирей пищевых организмов, на что указывает и сильная снашиваемость глоточных зубов, отмеченная также Люманом (Lühmann, 1954а : 498).
Остается неясным, каким образом подается пища от челюстных зубов к далеко отодвинутым от них глоточным. Движения глоточных зубов, по-видимому, гораздо сложнее тех, которые нам удалось наблюдать, насколько об этом можно судить по сложной мускулатуре не только верхне-, но и нижнеглоточных костей.
Для раздавливания панцирей пищевых организмов у зубаток несомненно приспособлены только зубы челюстей и нёба, но не глоточные. Размер дробящего аппарата относительно общих размеров головы у зубаток больше, чем у других рыб с таким же аппаратом. Все это свидетельствует о тенденции к питанию возможно более крупными организмами с прочным панцирем, которых, может быть, другие рыбы использовать не могут. Гуд (Good, 1884 : 249) также считает, что зубатки питаются такими моллюсками и ракообразными, панцирь которых слишком прочен для других рыб. Своеобразное расположение противостоящих друг другу наибольших давящих зубов (сошник — нижнечелюстные) также может отчасти объясняться этой тенденцией. Она же могла явиться одной из причин, вследствие которых взрослые особи всех видов зубаток достигли самых крупных размеров среди всего подотряда Blennioidei.
Особо следует остановиться на весьма своеобразном процессе смены зубов у зубаток. Предварительное сообщение об этом было опубликовано ранее (Барсуков, 1954 : 897). Лишь позднее мне стало известно, что очень сходное сообщение было сделано еще раньше Люманом (Lühmann, 1951 : 241), а в 1954 г. тот же автор опубликовал весьма обстоятельную статью, посвященную этому вопросу (Lühmann, 1954а), с подробными гистологическими исследованиями. В результате можно считать твердо установленным, по крайней мере для атлантических зубаток, что все их виды отличаются от большинства других костистых рыб регулярной ежегодной сменой всех зубов одновременно (или почти одновременно).
Примерно через 2—3 месяца после сбрасывания старых зубов появляются закладки новых, углубляющиеся в костную ткань цоколей, которая при этом разрушается одонтокластами. Закладки обычно внедряются в бороздки, намечающие границы слившихся цоколей, вблизи их нижнего края (т. е. в места, испытывающие наименьшее давление при работе, как это справедливо отмечает Люман). С ростом зубных закладок прогрессирует и разрушение цоколей. Наконец, примерно через 7—8 месяцев после внедрения закладок прочность зубного аппарата теряется. В это время и происходит сбрасывание старых зубов, хотя вершины новых зубов еще не соприкасаются с основаниями старых. Зубы вместе с остатками цоколей сбрасываются в течение очень короткого времени. Нам не встречались особи со сброшенными старыми зубами и еще не поднявшимися новыми. Конкретных данных о таких зубатках нет и в литературе, если не считать беглого замечания Виллигби и Рея (Willoughby a. Ray, 1743 : 130) о зубатке, у которой все зубы были обломаны. После сбрасывания старых зубов происходит быстрый рост новых в высоту. Одновременно возникают и новые цоколи. Вначале они очень рыхлые, в силу чего зубы сидят непрочно. В это время зубатки не питаются или питаются лишь мягкой пищей (мелкая рыба, креветки, кишечники трески, выброшенные с траулеров). Примерно через 6—7 недель зубной аппарат становится крепким, годным к употреблению. Зубы на глоточных костях меняются, по-видимому, также все одновременно и в те же сроки, что и остальные зубы. У камбал рода Pleuronectes тоже наблюдается периодическая смена зубов — у тех видов и возрастных групп, которые питаются в основном моллюсками с твердой раковиной (Lühmann, 1955). При этом челюстные и глоточные зубы меняются одновременно, несмотря на то, что те или иные из них могут быть неприспособленными к раздавливанию панцырей добычи. Следует отметить, что у Pleuronectes специализация зубного аппарата выражена заметно слабее, чем у зубаток.
Из атлантических видов зубаток лишь Lycichthys parvodens (Lühmann, 1954а : 470) меняет зубы не одновременно, а поочередно. Описания этого вида Люман не дает, к тому же принимает без оговорок явно необоснованное деление Anarhichas (Lycichthys) latifrons на 4 вида, произведенное Гиллом. По всей вероятности, Lycichthys parvodens — всего лишь молодой экземпляр Anarhichas latifrons, подобно Lycichthys paucidens Gill. Возможно, в самом раннем возрасте Anarhichas lupus и Anarhichas minor меняют зубы также поочередно либо сохраняют какие-то следы поочередной смены зубов. Мы вправе ожидать, что вместе с общей специализацией зубного аппарата одна из ее черт — одновременность смены зубов — сдвигается в ряду Anarhichas latifrons — Anarhichas minor — Anarhichas lupus на все более ранние стадии. Замечательно, что у особей, которые, как предполагается, являются гибридами Anarhichas minor х Anarhichas lupus (Lühmann, 1954b), периодичность смены зубов нарушена. Люман считает это нарушение следствием объединения двух различно отрегулированных систем реакций: смены зубов, не связанной или слабо связанной с половым циклом (Anarhichas minor) и прочно связанной с ним (Anarhichas lupus). Но так как подобная связь для Anarhichas lupus не подтверждается (см. ниже), причину нарушения смены зубов у этих особей следует искать в чем-то ином.
Ткани новых зубов в процессе роста показывают целый ряд приспособлений к его ускорению после сбрасывания старых зубов. Эти приспособления детально рассматриваются Люманом (Lühmann, 1954а, 1955) и сравниваются с приспособлениями подобного рода у других рыб с периодической сменой зубов; к его работам и отсылаются те, кто интересуется данным вопросом более подробно.
Время, требующееся для появления закладок новых зубов, их роста и смены, указанное выше, приведено Люманом для Anarhichas lupus lupus; как сильно отличаются в этом отношении другие виды зубаток, пока по существу неизвестно. Приспособления в тканях, ускоряющие формирование новых зубов, усиливаются в ряду Anarhichas latifrons — Anarhichas minor — Anarhichas lupus lupus, но вызывают ли они действительно ускорение смены или только возрастание прочности зубов в указанном ряду, остается нерешенным.
Сроки смены зубов у отдельных особей каждого из названных видов сильно варьируют. По Люману (Lühmann, 1954b : 323), у Anarhichas lupus lupus они колеблются с ноября по февраль и даже по май-июнь, у Anarhichas minor — с апреля по декабрь; у Anarhichas latifrons, очевидно, следует ожидать еще большего размаха колебаний. Однако основная масса особей каждого из видов, надо полагать, меняет зубы в значительно более сжатые сроки: у Anarhichas lupus maris-albi примерно в сентябре-октябре, у Anarhichas lupus lupus — в декабре, у Anarhichas minor и Anarhichas latifrons — в январе-мае, причем смена зубов у Anarhichas latifrons происходит несколько позднее, чем у Anarhichas minor (Барсуков, 1954 : 897). В основном, очевидно, в декабре Anarhichas lupus lupus меняет зубы и в Баренцевом море (Барсуков, 1954 : 899) и у побережья Исландии (Lühmann, 1954b : 323). Интересно, что у Pleuronectes platessa, Pleuronectes flesus и Pleuronectes cynoglossus у побережья Исландии и в Балтийском море зубы меняются примерно в те же сроки — в ноябре-декабре (Lühmann, 1955 : 444—446). Люман считает, что у Anarhichas lupus смена зубов приурочена к определенной фазе полового цикла и происходит незадолго перед нерестом; но у Anarhichas lupus maris-albi она начинается после нереста. Мною исследовано состояние замещающих зубов у 59 особей до нереста (в июне-июле) и у 21 особи после нереста (начало сентября). Длина особей от 21 до 66 см. Заметных возрастных и половых различий в смене зубов не обнаружено. По-видимому, у всех зубаток смена зубов приурочена к началу сезона, наименее благоприятного в отношении питания; здесь не последнюю роль могут играть изменения температуры воды, глубины, течения и т. д.
Закладки всех зубов у Anarhichas lupus, как и у других видов зубаток, имеют вначале коническую форму, но у давящих зубов, по Шмидту (Schmidt, 1954 : 37), основание конуса в дальнейшем расширяется, вершина же сохраняется в виде сосочка на округлой верхней поверхности зуба. Эти сосочки остаются первое время на уже сменившихся и даже окрепших зубах, но затем скоро стираются. Вряд ли они имеют какое-либо функциональное значение: прорезывание зуба обеспечивается гораздо более основательными приспособлениями (в виде разрыхления тканей над всей его верхней поверхностью и т. д. — Lühmann, 1954а : 485). Таким образом, взаимодействие тканей при закладке давящих зубов приводит первоначально к образованию закладок остроконических схватывающих зубов, лишь впоследствии видоизменяясь. Очевидно, начальный этап развития закладок обусловлен прежней, ныне угасшей функцией указанных зубов; по всей вероятности, у предков зубаток, как и у личинок, все зубы были острыми, служили только для схватывания добычи, как это рассматривает и Шмидт, правда с другой точки зрения.
Одновременность смены всех зубов у зубаток объясняется приспособлением зубного аппарата к раздавливанию организмов с твердым панцирем; точнее, в развитие указанной мысли Люмана, к отрыванию их от субстрата (клыки), разрыванию (конические), раздавливанию (давящие) и выскребанию мягких частей из обломков (глоточные). Во всех зтих случаях зубы испытывают значительно большее давление на основание, чем при схватывании добычи, лишенной панциря, а отсюда увеличивается угроза повреждения новых зубов при их нерегулярной смене.
Ряд особенностей в строении тканей зуба (ячеисто-трубчатый мантельдентин, слабое обызвествление), по Люману, направлен против опасности раскалывания зубов при раздавливании твердой добычи, а также против соскальзывания ее; но те же самые особенности обусловливают сильную снашиваемость зубов, а следовательно, и необходимость их частой смены. Эта снашиваемость иногда еще усиливается вследствие проникновения паразитического микроорганизма «Mycelites ossifragus» (Schmidt, 1954), развитию которого, возможно, благоприятствует «рыхлое» строение дентина.
Частая одновременная смена зубов обусловливает не более чем двурядное их расположение на тех местах, где зубы несут достаточную нагрузку. Необходимость противостоять значительным силам, стремящимся сломать, раздавить и сдвинуть зубы, требует слияния цоколей и тесного соприкосновения оснований зубов, а у давящих зубов также и боковых поверхностей. Уже при одном только слиянии цоколей смена зубов, окруженных со всех сторон другими зубами, затрудняется: закладки новых зубов при этом могут внедряться в ткань цоколей лишь сверху, между основаниями старых зубов, но не сбоку. О том, что в первом случае смена зубов действительно затруднена, свидетельствуют следы неудавшихся, прекративших развитие закладок (Lühmann, 1954а; собственные наблюдения). Сильное сокращение и исчезновение промежутков между основаниями зубов делает окончательно невозможным расположение зубов более чем в два ряда. В один-два ряда они и расположены на большей части костей, несущих зубы, у большинства особей всех видов зубаток. У предков зубаток расположение зубов на всех этих костях было, по-видимому, многорядным, с неправильными, сбитыми рядами, сходно с молодью Anarhichas latifrons, имеющей наиболее примитивное строение зубов. Включение зубов между обычными двумя рядами, иногда встречающееся у взрослых особей атлантических видов зубаток, очевидно, следует рассматривать как атавизм; в норме оно происходит лишь на глоточных костях, исключая переднюю верхнеглоточную.
Смена зубов у тихоокеанских зубаток, по-видимому, ничем существенным не отличается от таковой же у атлантических (Барсуков, 1954 : 899).
В заключение остается сказать несколько слов о «губах» зубаток — довольно толстых складках кожи, окаймляющих рот. Из них верхняя поддерживается связкой, идущей вдоль нижнего края межчелюстной кости и натягивающейся от конца ее горизонтальной ветви к заднему концу верхнечелюстной кости. Нижняя губа поддерживается стержнем из плотной соединительной ткани, похожей на хрящ. Основание стержня прикрепляется к пережиму нижнечелюстной кости, ниже цоколей зубов, а вершина переходит в связку, прикрепляющуюся тоже к заднему концу верхнечелюстной кости. Стержень расположен таким образом, что обеспечивает обтекаемый обвод нижней части головы, ликвидируя вредное действие пережима нижнечелюстной кости.

Примечания. 1) В этой и следующих таблицах цифры, выделенные курсивом, указывают количество экземпляров. 2) Для Anarhichas latifrons измерялись особи длиной более 100 см.
У всех зубаток губы устроены довольно сходно, но у Anarhichas latifrons задняя часть нижней губы выдается вверх в виде клапана, которому соответствует небольшое углубление на верхней губе. Возможно, этот клапан препятствует выскальзыванию изо рта гребневиков и медуз, составляющих обычный объект питания синей зубатки. У всех зубаток поверхность губ спереди, перед клыками, покрыта складками, похожими на бахрому, но у Anarhichas latifrons вместо бахромок развиты хорошо выраженные сосочки, а сами губы гораздо толще (табл. 2).
По-видимому, эта особенность в строении губ вызвана большей их ролью в отыскании пищи. Последнее может быть вызвано уменьшением роли зрения в связи с обитанием на наибольших глубинах по сравнению с другими зубатками. Нижняя губа у Anarhichas latifrons значительно тоньше верхней, притом нижняя челюсть и без того несколько укорочена. Укорочение нижней челюсти и преимущественное развитие верхней губы свидетельствуют о том, что взрослые особи Anarhichas latifrons берут пищу преимущественно со дна. У молоди нижняя челюсть не столь укорочена, губы тоньше, верхняя губа по толщине меньше превышает нижнюю, сосочки развиты слабее — и все это несмотря на большие глубины обитания, чем у взрослых; таким образом, доставляется еще одно доказательство меньшей роли донной фауны в питании молоди. Не исключена возможность, что молодь Anarhichas latifrons ведет батипелагический образ жизни и у дна встречается реже, чем взрослые особи.
Дополнение к главе «Морфология», раздел «Зубы». У пелагических мальков атлантических видов зубаток все зубы острые, обычно с загнутыми назад вершинами. Относительная величина зубов увеличивается в ряду Anarhichas latifrons — Anarhichas minor — Anarhichas lupus lupus; по-видимому, для этого же ряда характерно увеличение относительных размеров и подвижности пищевых организмов. На площадках межчелюстных и нижнечелюстных костей сидят два ряда зубов; на нёбных костях, а также на нижнечелюстных за пережимом — лишь один ряд. У мальков Anarhichas latifrons ряды зубов на соседних praemaxillaria подходят друг к другу под более тупым углом, чем у Anarhichas minor и Anarhichas lupus lupus, а пережим на dentale развит слабее. Расстояние от заднего зуба сошника до уровня задних зубов нёбных костей у мальков Anarhichas latifrons превышает длину сошникового ряда зубов примерно в два раза, у мальков Anarhichas minor — равно длине сошникового ряда или превышает ее, но не более чем в полтора раза, у мальков Anarhichas lupus lupus — меньше половины этой длины. Изменения в форме и расположении зубов наиболее интенсивны при переходе к питанию организмами с прочным панцирем и происходят у всех трех видов в общем параллельно. Одно из наиболее резких отклонений от этого параллелизма наблюдается у Anarhichas latifrons: как подтвердилось на материале, собранном в марте 1959 г., для особей данного вида длиной по крайней мере от 67 и до 75—80 см действительно характерна многорядность зубов на всех костях, несущих зубы, — в отличие от мальков длиной 2.5 см и от особей крупнее 75—80 см. Возможно, указанный этап развития зубной системы у других зубаток выпал вследствие гораздо более раннего перехода к питанию организмами с прочным панцирем. Изменения в строении зубной системы Anarhichas latifrons, связанные с частичным переходом к питанию организмами с прочным панцирем, происходят в основном при длине 7—115 см. В это время особи одной и той же длины нередко сильно отличаются друг от друга по степени изменений в зубной системе, что может быть связано с различиями в темпе их роста.
При смене зубов, по крайней мере у Anarhichas latifrons и Anarhichas minor, вначале выпадают старые зубы, легко отделяющиеся в это время от цоколей; закладки новых зубов, основания которых расположены в «альвеолах» (в углублениях, разъеденных в кости одонтокластами), первое время защищены сверху остатками старых цоколей. Затем выпадают и эти остатки; ткань новых цоколей затягивает «альвеолы», подрастая в высоту вместе с основанием нового зуба.
С помощью глоточных зубов Anarhichas latifrons иногда издает скрипящие (скрежещущие) звуки, обычно при закрытом рте. Такие же звуки издают и другие виды зубаток — возможно, тем же образом.
Мышцы и скелет
Здесь будет рассмотрено своеобразие в строении мышц головы и в строении черепа зубаток, которое связано с питанием организмами с твердым панцирем. Влияние движения и ряда других функциональных особенностей на строение мышц тела, осевого скелета и черепа удобнее рассмотреть в соответствующих разделах, касающихся формы тела и органов чувств.
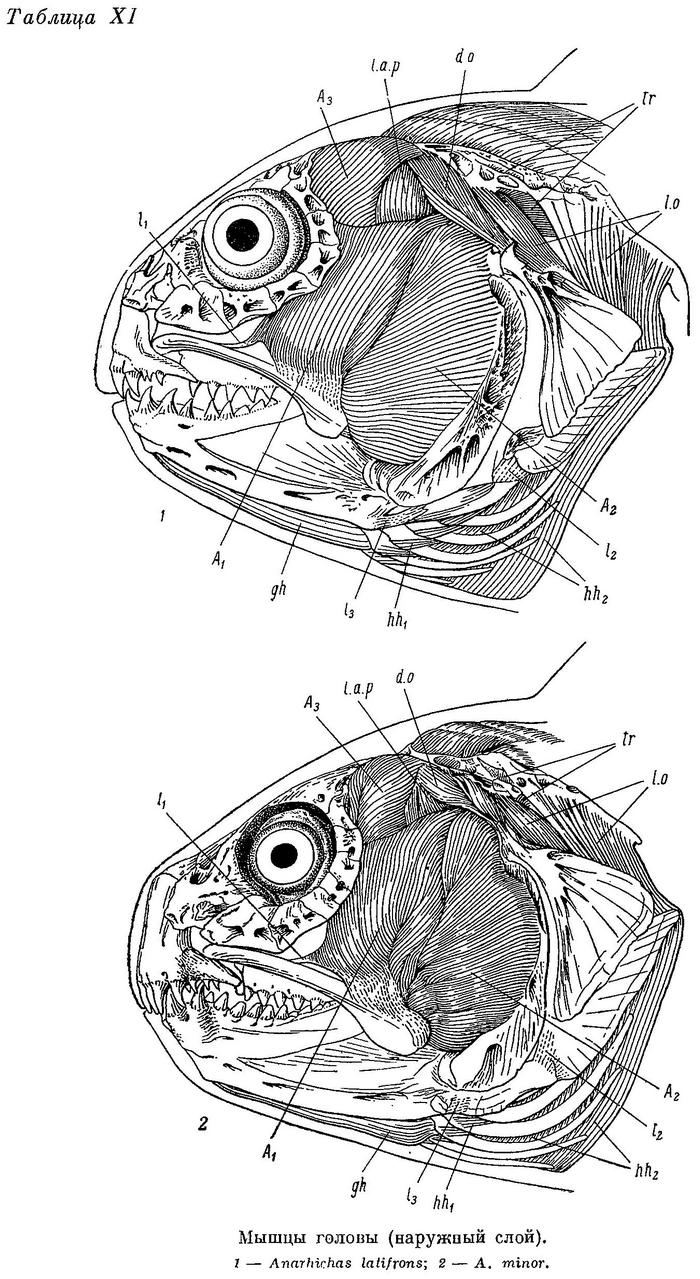
Дробящий зубной аппарат зубаток требует прежде всего приложения большой силы, сжимающей челюсти. Поэтому особенно интересно строение жевательных мышц (m. adductor mandibulae), состоящих у зубаток (табл. XI—XII), как и у многих других костистых рыб, из трех основных порций — А1, А2, А3 и челюстной порции Aw. Обозначения мышц даны по Феттеру (Vetter, 1878).
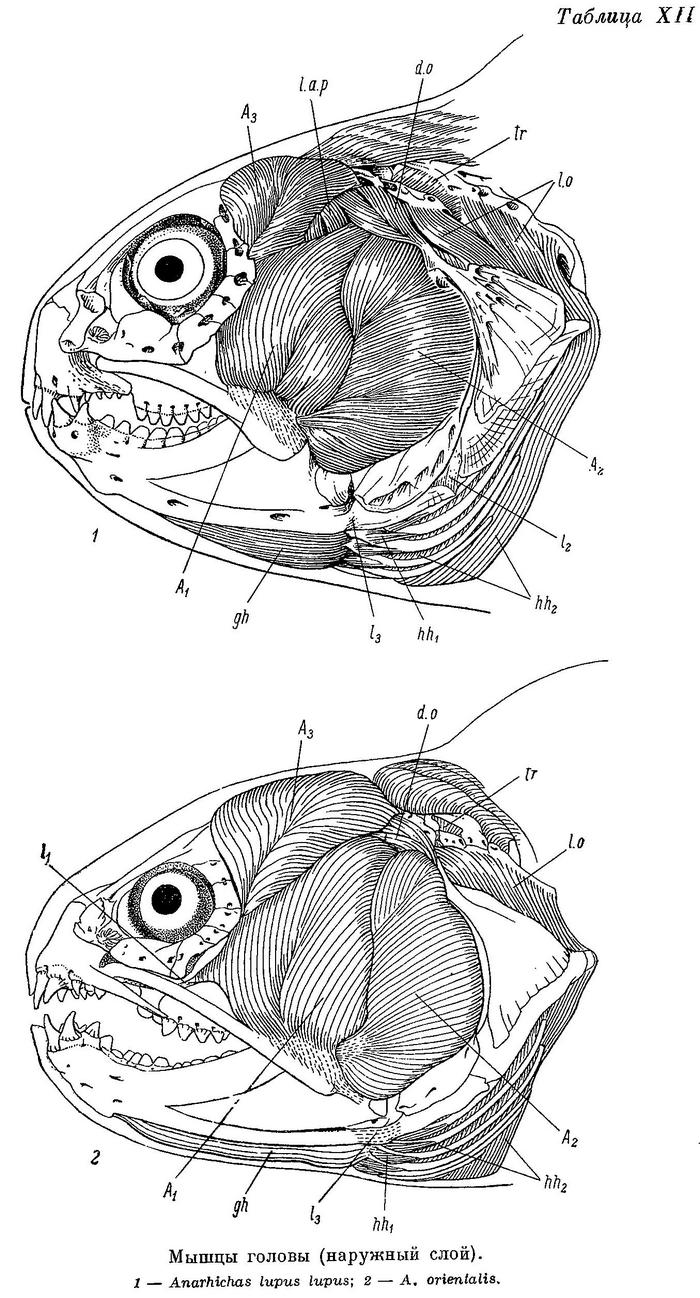
А1 — наиболее слабая из основных порций (17—24% от их общего веса), имеет два места прикрепления к неподвижным костям (punctum fixum): передняя часть ее прикрепляется ко второй сзади suborbitale, обычно утолщенной и выдающейся немного назад, задняя — к гребню на pteroticum, расположенному над суставной канавкой для hyomandibulare, и к срединному гребню на hyomandibulare вниз до отверстия, через которое выходит r. hyomandibularis n. facialis (табл. XIII—XIV). Фасция А1 прикрепляется к внутренней поверхности suborbitalia от второй сзади до первой-второй спереди. Между верхними краями этих двух участков натянута прочная связка, к которой прикрепляется часть волокон данной порции, а также небольшая часть волокон m. levator arcus palatini. Мест прикрепления к подвижным костям (punctum mobile) также два: на верхней и нижней челюстях. Сухожилия передней части А1 прикрепляются к верхнему краю заднего конца maxillare и к наружной стороне короноидного отростка articulare. Сухожилия задней части А1 прикрепляются частично тоже к наружной стороне articulare, рядом с сухожилиями передней части, но в основном уходят внутрь, в выемку нижней челюсти, сливаясь здесь с сухожилиями от порций А2 и А3. Последние прикрепляются вблизи стержневидного остатка меккелева хряща — от сесамоидной косточки, возле заднего конца хряща, до переднего края выемки, примерно на уровне пережима dentale. По Бергу (1940 : 237—238, фиг. 129), это остаток меккелевой кости; в той же работе приводятся иные мнения и названия. Сесамоидную косточку Макушок (1958 : 63) именует «pararticulare» и приводит также два других названия. От нижнего края А1 у всех зубаток, кроме Anarhichas lupus, отходит спереди очень длинное, тонкое, но весьма прочное сухожилие (табл. XI—XII l1), прикрепляющееся к внутренней поверхности головки maxillare под связкой, натянутой между головкой и передним краем palatinum.

А2 расположена кзади от А1 и перекрывается последней лишь у личинок; составляет 26—32% от общего веса основных порций. Punctum fixum А2 охватывает задний угол pteroticum, заднюю половину hyomandibulare, почти всю praeoperculum (исключая задний нижний угол), свободную часть symplecticum и задний отросток quadratum. Punctum mobile располагается на заднем крае короноидного отростка articulare с внешней стороны (табл. XIII—XIV). Кроме того, широкое сухожилие идет от переднего края А2 внутрь, в выемку нижней челюсти (см. выше).
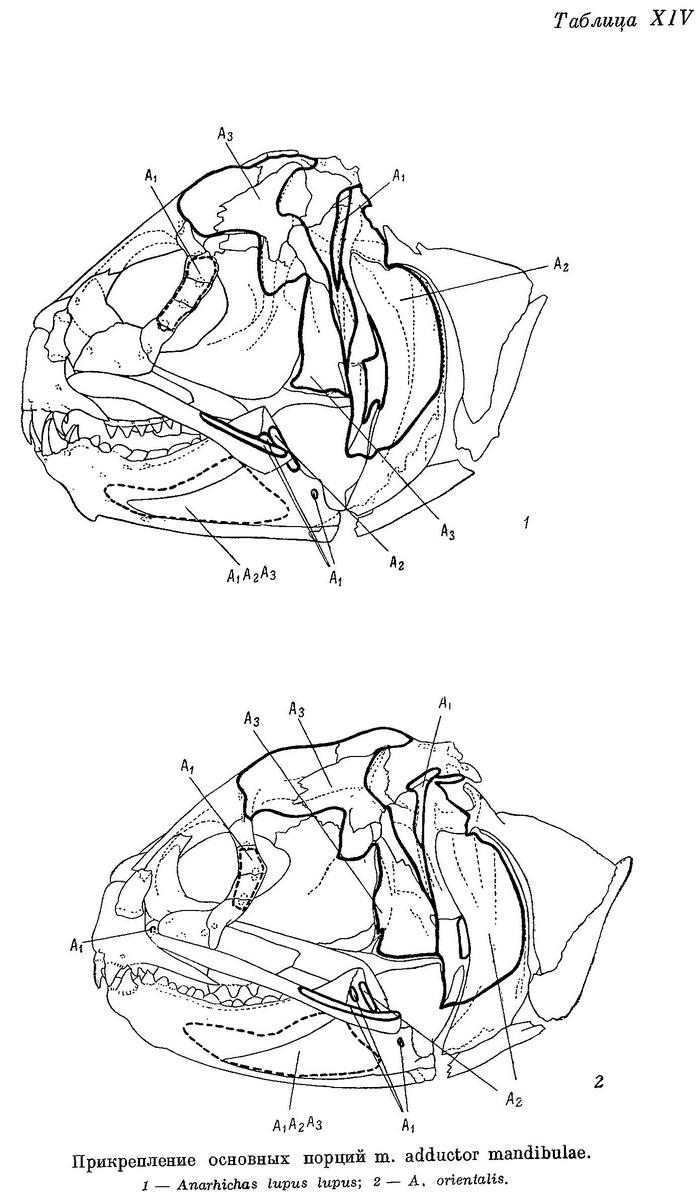
А3 — наиболее мощная из всех порций (48—55% от их общего веса). Нижние части ее накрыты порциями А1 и А2. Punctum fixum этой порции охватывает боковую поверхность frontale за заглазничными отростками, боковую поверхность sphenoticum до гребня, небольшой участок prooticum перед сочленовной поверхностью с hyomandibulare, боковую поверхность крыльев парасфеноида, передний край hyomandibulare и наружную поверхность метаптеригоида (табл. XIII—XIV). В самой нижней части области прикрепления волокна А3 слабо отделены от волокон А2 (в остальных местах все порции отделены друг от друга четко). Punctum mobile располагается в основном в области сесамоидной косточки. К ней прикрепляется задняя часть широкого сухожилия от А3, передняя же часть срастается с сухожилиями от А1 и А2, уходящими в глубь выемки в нижней челюсти.
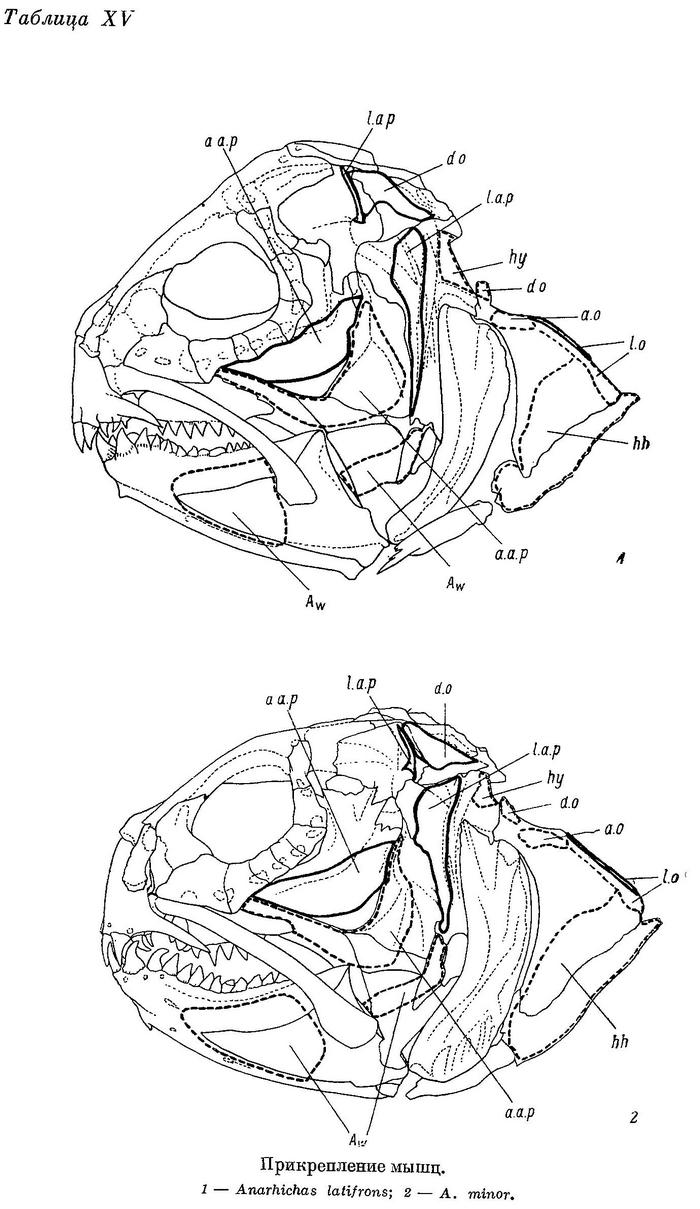
Aw — по весу в несколько раз меньше каждой из основных порций. Ее волокна занимают все свободное место в выемке нижней челюсти — от сесамоидной косточки до переднего конца выемки. Лишь самые поверхностные волокна прикрепляются к собственному сухожилию, идущему вдоль внутренней поверхности quadratum и прикрепляющемуся к внутренней поверхности symplecticum, а также к гребню на внутренней стороне заднего отростка quadratum (табл. XV—XVI). Остальные волокна прикрепляются к сросшимся сухожилиям основных порций. Волокна Aw покрыты очень прочной фасцией, натянутой между верхней и нижней ветвями dentale.
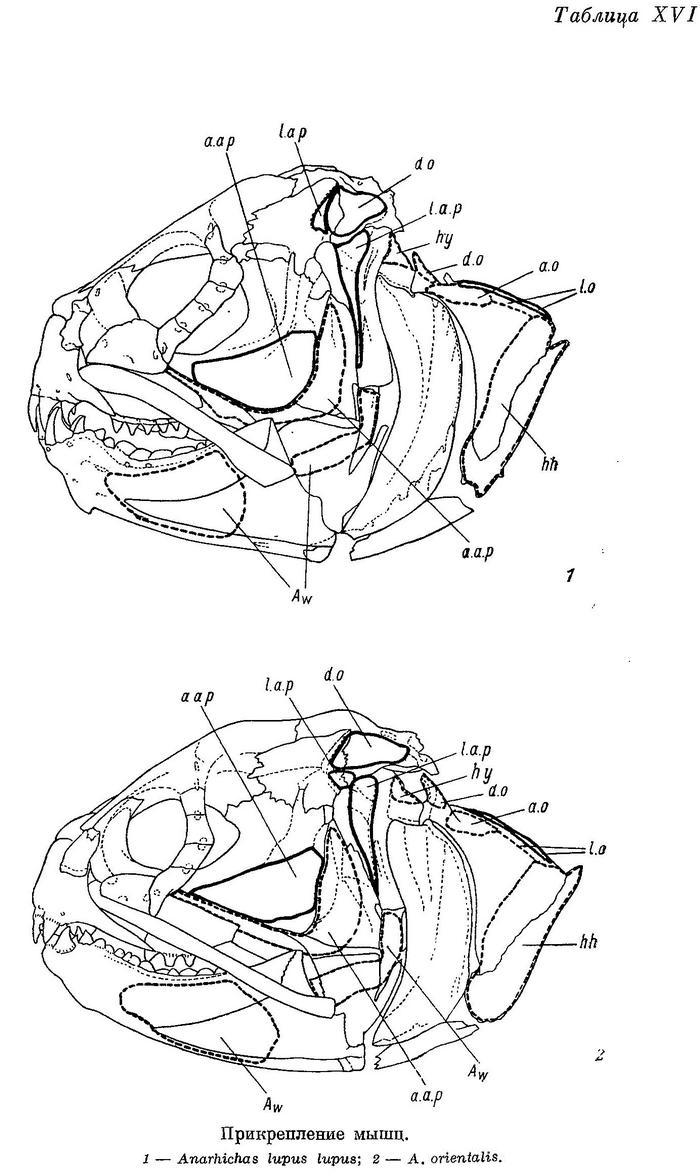
Для зубаток наиболее характерны следующие особенности в строении m. adductor mandibulae: 1) эти мышцы развиты необычайно сильно (составляют 1—6% от веса всего тела); 2) угол, под которым расположены их порции по отношению к нижней челюсти (вид сбоку), приближается к прямому; он особенно близок к прямому у А3 — самой мощной порции; 3) области прикрепления всех трех основных порций распространяются на неврокраниум, являющийся более прочным, чем висцеральный скелет; 4) более всего простирается на неврокраниум область прикрепления А3, что сопровождается сужением черепа с боков, оттеснением sphenoticum от глазницы и разрывом области прикрепления А1 на два участка. Все эти приспособления обеспечивают большую силу сжатия челюстей, необходимую для раздавливания прочных панцирей пищевых организмов. Непонятно лишь сужение черепа с боков в области прикрепления А3, так как при этом угол, который образует А3 по отношению к нижней челюсти, становится менее выгодным, отклоняясь от прямого (вид спереди). В чем же тут дело?
Как уже было сказано, у зубаток зубы на сошнике приходятся между нижнечелюстными костями, к тому же давящие зубы на одной из них наклонены навстречу зубам другой. Естественно, что при таких условиях во время раздавливания твердых панцирей пищевых организмов наряду с силами, противодействующими сближению нижней и верхней челюстей, возникают весьма значительные силы, разводящие нижнечелюстные, а отчасти и нёбные кости (рис. 3). Мышцами, которые могут противодействовать разведению нижнечелюстных костей, являются m. intermandibularis, m. adductor arcus palatini и m. hyomandibularis.
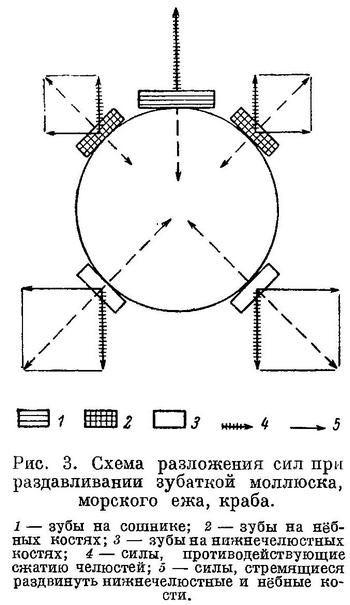
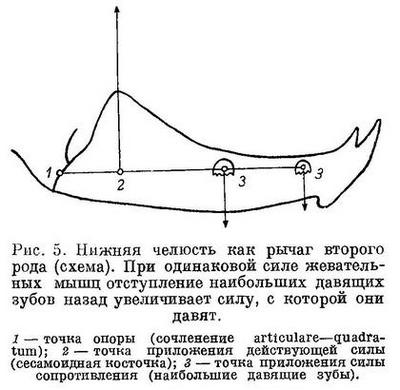
М. intermandibularis протянут в виде широкой ленты между нижнечелюстными костями, прикрепляясь к каждой из них от места их сочленения друг с другом до края выемки. Он всегда располагается под наибольшими давящими зубами и несомненно играет немалую роль в этом противодействии. Однако развитие m. intermandibularis сильно ограничено выемками в нижнечелюстных костях, необходимыми для увеличения силы сжатия челюстей. Нижняя челюсть представляет собой рычаг второго рода, где точкой опоры является сустав articulare—quadratum, точкой приложения действующей силы — место прикрепления сухожилий основных порций m. adductor mandibulae, точкой приложения силы сопротивления (при раздавливании) — давящие зубы. Развитие выемки в нижней челюсти приближает точку приложения действующей силы к точке приложения силы сопротивления, что делает рычаг более выгодным.
М. adductor arcus palatini развит сильно. Punctum fixum зтой мышцы располагается на боковой стороне киля парасфеноида (табл. XV—XVI), punctum mobile охватывает внутренние стороны эндоптеригоида и метаптеригоида. Этот киль сокращает высоту ротовой полости зубаток, что, однако, не мешает им заглатывать добычу значительных размеров: Anarhichas lupus maris-albi, например, может заглатывать рыбу длиной до 63% от своей собственной (при близкой форме тела). Основной функцией правого и левого m. adductor arcus palatini является противодействие раздвиганию нёбных костей при раздавливании организмов с прочным панцирем. Эти мышцы могут противодействовать раздвиганию нижнечелюстных костей лишь косвенно, через сустав articulare—quadratum.
М. hyomandibularis развит слабо. Его punctum fixum располагается на боковой поверхности pteroticum, ниже сочленения с hyomandibulare, punctum mobile — на заднем верхнем углу hyomandibulare, между срединной осью и оперкулярным отростком (табл. XV—XVI, XVIII). Роль этих мышц в предупреждении раздвигания челюстей несомненно ничтожна.

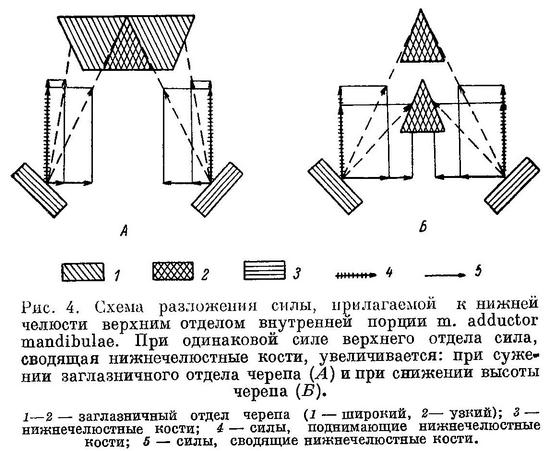
Всех указанных выше мышц (двух парных и одной непарной), надо полагать, недостаточно для противодействия раздвиганию челюстей при раздавливании наиболее прочных панцирей пищевых организмов. Если это так, mm. adductor mandibulare одновременно с силами, сближающими нижнюю и верхнюю челюсти, должны развивать силы, сближающие правую и левую нижнечелюстные кости. Эти силы будут тем больше, чем сильнее сближены punctum fixum левого и правого m. adductor mandibulae, расположенные на неврокраниуме, по сравнению с punctum mobile данных мышц (рис. 4, А); чем ниже череп в области этих punctum fixum, (рис. 4, Б), чем ближе к прямому (вид сбоку) угол между линией punctum fixum—punctum mobile и нижней челюстью. Отсюда понятно, почему именно внутренние порции могут наилучшим образом обеспечить сведение нижнечелюстных костей одновременно с поднятием их вверх, перенеся свои punctum fixum на неврокраниум: у этих порций легче всего может быть достигнуто сближение punctum fixum противолежащих сторон по сравнению с punctum mobile. Необходимостью такого сближения и объясняется сильное сужение неврокраниума в области прикрепления верхних отделов A3. Верхним отделом А3 названы волокна внутренней порции m. adductor mandibulae, прикрепляющиеся к неврокраниуму, нижним — волокна, прикрепляющиеся к нёбной дуге. У Azygopterus и Xiphister, относящихся к тому же надсемейству Stichaeoidae, что и зубатки, сильно развитые жевательные мышцы тоже заходят на крышу черепа, что сопровождается сужением его заглазничной части. Следовательно, эти рыбы также должны затрачивать большую силу на сведение нижнечелюстных костей одновременно с их поднятием. Однако зубная система у Azygopterus и Xiphister не обнаруживает никаких признаков специализации к питанию организмами с твердым панцирем (Макушок, 1958 : 17, 51). Остается предположить, что сведение нижнечелюстных костей необходимо этим рыбам для другой цели — а именно для мощного сжатия ротовой и околожаберных полостей одновременно. Возможно, проталкивание воды через жаберные лепестки и тычинки требует большей силы, чем у других Stichaeoidae, что может быть связано с какими-то особенностями питания и (или) дыхания. Выяснение этого вопроса, может быть, позволило бы лучше понять особенности строения ротового аппарата у предков зубаток, явившиеся исходными для современных представителей семейства. Понятна отсюда и основная роль порций А3 при раздавливании, а следовательно, и их более мощное развитие. Сила, с которой А3 и поднимают и сводят нижнечелюстные кости, будет тем большей, чем ближе к прямому (вид сбоку) угол их расположения по отношению к нижнечелюстным костям. Поэтому верхние отделы А3 могли переместиться с нёбной дуги на неврокраниум только вперед, разрывая первоначально, вероятно, единые области прикрепления каждой A1. Снижение высоты черепа впереди нёбной дуги в свою очередь увеличило долю силы, сводящей нижнечелюстные кости, в общей равнодействующей.
Вследствие сильного развития верхнего отдела А3 m. levator arcus palatini оттесняется назад, что должно было бы вызвать также оттеснение m. dilatator operculi, расположенного сзади m. levator arcus palatini. Однако оттеснение m. dilatator operculi назад затруднено, поскольку угол расположения указанной мышцы по отношению к жаберной крышке становится при этом менее выгодным. Выходом из данного противоречия является образование сильно развитого гребня на sphenoticum, к вершине которого и прикрепляется m. levator arcus palatini. Приподнятость punctum fixum, кроме того, делает угол действия m. levator arcus palatini более выгодным. Образование гребня на sphenoticum увеличивает также площадь прикрепления верхнего отдела порции А3.
Чем меньше промежуток между нижнечелюстными костями в области давящих зубов, тем успешнее должно быть раздавливание моллюска (при большом промежутке могут обламываться лишь его края), тем меньше вероятность его проскальзывания между этими костями. Площадка же клыков должна быть достаточно широкой для того, чтобы обеспечить успешное отрывание моллюска от субстрата. Отсюда становится понятным развитие пережима на нижнечелюстных костях, столь характерного для зубаток.
В связи с сильным развитием зубов и жевательных мышц у зубаток увеличивается мощность всех костей черепа, особенно костей, вооруженных большими зубами, — нижнечелюстных, вертикального отростка межчелюстных, сошника, нёбных — и костей, их поддерживающих. Поверхности костей, несущие цоколи зубов, укрепляются с краев утолщениями костной ткани. Такие утолщения лучше выражены под большими клыками и наибольшими давящими. Утолщения под клыками выдвигают их вперед, что облегчает отрывание моллюска от субстрата. Утолщения под давящими сближают давящие правой и левой нижнечелюстных костей, усиливая действие их пережимов. Кости неврокраниума, выдерживающие сильную тягу со стороны прикрепленных к ним порций m. adductor mandibulae и давление со стороны костей, несущих зубы, соединены между собой очень крепко (у Anarhichas lupus maris-albi они не разваливаются даже у сваренной зубатки). Suborbitalia, к которым прикрепляется порция А1, приобретают у зубаток, в отличие от других Stichaeoidae, жесткое соединение (Макушок, 1958 : 48).
Интересно, что метаптеригоид, служащий местом прикрепления двух мощных мышц — части порции А3 снаружи и части m. adductor arcus palatini изнутри — тем не менее представляет собой тонкую пластинку. Это свидетельствует об одновременности действия указанных мышц.
Следует остановиться на работе выдвижного аппарата рта у зубаток, которую я представляю себе несколько иначе, нежели Ван-Доббен (Van Dobben, 1937). Верхнечелюстная кость имеет две сочленовных головки: одной из них она прикрепляется к переднему краю сошника, другой — к внутренней стороне межчелюстной кости. При открывании рта связки поворачивают верхнечелюстную кость таким образом, что ее сочленовная головка с межчелюстной костью поднимается вверх; соответственно выдвигается вверх и вперед и межчелюстная кость. Rostralia, представляющие собой хрящевые подушечки, приращенные к суставной поверхности вертикального остростка межчелюстной кости, скользят при этом по передней, суставной поверхности мезэтмоида вверх и вперед. При закрывании рта верхнечелюстная кость поворачивается в обратном направлении действием переднего отдела порции А1 — межчелюстная кость при этом движется вниз и немного внутрь, навстречу нижней челюсти. Способность к выдвиганию praemaxillare у предков зубаток, возможно, была развита лучше, чем у современных видов, однако и у последних она не утрачена, вопреки утверждению Макушка (1958 : 51). При изучении зубаток между мною и В. М. Макушком возникли разногласия, многие из которых можно было бы устранить в процессе совместной работы; однако в течение последних трех лет нам пришлось работать порознь в Антарктической экспедиции, не имея между собою достаточной связи. В этих разногласиях, приведенных здесь и ниже, повинны мы оба, но я нахожусь в более выигрышном положении, поскольку моя работа выходит позднее.
Submaxillare у зубаток представлено лишь соединительнотканной подушечкой; значительной роли при выдвижении межчелюстной кости оно играть не может. Выдвижной аппарат рта у зубаток, таким образом, стоит на довольно низкой ступени развития (Еремеева, 1948 : 134 и сл.).
Роль длинного сухожилия, идущего от переднего края А1 к головке верхнечелюстной кости, неясна. Возможно, оно устраняет смещение вверх переднего конца верхнечелюстной кости при опускании межчелюстной. Отчасти такое смещение устраняется мощным praeorbitale (lacrimale), а также передним отростком нёбной кости, удлиняющимся и перегибающимся над передним концом верхнечелюстной; последнее лучше всего выражено у Anarhichas lupus. Может быть, в связи с этим сухожилие, упомянутое выше, у Anarhichas lupus исчезает. У Anarhichas latifrons оно развито слабее, чем у Anarhichas minor и Anarhichas orientalis, а у одной особи Anarhichas latifrons совсем не было обнаружено. Эта редукция, по-видимому, связана с утратой значения выдвижного аппарата ввиду укорочения нижней челюсти.
Как видно из вышеизложенного, А1, поднимая нижнюю челюсть, одновременно опускает верхнюю. Очевидно, именно эту порцию жевательных мышц зубатка вводит в действие при отрывании пищевого организма от субстрата. Расположение А1 под прямым углом к челюстям, впереди А2, выгодно в отношении силы действия указанной порции, а значительной скорости от нее не требуется. При схватывании более подвижных организмов, по всей вероятности, употребляется лишь А2, волокна которой расположены под наибольшим углом к нижней челюсти и, следовательно, обеспечивают большую скорость ее закрывания. Сближения нижнечелюстных костей в обоих случаях не требуется, а поэтому punctum fixum А1, а также А2 правой и левой сторон далеко расставлены друг от друга. Наконец, при раздавливании организмов с твердым панцирем пускаются в ход все три порции, из которых наибольшую роль играет А3. Эти предположения безусловно требуют экспериментальной проверки.
Рассмотрим теперь видовые различия в строении жевательных мышц и связанных с ними костей.
Как и следовало ожидать, исходя из особенностей строения зубного аппарата, в ряду взрослых особей Anarhichas latifrons — Anarhichas minor — Anarhichas lupus lupus — Anarhichas lupus maris-albi — Anarhichas orientalis сила m. adductor mandibulae, судя по увеличению относительного веса этой мышцы, возрастает (табл. 3).

Угол основных порций m. adductor mandibulae, в том числе и А2, по отношению к нижней челюсти в указанном ряду взрослых особей понемногу приближается к прямому (вид сбоку), что особенно заметно при переходе от Anarhichas latifrons к Anarhichas minor. Очевидно, в том же порядке уменьшается и роль подвижных организмов в питании, как на то уже указывало строение зубной системы. Приближение угла, под которым расположены порции m. adductor mandibulae по отношению к нижней челюсти, к прямому увеличивает силу сжатия челюстей, но уменьшает скорость их закрывания. Известно, что укус полосатой зубатки относительно медленный, сдавливающий (Lühmann, 1954а : 470; собственные наблюдения). Л. Д. Марцинкевич исследовала микроскопическое строение m. adductor mandibulae у баренцевоморской Anarhichas minor длиной 54 см и Anarhichas lupus lupus длиной 45 см. Эта мышца у Anarhichas minor содержит всего лишь около 7% красных мышечных волокон, у Anarhichas lupus lupus — около 25%. Остальную массу мышцы составляют белые волокна. Как известно, нетонические (белые) волокна быстро сокращаются, но столь же быстро утомляются; тонические (красные), напротив, совершают длительную работу (Литвер, 1948 : 127; Колосс, 1957 : 704). Таким образом, скорость схватывания добычи может у Anarhichas lupus уменьшаться не только вследствие несколько иного расположения «жевательных» мышц, но и вследствие иного их состава. Впрочем, функция красных мышечных волокон у рыб в настоящее время не может еще считаться вполне ясной (Braekkan, 1956 : 747), а поэтому их значение в общей работе мышц может оказаться более сложным. Мышечные волокна m. adductor mandibulae у Anarhichas lupus lupus, несмотря на большую массу ее мышц, тоньше, чем у Anarhichas minor (27—35 и 35—56 μ, соответственно); интересно было бы исследовать возрастные изменения этого различия.
В ряду Anarhichas latifrons — Anarhichas minor — Anarhichas lupus lupus — Anarhichas lupus maris-albi — Anarhichas orientalis наблюдается постепенный сдвиг наибольших давящих зубов назад (табл. II, 1—6). Расстояние от них до центра прикрепления сухожилий m. adductor mandibulae уменьшается по сравнению с расстоянием от этого центра до сустава articulare—quadratum. Следовательно, даже при одинаковой силе m. adductor mandibulae давящая сила челюстей увеличивается вследствие более выгодного соотношения плечей рычага (рис. 5); но по той же самой причине увеличивается и сила, раздвигающая нижнечелюстные кости. Более того, в описанном ряду увеличивается наклон давящей поверхности зубов внутрь, что должно особенно увеличивать раздвигающую силу. Вес мышц, противодействующих этой силе — m. intermandibularis, m. adductor arcus palatini, m. hyomandibularis — по отношению к весу m. adductor mandibulae должен, казалось бы, возрастать. В действительности наблюдается совершенно обратное (табл. 4).

Следовательно, должна нарастать роль верхних отделов порций А3 в сведении нижнечелюстных костей. Действительно, вес верхнего отдела А3, прикрепленного к неврокраниуму, возрастает по сравнению с весом нижнего отдела, прикрепленного к нёбной дуге (табл. 5).

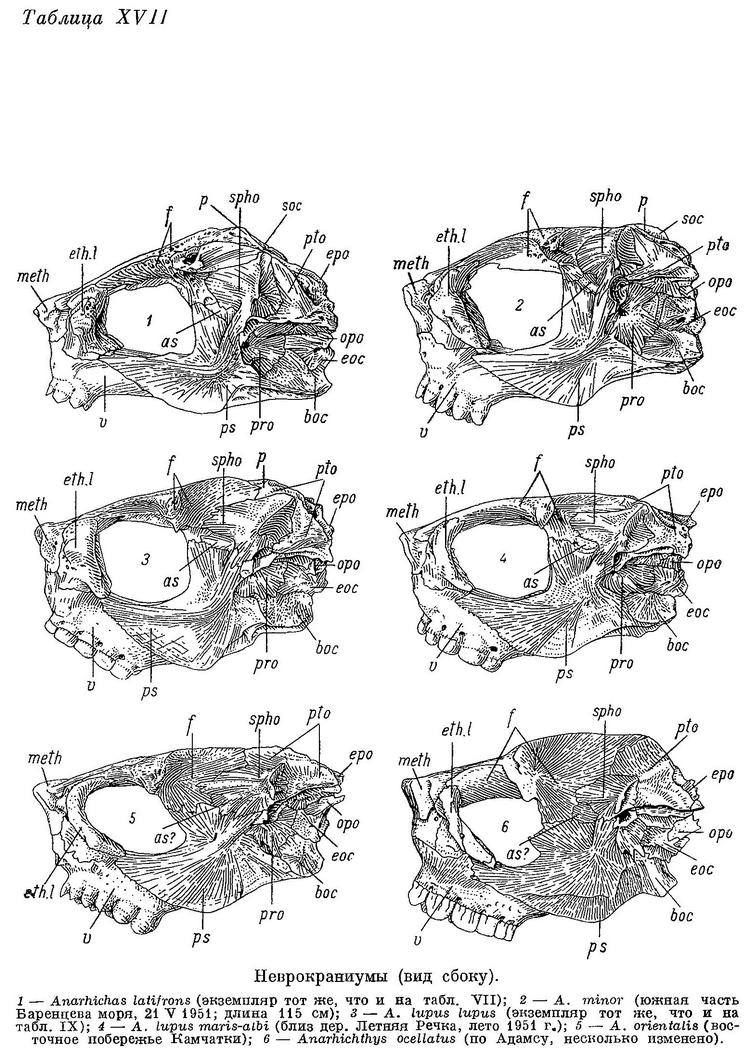
Та часть неврокраниума, где прикрепляются верхние отделы А3, сильно суживается (табл. XIX—XX), а высота черепа уменьшается (табл. VII—X). Увеличиваются площади прикрепления этих отделов, в связи с чем sphenotica сдвигаются назад и вниз (табл. XVII—XVIII). Punctum fixum m. levator arcus palatini постепенно вытесняется в нижнюю часть гребня sphenoticum, оттягивающуюся вниз, что до известной степени компенсирует вредное влияние сужения неврокраниума на угол действия указанной мышцы.
Прочность костной ткани в ряду Anarhichas latifrons — Anarhichas minor — Anarhichas lupus — Anarhichas orientalis заметно увеличивается. Мощность ее утолщений под цоколями больших клыков и давящих возрастает. Площадь сочленовных площадок между dentalia увеличивается; нижний край этих площадок смещается назад, поэтому m. intermandibularis находится всегда под наибольшими давящими, что облегчает противодействие раздвиганию нижнечелюстных костей при раздавливании моллюска.
Укрепление окологлазничного кольца происходит не только за счет увеличения мощности praeorbitale и suborbitalia, но и за счет уменьшения числа suborbitalia (табл. 6).

У трех особей (Anarhichas latifrons, Anarhichas minor, Anarhichas lupus lupus) было обнаружено различное число suborbitalia на разных сторонах головы. Во всех этих случаях большее число suborbitalia наблюдалось на той стороне, где А1 прикрепляющаяся частично к окологлазничному кольцу, была слабее, судя по ее меньшему весу; слабее оказывались и другие порции m. adductor mandibulae.

Автору не представилось возможности исследовать мышцы Anarhichthys ocellatus. Однако особенности ее черепа (табл. X, 2, XVII, 6, XIX, 6, XXI, 6) не вызывают сомнения в том, что и по строению мышц этот вид является крайним членом описанного ряда зубаток.
перейдем теперь к возрастной изменчивости.
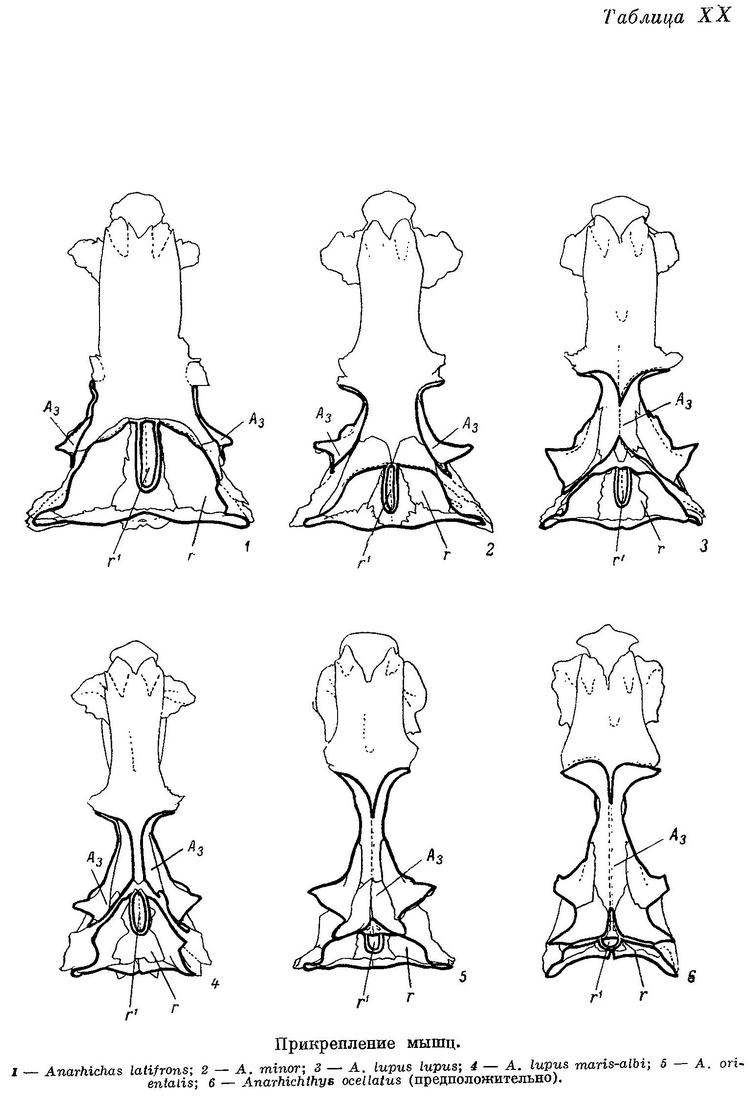
У молоди Anarhichas latifrons длиной 60—70 см, у сеголеток Anarhichas minor, Anarhichas lupus lupus, Anarhichas lupus maris-albi и Anarhichas orientalis основные порции m. adductor mandibulae сильнее наклонены назад: угол, который они образуют по отношению к нижней челюсти (вид сбоку), больше, чем у взрослых. Это вызвано гораздо большим, чем у взрослых, относительным размером глаз. Относительный размер мозга у молоди также больше, поэтому неврокраниум в местах прикрепления А3 сужен значительно меньше. У сеголеток Anarhichas lupus и Anarhichas orientalis киля в этой области, характерного для взрослых, еще нет, и верхние края порций А3 противоположных сторон еще далеко не соприкасаются. Ширина неврокраниума в этом месте значительно превышает его ширину между глазницами. В дальнейшем относительный размер глаз и мозга сильно уменьшается, угол основных порций m. adductor mandibulae приближается к прямому (вид сбоку), а сужение неврокраниума в местах прикрепления А3 увеличивается тем интенсивнее, чем специализированней данный вид зубаток. У Anarhichas lupus lupus и Anarhichas lupus maris-albi длиной 20—30 см заглазничная область неврокраниума уже суживается до узкого киля так, что верхние края А3 противоположных сторон соприкасаются. У Anarhichas minor такого сильного сужения не происходит, но все же ширина неврокраниума в этом месте становится у взрослых немного меньше, чем ширина его между глазницами. Наконец, у Anarhichas latifrons она в течение всей жизни остается заметно большей, чем ширина межглазничного промежутка, и лишь изредка равна ему.
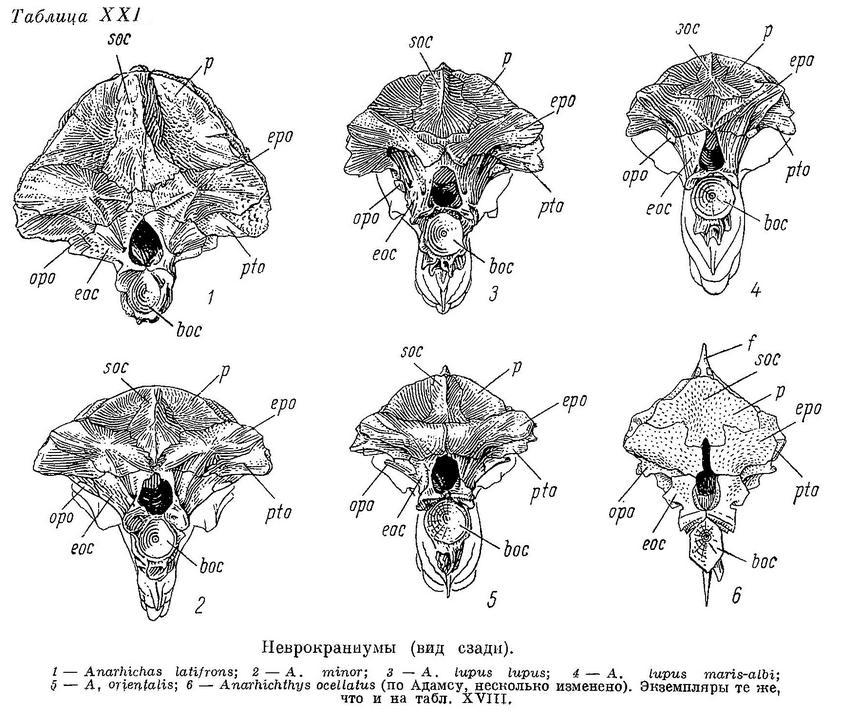
Следует отметить, что наибольшие давящие зубы нижней челюсти у молоди Anarhichas lupus lupus дальше сдвинуты назад, чем у взрослых (табл. II, 3а); то же возрастное различие наблюдается и у Anarhichas minor (однако у молоди это тупоконические — в большей степени рвущие, чем давящие зубы). Наклон зубов внутрь в обоих случаях заметно слабее, чем у взрослых: по-видимому, в основном из-за этого сила, разводящая нижние челюсти, у молоди невелика, на что указывает слабое сужение неврокраниума в области прикрепления А3. С возрастом увеличивается удельный вес тела, а с ним и относительная ширина головы снизу. Наибольшие давящие зубы нижней челюсти при этом смещаются вперед вследствие слишком сильного увеличения расстояния между ними. Чем больше приспособлен данный вид к питанию организмами с твердым панцирем, тем меньше это смещение. Оно вызывает снижение относительных размеров жертв, делает менее выгодным рычаг второго рода — нижнюю челюсть. Первое с избытком компенсируется увеличением наклона давящей поверхности зубов внутрь, второе — сильным развитием жевательных мышц, сопровождающимся, в силу увеличения этого наклона, сужением неврокраниума в области их прикрепления.
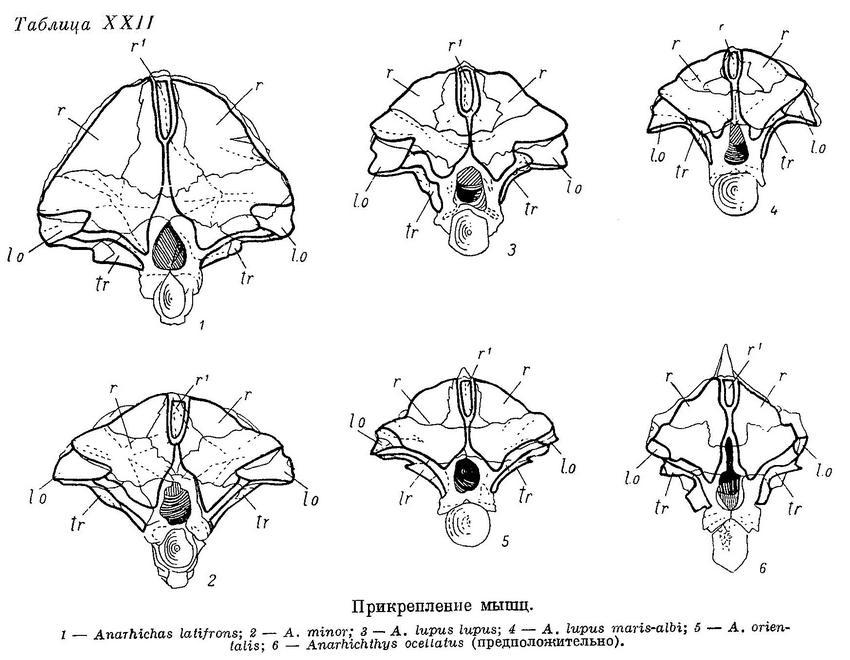
Изменения в строении жевательных мышц и связанных с ними костей, происходящие по мере роста зубаток, свидетельствуют о том, что в питании этих рыб увеличивается роль организмов с более твердым панцирем, притом увеличивается тем раньше и интенсивнее, чем больше эта роль в питании взрослых особей той или иной формы зубаток. Следовательно, изучение строения жевательных мышц и связанных с ними костей приводит к тем же выводам, что и изучение зубной системы. К сожалению, строение мышц и черепа у молоди Anarhichthys ocellatus размером менее 60 см совершенно неизвестно.
Как видно из изложенного, наиболее характерные особенности в строении черепа всего семейства в целом — суженная и низкая заглазничная часть черепа, отодвинутые от глазницы sphenotica с сильным гребнем на каждой из них, мощный киль на парасфеноиде, пережим нижнечелюстных костей, жесткое соединение suborbitalia — обусловлены особенностями в строении жевательных мышц и зубной системы, а в конечном счете — особенностями питания. Замечательно, что наиболее существенные различия между видами в строении черепа заключаются в степени развития указанных черт, наиболее характерных для всего семейства.
Мышцы, связанные с отведением нижней челюсти, работой глоточного и дыхательного аппаратов, достаточно подробно не исследовались. Нужно лишь отметить, что видовые отличия в строении этих аппаратов и принадлежащих им мышц меньше, нежели в строении челюстного аппарата.
В ряду Anarhichas latifrons — Anarhichas minor — Anarhichas lupus — Anarhichas orientalis, по-видимому, наблюдается при общем увеличении зажаберных полостей постепенное усиление жаберной перепонки по сравнению с жаберной крышкой. Жаберная крышка смещается вверх, мышцы, связанные с ее работой, укорачиваются, лучи жаберной перепонки и их мышцы укрепляются, ширина жаберной щели по отношению к длине головы снижается. Возможно, это связано с уменьшением подвижности зубаток в данном ряду.
Правый и левый m. levator operculi вместе с соединяющими эти мышцы поперечными пучками m. hyohyoideus представляют собой по существу единый сфинктер зажаберных полостей, прерванный в области неврокраниума и лишь частично в области каждой operculum. Антагонистами этого сфинктера являются m. dilatator operculi и косые пучки m. hyohyoideus (табл. XI—XII, XV—XVI).
Почти все мышцы у всех видов зубаток прикрепляются не только к одним и тем же костям, но и к одним и тем же их участкам. Случаи перемещения областей прикрепления мышц с одних участков кости на другие редки, перемещения незначительны. Изменениям в форме и расположении мышц сопутствуют соответствующие изменения в форме и расположении костей. Связь между характером развития отдельных мышц и строением тех костей, к которым они прикрепляются и на которых лежат, очевидна.
Кишечник
Пищеварительный тракт зубаток состоит из короткого и широкого пищевода, слабо обособленного от него желудка и четко отграниченной от желудка кишки.
Пищевод одет, в отличие от пищевода ряда других рыб, таким же эпителием, как и глотка, и имеет некоторые особенности в расположении мышечных волокон (известно для Anarhichas lupus lupus — Haus, 1897 : 43). Желудок сильно растяжим: вес его содержимого иногда составляет до 20% от веса всего тела, превышая в 14 раз вес самого желудка (Anarhichas minor, Anarhichas lupus maris-albi). При сильном раздражении (удар, порез) пища из желудка, может выбрасываться наружу. Между желудком и кишкой клапана нет. Кишка растяжима значительно меньше, чем желудок. Вес ее содержимого достигает лишь 8.3 (Anarhichas lupus maris-albi), 9.3% (Anarhichas minor) от веса тела. Вес пустых желудка и кишки примерно одинаков: в среднем 1.6 (желудок) к 1.8% (кишка) от веса тела. Кишка очень широкая; от желудка к анусу незначительно суживается, делая несколько изгибов. Длина кишки примерно равна длине зубатки, и превышает длину туловища (от конца жаберной крышки до начала анального плавника) у Anarhichas minor в среднем в 2.6 (17 экз.), у Anarhichas lupus lupus в 3.3 (1 экз.), у Anarhichas lupus maris-albi в 3.8 раза (104 экз.). Увеличение относительной длины кишки, вероятно, связано с увеличением роли моллюсков в питании, как известно, трудно усваиваемых (Карпевич, 1940 : 31).
Переваривание пищи происходит и в желудке, и в кишке. Рыбы, заглотанные зубаткой, перевариваются на протяжении всего кишечника: даже в задней части кишки позвоночники добычи еще не освобождаются полностью от мяса.
У зубаток, пойманных с сильно наполненным кишечником, задняя часть кишки часто бывает вывернута наружу. У одной из вскрытых мной Anarhichas lupus maris-albi средний участок кишки, наполненной обломками мидий и мелких брюхоногих, был вдвинут в соседний участок. Возможно, выворачивание кишки для освобождения ее от застрявших обломков — естественное явление у зубаток. Необычайно большая ширина ануса зубаток (Kämpfe u. Kittel, 1952—1953, Аbb, 5) несомненно связана со значительной величиной обломков раковин и панцирей пищевых организмов, проходящих через их кишечник, что отмечалось еще Пеннантом (Pennant, 1769 : 120).
Пилорических придатков нет, но на самом широком месте кишки, в ее начале, имеются две выпуклости, являющиеся их рудиментами (Haus, 1897 : 49; Макушок, 1958, рис. 6, Д).
Печень крупная, состоит из двух или, реже, трех долей; содержит 22—42% жира (по данным ПИНРО). Вес ее сильно колеблется в зависимости от сезона, состояния половых продуктов и т. п. Желчный пузырь сильно растяжим, желчный проток один, очень длинный, впадает в кишку сразу под указанными выше рудиментами пилорических придатков.
В поджелудочной железе молодой Anarhichas lupus lupus найден только один «основной» островок Лангерганса. Он расположен в области воротных сосудов печени, так же как у бельдюги и некоторых других рыб (Baron, 1935 : 34).
Объекты питания и способ их размельчения зубами накладывают заметные отпечатки на строение пищеварительного канала зубаток; особенно бросается в глаза значительная ширина кишки и ее выводного отверстия.
Дополнение к разделу «Кишечник». У пелагических мальков всех трех атлантических видов зубаток (длина 19—25 мм и выше), по данным Т. К. Сысоевой, желудок хорошо обособлен от кишки.
Форма тела. Плавники.
Атлантические зубатки близки между собой по форме тела, особенно Anarhichas lupus и Anarhichas minor; тихоокеанские своеобразнее, прежде всего Anarhichthys ocellatus, отличающаяся чрезвычайно удлиненным телом. В ряду Anarhichthys ocellatus — Anarhichas orientalis — Anarhichas lupus maris-albi — Anarhichas lupus lupus — Anarhichas minor — Anarhichas latifrons относительная высота тела увеличивается; возрастные изменения каждой из форм происходят в том же направлении (рис. 6). На рисунке не приведена наибольшая высота тела, так как она сильно изменяется в зависимости от степени наполнения кишечника, зрелости гонад и т. д. Судя по изгибу спины, наибольшая высота тела в указанном ряду видов и с возрастом у каждого из них не только в общем возрастает, но и перемещается от конца головы к середине туловища. В данном ряду видов ослабевают задние нижние части туловищной мышцы и усиливаются передние верхние; площадь прикрепления последних на черепе увеличивается (табл. XXII). Верх головы становится шире. Форма головы в поперечном сечении становится менее треугольной, более округлой, профиль ее, в задних частях головы, приобретает более крутые скаты, особенно верхний. Обводы тела становятся более плавными, обтекаемыми. Вес тела при той же длине возрастает (рис. 7), по-видимому в основном за счет увеличения общей массы мышц. Границы между спинным и анальным плавниками и телом становятся менее определенными, так как основания плавников утолщаются. Это должно улучшать обтекаемость оснований плавников.
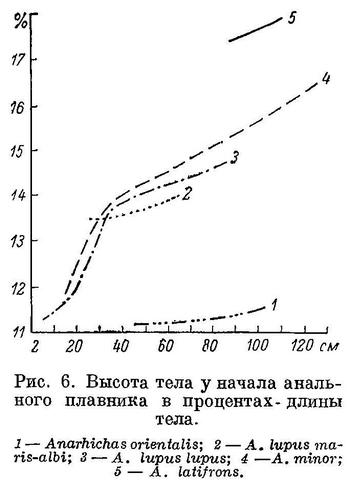
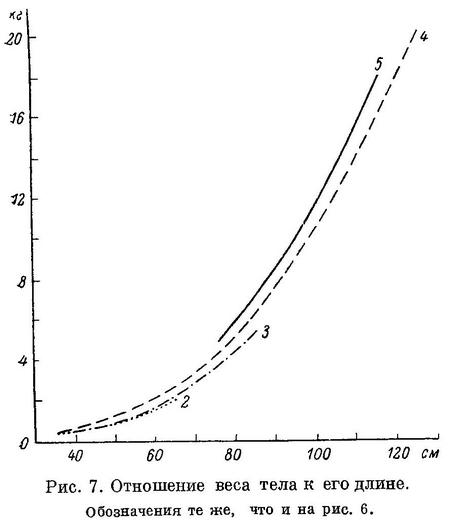
Таким образом, в приведенном выше ряду видов и с возрастом у каждого из них наблюдается тенденция к приближению формы тела к форме тела «обычных» рыб — торпедообразной, но несколько сжатой с боков. Несмотря на это, все зубатки отличаются очень большой и довольно слабо увеличивающейся к хвосту амплитудой волны, проходящей по телу при плавании. Иными словами, тип движения всех зубаток близок к угреобразному (Gray, 1933а, 1933b, 1933с; Шулейкин, 1934, 1953 : 838). Эта особенность их движения по сравнению с другими рыбами, довольно близкими по общей форме тела, бросается в глаза и издавна отмечалась многими авторами (Fabricius, 1780 : 139, и др.). Плавательная волна у пятнистой зубатки и трески наблюдалась мной в небольшом бассейне « морской водой на борту экспедиционного судна «Персей-2». Разница была весьма значительной несмотря на то, что по общей форме тела, исключая плавники, эти рыбы не так уж резко отличаются друг от друга.
Анализ плавательной волны, а именно анализ возникающих сил, препятствующих движению и поддерживающих его, а также сил, отклоняющих движение от прямой, доступен современной гидродинамике, очевидно, лишь в самых общих чертах. В настоящее время различаются два основных типа плавательной волны: 1) с большой и слабо увеличивающейся к хвосту амплитудой плавательной волны у рыб с угреобразной формой тела; 2) с малой и сильно увеличивающейся к хвосту амплитудой у рыб с «обычной» формой тела. Внутри каждого из этих типов существует масса вариаций. В общем для продвижения «обычных» рыб, при равных условиях, требуется меньшая сила благодаря их хорошей обтекаемости, чем для движения плохо обтекаемых угреобразных рыб, но зато больше непроизводительных затрат на дрейфовые движения и рыскание, сведенные у угреобразных рыб до минимума (Шулейкин, 1934, 1953). В связи с этим приведенный шаг, т. е. расстояние, проходимое рыбой за период одной волны, отнесенное к длине рыбы, как утверждает Шулейкин (1953 : 838) на основании кинематограмм Грея (Gray, 1933а, pls), больше всего у угря (0.57). Однако Бейнбридж (Bainbridge, 1958) обнаружил у форели, ельца и карася больший приведенный шаг, а именно 0.61—0.63. Возможно, при исследовании угря с помощью той же новейшей техники его приведенный шаг окажется еще большим. Достаточно тонкого гидродинамического анализа, с помощью которого можно было бы решить, какая форма тела отвечает большей скорости движения, пока что, очевидно, не имеется. Однако наблюдения над скоростью миграций рыб (Meek, 1916; Шмидт, 1947, и др.) позволяют с достаточной уверенностью полагать, что именно торпедообразная, хорошо обтекаемая форма тела лучше всего способствует быстрому движению при больших переходах, не говоря уже о коротких бросках: рыбы, способные к наиболее быстрым броскам, а именно марлины и мечь-рыба (Anonimous, 1947 : 153; Wisner, 1958 : 67, и др.), обладают торпедообразной формой тела. Поэтому приближение формы тела к торпедообразной, увеличение обтекаемости в ряду Anarhichthys ocellatus — Anarhichas orientalis — Anarhichas lupus maris-albi — Anarhichas lupus lupus — Anarhichas minor — Anarhichas latifrons, а также у каждой из форм с возрастом может свидетельствовать о возрастании средней скорости движения. Казалось бы, указанная скорость должна быть очень большой, так как в данном случае довольно хорошая обтекаемость тела, очевидно, сочетается с выгодным коэффициентом полезного действия. Однако «угреобразный» характер волны должен сам по себе заметно снижать выгоды хорошей обтекаемости тела. Это тем более вероятно, что частота плавательной волны у рыб находится в общем в обратной зависимости от амплитуды и наиболее быстрые пловцы характеризуются наименьшей амплитудой плавательной волны, причем значение хвостового плавника у них сильно возрастает (Nursall, 1958а : 119). Причины обратной связи между частотой и амплитудой волны, судя по всему, кроются не только во внешней среде, но и во внутренней, а именно в свойствах скелета и мышц. Штрассер (Strasser, 1882 : 86) справедливо указывал, что тело незначительной гибкости лучше приспособлено к быстрому движению, чем гибкое. Это положение ярко иллюстрируется таблицей, приведенной Нерсаллом (Nursall, 1958а : 119). Большая гибкость тела зубаток свидетельствует о том, что скорость движения их в общем невелика.
Выгоды лучшей обтекаемости могут использоваться зубатками полнее во время периодов «планирования» при отсутствии плавательных движений, если чередование активного движения с движением по инерции, вообще нередкое у костистых рыб (Harris, 1936 : 4.76), существует и у зубаток. Приближается ли характер плавательной волны у рода Anarhichas при больших скоростях движения к волне «обычных» рыб и в какой степени, вопрос остается открытым. Неизвестно также, как сильно изменяется амплитуда плавательной волны при сохранении примерно того нее самого соотношения между амплитудой головы и хвоста. В литературе до сих пор не имеется сведений ни о вариациях движений зубаток, ни о их скорости, исключая некоторые предположения, касающиеся полосатой зубатки: одни авторы (Bloch, 1785 : 24) считают ее медленным пловцом, другие (Yarrell, 1841 : 279) — быстрым. Ближе всего к истине, возможно, Ласепед, считающий, что зубатки плавают не очень быстро, но способны на короткие, довольно быстрые рывки (Lacepède, 1801 : 40).
Видовые и возрастные различия в характере плавательной волны у зубаток, за отсутствием достаточных прямых наблюдений, следует попытаться представить себе косвенным путем. Логично предположить, что с возрастанием амплитуды плавательной волны к хвосту число позвонков по направлению к последнему увеличивается быстрее. У зубаток и некоторых других рыб мной было подсчитано число позвонков в каждой десятой доле длины позвоночника, начиная от головы. Эти цифры, выраженные в процентах к общему числу позвонков, представлены в виде кривых (рис. 8, А и Б). Из рис. 8, А I, видно, что рыбы с «торпедообразной» формой тела — треска и сельдь — действительно отличаются от угря более резким возрастанием числа позвонков к концу хвоста. Казалось бы, у зубаток степень возрастания этого числа должна увеличиваться вместе с возрастанием относительной высоты тела. В действительности же увеличение числа позвонков к концу хвоста замедляется от Anarhichthys ocellatus к Anarhichas orientalis и далее к Anarhichas lupus, и лишь к концу ряда (Anarhichas minor — Anarhichas latifrons) начинает ускоряться (рис. 8, А, II). Почему же начало ряда не удовлетворяет нашим ожиданиям?

Аквариальные наблюдения над движением угря (Lowndes, 1955 : 250) свидетельствуют о том, что плавательная волна у угря не латеральная, а спиральная. Очевидно, это следует понимать таким образом, что тело угря изгибается не по кривой, лежащей в горизонтальной плоскости, а по спирали, так что изгибы определяются не двумя, а тремя измерениями. Следовательно, можно говорить о горизонтальной и вертикальной (латеральной и дорзовентральной) амплитудах плавательной волны. (Т. е. об амплитудах волны на горизонтальной и вертикальной проекциях.) Надо полагать, спиральный тип волны у угря не всегда бывает резко выражен, а когда хорошо выражен, то ограничивается в основном концом хвостового отдела, иначе он не ускользнул бы от внимания других исследователей, в том числе и Грея. Если это так, то вертикальная амплитуда волны угря должна резко увеличиваться к концу хвоста, что и вызывает более значительное увеличение числа позвонков к концу хвоста по сравнению с зубатками. Ближе всего к угрю стоит «позвоночная кривая» Anarhichthys ocellatus, и на основании большого сходства формы тела этого вида с угрем у Anarhichthys ocellatus также можно предполагать способность к спиральной плавательной волне. Правда, форма тела Anarhichthys ocellatus несколько уклоняется от угреобразной в сторону лентообразной, особенно, по-видимому, у молоди, ведущей, надо полагать, пелагический образ жизни. Поэтому дорзовентральная гибкость тела у Anarhichthys ocellatus должна быть несколько снижена по сравнению с угрем (Schlesinger, 1911 : 320), вертикальная амплитуда спиральной волны также, в соответствии с чем число позвонков к концу хвоста увеличивается немного медленнее, чем у угря. Anarhichthys ocellatus отличается чрезвычайно большим числом позвонков, латеральная гибкость тела при этом должна быть весьма значительной, а скорость движения в общем меньшей, чем у угря. У рода Anarhichas вертикальная амплитуда спиральной волны с увеличением высоты тела постепенно уменьшается, что и вызывает замедление увеличения числа позвонков к концу хвоста вплоть до Anarhichas lupus (точнее до Anarhichas lupus maris-albi; «позвоночные кривые» подвидов различаются незначительно и для упрощения рисунка объединены в одну кривую). В дальнейшем у Anarhichas minor и особенно у Anarhichas latifrons влияние уменьшения вертикальной амплитуды перевешивается влиянием увеличения горизонтальной амплитуды, что вновь ускоряет увеличение числа позвонков к концу хвоста, вызывает новое приближение «позвоночной кривой» к кривой Anarhichthys ocellatus, но уже на другой основе.

Это предположение как будто подтверждается особенностями строения хвостового плавника у зубаток. С увеличением высоты тела зубаток форма хвостового плавника закономерно изменяется от заостренной у Anarhichthys ocellatus к округлой у Anarhichas orientalis; далее в ряду Anarhichas lupus maris-albi — Anarhichas lupus lupus — Anarhichas minor — Anarhichas latifrons хвостовой плавник все более приближается по форме к усеченному (рис. 9). Таким образом, форма хвостового плавника имеет тенденцию приближаться к выемчатой, хотя и не достигает ее; выемчатая же форма характерна для более быстрых пловцов (Суворов, 1948 ; 33; Nursall, 1958а : 119, и др.). Однако в рассматриваемом частном вопросе для нас интереснее тот намек, который доставляют опыты над моделями гомоцеркального хвоста округлой и полулунной формы: оказывается, при волнообразных движениях хвоста округлый хвостовой плавник предоставляет заметно большую свободу для вертикальных движений, чем полулунный (Grove a. Newell, 1936 : 287, fig. 2Е, 2F). Если при этом учесть, что спиральная плавательная волна до сих пор отмечена лишь у рыб с заостренным либо гетероцеркальным хвостом (Lowndes, 1955 : 247 , 250), то законно предположить, что изменение формы гомоцеркального хвостового плавника от заостренной к усеченной связано с уменьшением вертикальной амплитуды спиральной плавательной волны. С укорочением хвоста (по сравнению с туловищем) в ряду Anarhichthys ocellatus — Anarhichas orientalis — Anarhichas lupus относительные размеры хвостового плавника увеличиваются. У Anarhichas minor и особенно Anarhichas latifrons они вновь уменьшаются, главным образом в связи с уменьшением относительной высоты хвостового стебля при основании хвостового плавника (рис. 10). Мощность мышц в конце хвостового отдела у Anarhichas minor и особенно у Anarhichas latifrons снижается по сравнению с Anarhichas lupus lupus (рис. 11), действие конца хвоста ставится в большую зависимость от его упругости и в меньшую — от силы мышц. Изменение относительных размеров хвостового плавника происходит сходно с изменением «позвоночной кривой» и, возможно, сходным образом связано с изменениями в характере плавательной волны.
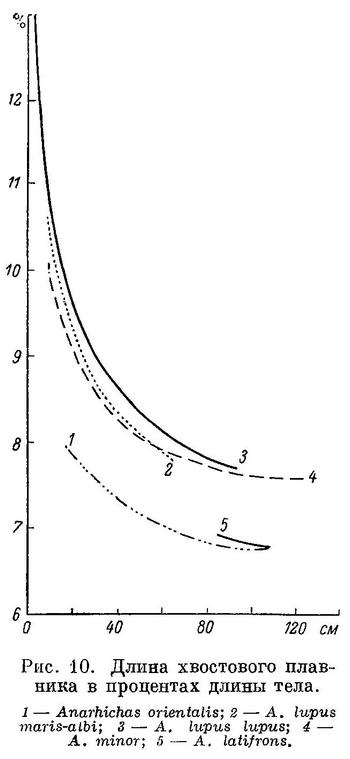
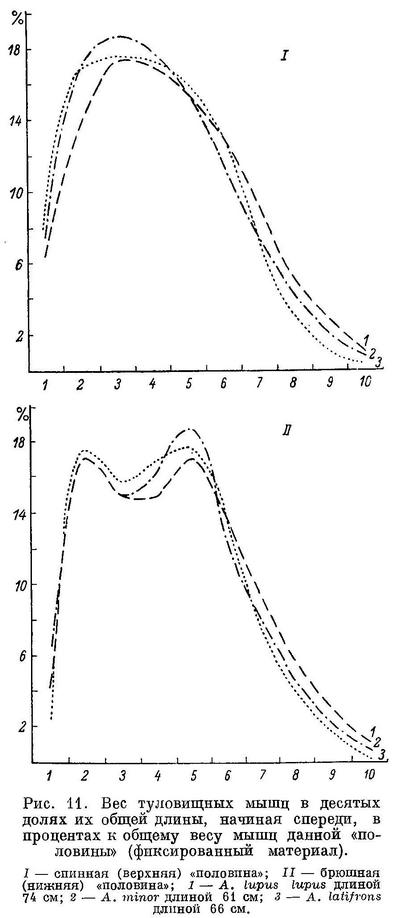
С возрастом у всех зубаток рода Anarhichas (по Anarhichthys ocellatus сведений нет) относительные размеры хвостового плавника уменьшаются, тогда как форма его почти не изменяется. Следовательно, уменьшение относительных размеров плавника с возрастом, как и увеличение степени возрастания числа позвонков к хвосту (рис. 8, Б),: может быть связано исключительно с увеличением горизонтальной амплитуды плавательной волны. Увеличение горизонтальной амплитуды, очевидно, может происходить при различной степени уменьшения вертикальной амплитуды. Характер «позвоночной кривой», возможно, не претерпевает значительных изменений в том случае, когда увеличение горизонтальной амплитуды уравновешивается уменьшением вертикальной амплитуды. В связи с этим интересно гораздо большее сходство «позвоночных кривых» трески и угря, чем трески и сельди, хотя по характеру плавательной волны треска должна быть гораздо ближе к сельди, чем к угрю.
Более подробное изучение формы позвонков и подвижности их относительно друг друга, естественно, даст гораздо более точное представление о характере плавательной волны, чем «позвоночные кривые». В частности, надлежит выяснить, насколько свойственны зубаткам другие особенности плавательной волны, отмеченные у рыб с угреобразной формой тела. Известно, например, что угорь может производить плавательные движения, не трогаясь с места; более того, он способен менять направление плавательной волны и двигаться хвостом вперед (Gray, 1933а, pl. IV; Lowndes, 1955 : 250), притом со значительной скоростью (Шулейкин, 1953 : 838). Способность производить плавательные движения, не трогаясь с места, известна и у зубаток (Anarhichas lupus lupus — Fabricius, 1780 : 139), но способны ли они плыть хвостом вперед, изменяя направление плавательной волны, неизвестно. Скорее всего, такой способностью они не обладают. Выяснение этого вопроса очень важно для правильного понимания особенностей работы плавников.
Осевой скелет зубаток исследован мной гораздо менее подробно, чем череп, что сильно затрудняет анализ связи его строения с характером движения. Неясно, например, значение ассиметрии позвонков, свойственной всем зубаткам. Правда, передняя половина тела позвонка, до перетяжки, в общем примерно равна задней, но боковая поверхность первой вогнутая, а последней выпуклая. Общее число позвонков уменьшается в ряду Anarhichthys ocellatus — Anarhichas orientalis — Anarhichas lupus maris-albi и возрастает в ряду Anarhichas lupus lupus — Anarhichas minor — Anarhichas latifrons (табл. 7), т. е. изменяется сходно с изменением «позвоночной кривой». Правда, увеличение общего числа позвонков по сравнению с Anarhichas lupus происходит у Anarhichas minor и Anarhichas latifrons гораздо медленнее, чем изменение «позвоночной кривой», в отличие от Anarhichas orientalis и Anarhichthys ocellatus. Интересно, что срастание позвонков наблюдается у зубаток только в хвостовом отделе. Чаще всего срастаются лишь тела позвонков, но иногда и основания остистых отростков. Частота встречаемости особей со сросшимися позвонками велика: 2 из 5 у Anarhichas orientalis, 16 из 33 у Anarhichas lupus maris-albi, 17 из 43 у Anarhichas lupus lupus, 12 из 33 у Anarhichas minor; 3 из 58 у Anarhichas latifrons.
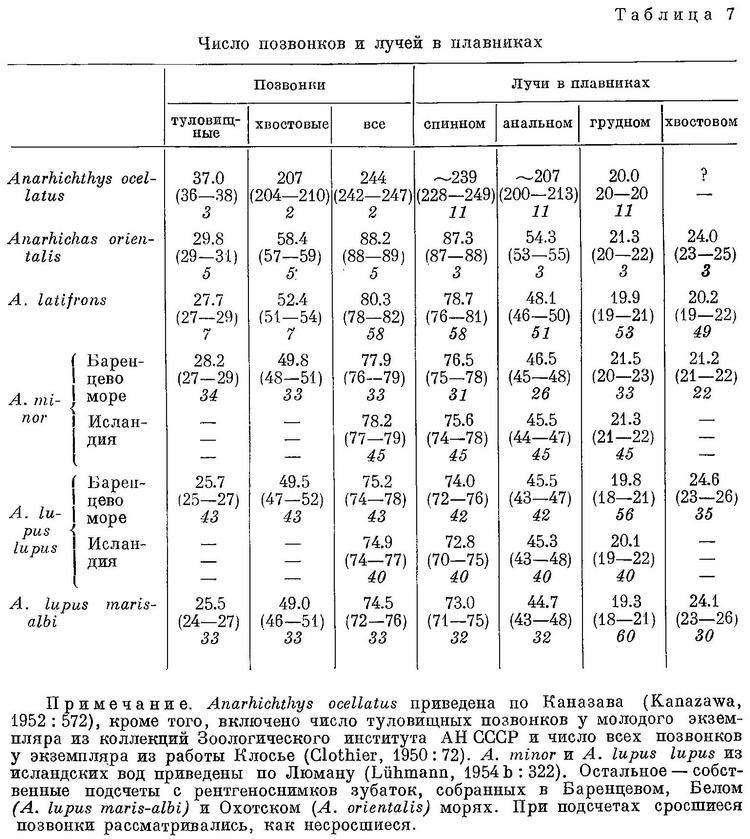
Число лучей в снинном и анальном плавниках непосредственно связано с числом позвонков и изменяется сходно с ним. Число лучей в грудном и хвостовом плавниках изменяется иначе: возможно, при формировании лучей их число на единицу площади плавника возрастает в ряду Anarhichthys ocellatus — Anarhichas orientalis — Anarhichas lupus — Anarhichas minor — Anarhichas latifrons. Это предположение не может быть пока подтверждено никакими конкретными данными.
Спинной и анальный плавники у зубаток простираются — спинной от затылка, анальный от ануса — почти до самого хвостового плавника. У Anarhichthys ocellatus они сливаются с хвостовым плавником, отделяясь от него лишь небольшими выемками (Clemens a. Wilby, 1946 : 171). У зубаток рода Anarhichas происходит гораздо большее обособление непарных плавников, однако они остаются связанными низкими перепонками. Перепонка, связывающая спинной плавник с хвостовым, поддерживается укороченными жесткими лучами спинного плавника, последний из которых находится в непосредственной близости с первым лучом хвостового плавника, что давно отмечалось (Parnell, 1838 : 239, и др.). Высота перепонки относительно высоты хвостового стебля с возрастом уменьшается, но все же у самых старых особей любой из форм перепонка не сливается с хвостовым стеблем. Перепонка между анальным и хвостовым плавниками лучами не поддерживается; у старых особей она утолщается и сливается с хвостовым стеблем.
Спинной и анальный плавники зубаток, очевидно, участвуют в плавательной волне, увеличивая площадь отталкивания от воды, а также вызывая «срыв тока» воды на своих вершинах, порождающий пропульсаторную силу (Gray, 1949 : 1704). У Anarhichas minor, а возможной у всех других зубаток, участие этих плавников в плавательной волне возрастает по направлению к хвосту, так как при прямолинейном движении приподнятость их лучей увеличивается вплоть до задних концов плавников. Впрочем, степень участия спинного и анального плавников в плавательной волне может зависеть от соотношения между скоростью движения рыбы в воде и скоростью плавательной волны; обе скорости могут изменяться даже при равномерном, на первый взгляд, движении (Nursall, 1958b).
Задние концы спинного и анального плавников, кроме пропульсаторной, должны постоянно порождать и подъемную силу, так как зубатки лишены плавательного пузыря и их удельный вес постоянно превышает удельный вес воды. Судя по всему, удельный вес зубаток в ряду Anarhichthys ocellatus — Anarhichas orientalis — Anarhichas lupus — Anarhichas minor — Anarhichas latifrons уменьшается, поэтому наиболее значительная подъемная сила должна вырабатываться хвостом взрослых особей Anarhichthys ocellatus. По-видимому, эта сила у них создается исключительно за счет вертикальной асимметрии плавательной волны. У зубаток же рода Anarhichas вертикальная амплитуда волны для этого, возможно, обычно недостаточна: высота задних концов спинного и анального плавников увеличивается и выработка подъемной силы ложится в основном на них. Анальный плавник, увеличивая пропульсаторную силу, приложенную к нижнему краю хвоста, если следовать Васнецову (1948 : 15), должен создавать при плавательной волне поворот вверх, спинной, напротив, поворот вниз. Поскольку высота спинного плавника значительно превышает высоту анального, действие первого должно преобладать. Однако основным является здесь другое: гибкие концы лучей анального плавника отгибаются под напором воды вверх и таким образом создают силу, поднимающую хвост (эта сила значительно превышает опускающую силу, вырабатываемую тем же плавником в результате его расположения по нижнему краю хвоста); сходное явление отмечено у сомовых (Harris, 1938 : 36). Если бы лучи спинного плавника оставались гибкими, то их концы при плавательной волне создавали бы значительную силу, опускающую хвост. Однако гибкость указанных лучей у зубаток из рода Anarhichas заметно уменьшается по направлению к хвосту, лучи утолщаются. Возможно, при плавательной волне общая поверхность задней части спинного плавника движется под небольшим углом к вертикали, навстречу воде, как это отмечено у акул (Le Маге, 1936 : 431, fig. 2), и таким образом дополнительно создает подъемную силу. Перед самым хвостовым плавником этот угол, очевидно, сильно уменьшается, и хвостовой плавник располагается вертикально, в связи с чем спинной плавник на заднем конце скошен, лучи его сильно укорочены. Разрыв между анальным и хвостовым плавниками также обеспечивает вертикальное положение хвостового плавника.
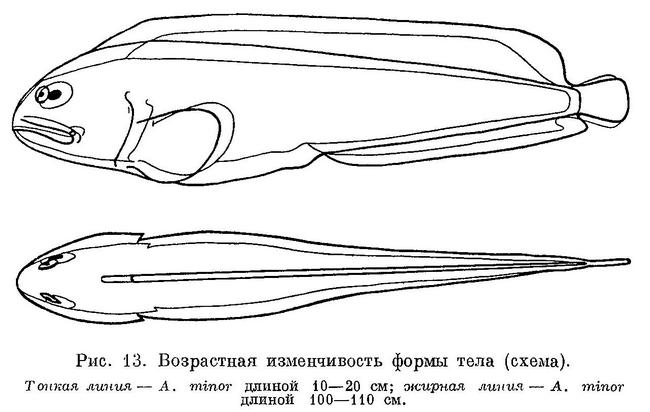
Высота задних концов спинного и анального плавников относительно высоты хвоста в этой области у Anarhichas lupus и Anarhichas minor увеличивается с возрастом (рис. 13) соответственно увеличению удельного веса и уменьшается с уменьшением удельного веса в ряду одноразмерных особей Anarhichas orientalis — Anarhichas lupus — Anarhichas minor. Однако у Anarhichas latifrons, отличающейся самым низким удельным весом, высота задних концов спинного и анального плавников не уменьшается далее, а вновь увеличивается, по-видимому, в силу того, что различия в высоте задних концов спинного и анального плавников становятся не столь резкими, как у других Anarhichas, и возможно, задняя часть спинного плавника образует меньший угол наклона навстречу воде. У Anarhichas latifrons задний конец анального плавника особенно сильно выдается назад в виде лопасти. Возможно, эта лопасть снижает вихреобразование при движении, как и хвостовой плавник; в таком случае понятно ее наибольшее развитие у Anarhichas latifrons, обладающей наиболее усеченным хвостовым плавником, т. е. наиболее близким по форме к выемчатому, заостренные концы которого почти несомненно способствуют снижению вихреобразования (Nursall, 1958а : 117). Вряд ли эту лопасть можно считать рулевой, так как лучи ее очень гибкие и снабжены наиболее слабыми мышцами. Мышцы, прикрепляющиеся к лучам спинного и анального плавников, весьма заметно ослабевают по направлению к задним лучам. Мышцы хвостового плавника у зубаток также развиты слабо и к тому же упрощены (Grenholm, 1923 : 190). Грей (Gray, 1933d : 121), изучавший повороты рыб в горизонтальной плоскости с помощью кинематограмм, не получил никаких доказательств использования хвостового плавника в качестве руля у рыб, двигающихся с помощью туловищных мышц. У рыб со сравнительно коротким и негибким телом хвостовой плавник при поворотах служит в основном точкой опоры, но не рулем. У зубаток роль хвостового плавника при поворотах должна быть небольшой, поскольку латеральная гибкость тела у них чрезвычайно велика (у только что пойманной Anarhichas lupus maris-albi хвост можно легко пригнуть к бокам головы). При прямолинейном движении задние концы спинного и анального плавников у Anarhichas несомненно до известной степени замещают хвостовой плавник в его функции, создавая пропульсаторную силу. Взаимодействие между этими плавниками и плавательной волной должно быть сложным и в этом отношении многое предстоит уточнить и разрешить.
Сила, поднимающая хвост, заставляла бы рыбу совершать поворот вниз, если бы к переднему концу тела не была также приложена подъемная сила. Такая сила должна вырабатываться у зубаток несколько уплощенной нижней поверхностью тела, прежде всего нижней поверхностью головы, широкой, плоской и расположенной под острым углом к направлению движения. Сверху голова и тело заметно уже, чем снизу. Сходная особенность в форме тела, связанная с высоким удельным весом, отмечена у акул; она увеличивает подъемную силу (Harris, 1936), но явно уменьшает обтекаемость, а с нею и скорость движения (Breder, 1930 : fig. 1, D). Возрастные и видовые различия в соотношении между шириной спинной и брюшной стороны тела происходят у зубаток в основном в соответствии с изменением удельного веса, и Anarhichas latifrons в данном отношении ничем особенным не отличается: это различие у нее наименьшее в связи с наименьшим удельным весом. Регулировка случайных вертикальных отклонений от прямой производится у акул с помощью грудных плавников. Возможно, и у зубаток это выполняется грудными плавниками, путем отведения верхних либо нижних лучей вперед или с помощью волны, пробегающей по плавникам сверху вниз либо снизу вверх. У донных костистых рыб с высоким удельным весом эти плавники могут быть даже постоянным источником подъемной силы (Шмальгаузен, 1916 : 191).
Повороты в горизонтальной плоскости у рыб, в том числе и у угря, обычно очень успешно совершаются с помощью плавательной волны с необычно большой амплитудой и малой скоростью; мышечные сокращения проходят при этом по тому боку, в направлении которого производится поворот (Gray, 1933d). Угловая скорость поворота может регулироваться и зависит от степени асимметрии плавательной волны и от частоты ее (Gray, 1937 : 97). Весьма вероятно, что и у зубаток повороты в горизонтальной плоскости совершаются тем же путем. При этом не только хвостовой плавник, но и задние части спинного и анального плавников могут служить точкой опоры, а также активно помогать повороту при прохождении через них плавательной волны. Возможно, в первой фазе поворота, когда поворачивается голова, спинной плавник спереди остается сложенным; в последней же фазе (когда поворачивается хвост) расправляется, если поворот должен быть ограничен. Повороту могут способствовать удары грудного плавника; на кинематограммах Грея (Gray, 1933d, pl. 7) это ясно заметно у мерланга.
Не исключена возможность, что повороты в вертикальной плоскости у зубаток в какой-то степени могут осуществляться за счет вертикальной асимметрии спиральной плавательной волны и что спиральный характер волны проявляется заметнее как раз при таких поворотах. При более резких и крутых поворотах, чем позволяет эта волна, а также при поворотах в ее отсутствие могут включаться передние части спинного и анального плавников, а также грудные плавники.
Спинной и анальный плавники могут вызывать повороты рыбы в вертикальной плоскости либо путем одновременного действия всеми лучами в одну сторону, либо путем поочередного движения лучей (Васнецов, 1948 : 15), либо путем сочетания этих двух способов (Шмальгаузен, 1913 : 214—219; Harris, 1937 : 185). Последнее, очевидно, наиболее вероятно при длинных плавниках. Одновременное действие всеми лучами основано исключительно на упругой волне, проходящей вдоль луча, от основания к вершине; поочередное движение лучами порождает волну, идущую вдоль плавника. Соотношение между двумя этими системами волн очень сложно. Скорость упругой волны зависит от гибкости лучей, а также, судя по всему, и от толщины покровов, облегающих и соединяющих лучи; направление — от характера изогнутости лучей. Гибкость и изогнутость лучей изменяются у зубаток на протяжении одного и того же плавника, а также с возрастом, видом и т. д.; толщина покровов с возрастом сильно увеличивается. Скорость и направление продольной волны регулируется скоростью и характером сокращений мышц плавников.
Возможно, действие спинного и анального плавников при поворотах в вертикальной плоскости довольно разнообразно. Как бы то ни было, прежде всего в таких поворотах участвуют передние части этих плавников, о чем свидетельствует возрастание мощности мышц плавников от задних лучей к передним. При поворотах на месте действие плавников, возможно, усиливается изгибанием тела в вертикальной плоскости. Дорзовентральная гибкость тела зубаток очень значительна, особенно дорзальная: у изогнутых вверх до отказа Anarhichas lupus maris-albi длиной 20—30 см расстояние от конца рыла до середины хвостового плавника составляет около 56 (38—67)%, у изогнутых вниз — около 64 (48—78)% длины тела, у особей длиной 50—65 см — 59 (40—79) и 69 (40—90)%. Замена дорзовентрального поворота латеральным при поворотах в вертикальной плоскости, характерная для рыб с ничтожной дорзовентральной гибкостью тела (Алеев, 1958а : 463), у зубаток вряд ли наблюдается.
При горизонтальных поворотах в отсутствие плавательной волны {одновременное сокращение мышц на стороне поворота), если таковые вообще существуют, спинной и анальный плавники расправляются, очевидно, таким образом, что приближают центр реакции к центру тяжести рыбы (Алеев, 1957).
Сочетание поворотов в горизонтальной плоскости с поворотами в вертикальной, надо полагать, также производится и с помощью асимметрии плавательной волны (включаются и горизонтальная и вертикальная амплитуды), и с помощью работы плавников. В данном случае роль грудных плавников, возможно, больше, чем роль непарных.
Грудные плавники у зубаток развиты сильно, широкие, округлые, посажены низко. Их основание расположено почти вертикально, вогнуто спереди. Наиболее длинные лучи находятся в верхней половине плавника. С помощью грудных плавников зубатки, очевидно, осуществляют движение вперед и назад при отсутствии плавательной волны, а также разнообразные повороты при отсутствии и наличии ее. Движения назад, очевидно, затруднены, так как m. adductor radialis дифференцирован слабо (Grenholm, 1923 : 190; Harris, 1937 : 186). В частности, отведение нижней части грудного плавника способствует быстрому и резкому повороту вниз. У молодых особей Anarhichas lupus lupus оно часто наблюдается вместе с плавательными движениями, даже на воздухе, если зубатку задеть. Расположение нижнего края плавника вблизи нижнего края тела облегчает такой поворот. Функция грудных плавников у костистых рыб вообще очень разнообразна, особенно у рыб с примерно одинаковым развитием отводящих и приводящих мышц грудных плавников; специализация в каком-либо направлении приводит к сдвигу весового соотношения этих мышц в ту или иную сторону (Harris, 1937 : 186). У зубаток отводящие и приводящие мышцы грудных плавников развиты примерно одинаково. В ряду Anarhichas lupus lupus — Anarhichas minor — Anarhichas latifrons вес отводящих мышц несколько увеличивается, составляя соответственно 95, 105 и 113% от веса мышц приводящих (исследовано по одной особи каждого вида длиной 74, 61 и 67 см). Возможно, роль грудных плавников при поворотах на быстром (относительно) движении увеличивается, что может быть связано с видовыми различиями в характере плавательной волны. Относительные размеры грудных плавников в указанном ряду уменьшаются. Правда, при одинаковой длине тела грудные плавники Anarhichas minor даже несколько превышают по длине плавники Anarhichas lupus, но уступают им по площади ввиду меньшей ширины плавника. У Anarhichas latifrons и длина грудных плавников, не говоря уже о площади, заметно меньше, чем у Anarhichas minor. Снижение относительных размеров этих плавников, наряду с приведенными выше соображениями, также может свидетельствовать о возрастании средней скорости движения в указанном ряду видов (Алеев, 19586), при условии, что зубатки редко употребляют грудные плавники в качестве тормозов.
Брюшные плавники у зубаток отсутствуют, но тазовые кости имеются. Задние концы их соприкасаются. Передние концы, далеко отодвинутые друг от друга, прикрепляются связками к внутренней стороне нижних концов cleithra. При переднем положении брюшных плавников их роль в поворотах в вертикальной плоскости больше, чем при заднем положении (Васнецов, 1941 : 505); кроме того, брюшные плавники совершенствуют торможение, производимое с помощью грудных плавников, устраняя поворот либо подъем вверх, сопровождающий это торможение (Harris, 1938 : 37—42, fig. 3—6). Возможно, у зубаток широкие грудные плавники совмещают функцию и грудных и брюшных. Кроме того, повороты в вертикальной плоскости у зубаток облегчаются большой дорзовентральной гибкостью тела и мощными непарными плавниками — спинным и анальным.
Остановимся теперь подробнее на различиях в удельном весе зубаток, оказывающем, как было видно из вышеизложенного, заметное влияние на их строение через характер движения. Правда, точных определений удельного веса зубаток, насколько мне известно, нет. Однако достаточно надежные косвенные данные свидетельствуют о том, что в ряду Anarhichas orientalis — Anarhichas lupus — Anarhichas minor — Anarhichas latifrons удельный вес уменьшается. В этом ряду возрастает, особенно в конце ряда, пористость костной ткани и частичное замещение ее хрящом. Содержание воды в мышцах увеличивается: у Anarhichas lupus lupus и Anarhichas minor оно достигает 75—82%, у Anarhichas latifrons — 90—92% (Травин, 1952 : 175). Мышцы трески содержат 80% воды (Lowndes, 1943 : 36); понятно, что у Anarhichas lupus и Anarhichas minor, не снабженных плавательным пузырем, удельный вес должен быть заметно большим, чем у трески. Anarhichas lupus lupus и Anarhichas minor в данном случае не различаются, по-видимому, лишь ввиду очень небольшого числа наблюдений. Величина сокращения размеров зубаток и, в частности, длины тела (табл. 8) при длительном хранении в спирте, отнимающем воду, свидетельствует о том, что содержание воды в тканях у Anarhichas lupus lupus и Anarhichas minor уменьшается с возрастом, особенно сильно у Anarhichas minor, и увеличивается в ряду одноразмерных особей Anarhichas lupus lupus — Anarhichas minor — Anarhichas latifrons, особенно у Anarhichas latifrons. Насыщение тканей водой, облегчение костной ткани и частичное замещение ее хрящом явно снижает удельный вес зубаток. Выброшенная за борт Anarhichas latifrons долго не тонет (Kampfe u. Kittel, 1952 : 240; собственные наблюдения), чего нельзя сказать об Anarhichas minor и особенно об Anarhichas lupus. Сокращение тела зубаток при хранении в спирте, пропорциональное содержанию воды в их тканях и обратно пропорциональное удельному весу, возрастает соответственно увеличению глубин, предпочитаемых данными видами и возрастами. Уменьшение удельного веса при уходе на глубины, по-видимому, связано с необходимостью чаще подниматься в толщу воды над дном в поисках пищи (ввиду снижения плотности поселения основных объектов питания), а возможно, и в поисках более благоприятного кислородного режима (грунты в местах обитания изменяются от каменистых к илистым. Anarhichthys ocellatus, по-видимому, обитает на наименьших глубинах по сравнению с другими зубатками, так как только этот вид отмечен в лужах, остающихся после отлива). У взрослых особей угревидной зубатки можно ожидать наибольший удельный вес, тогда как у молоди данного вида, ведущей, очевидно, пелагический образ жизни, удельный вес может оказаться наименьшим.

Различная прочность скелета, различное содержание воды в мышцах зубаток должны вызвать какие-то отличия в силе, выносливости и скорости сокращения мышц, а следовательно, и в характере движения. Этот вопрос, очень важный для правильного суждения о характере движения зубаток, остается, однако, совершенно нерешенным. Можно полагать, что уменьшение удельного веса, достигаемое таким путем, затрудняет переход к плавательной волне с небольшой амплитудой и большой частотой, обычной для быстро плавающих рыб и требующей, в частности, прочного скелета (Nursall, 1958а). Тем не менее даже и такое уменьшение удельного веса дает возможность перехода к более обтекаемой форме тела (см. выше). Отсюда следует, что уменьшение удельного веса, достигаемое путем насыщения тканей водою, с одной стороны, способствует увеличению скорости движения, а с другой стороны, мешает, и, возможно, довольно скоро ставит предел этому увеличению. У акул, по крайней мере у обычных видов из сем. Сarcharhinidae, которых нам случалось ловить в Индийском океане, гибкость тела весьма значительно уступает таковой у зубаток. При этом сохранение высокого удельного веса, по-видимому, неизбежно. Чтобы не тонуть, акула должна постоянно находиться в движении (Harris, 1936 : 486, и др.); с таким неудобством приходится мириться, очевидно, ради сохранения достаточно большой скорости движения. Возможно, увеличение скорости движения акул лимитируется главным образом их высоким удельным весом: как бы обтекаемость тела акул ни приближалась к идеалу, она остается довольно далекой от него ввиду необходимости выработки при движении подъемной силы. Сочетание наибольшей обтекаемости с наименьшей гибкостью тела, позволяющее перейти и этот предел скорости, в таком случае может быть достигнуто только рыбами, обладающими плавательным пузырем. Последний, однако, имеет свои отрицательные стороны — в частности, затрудняет быстрые передвижения по вертикали.
Таким образом, при типе строения, характерном для зубаток, приспособления к увеличению обтекаемости тела связаны с уменьшением удельного веса и притом противоречат приспособлениям к питанию организмами с прочным панцирем: особенности строения челюстей и жевательных мышц требуют тем большего сужения головы сверху одновременно с расширением ее снизу и уменьшением высоты за глазами, в области прикрепления внутренней порции жевательных мышц, чем сильнее приспособлен зубной аппарат к питанию организмами с прочным панцирем, увеличение же обтекаемости тела вызывает сглаживание разницы между шириной головы сверху и снизу, а также увеличение высоты головы за глазами. В соответствии с этим ряд видов, в котором увеличивается приспособленность к более быстрому движению, противоположен ряду с увеличивающейся специализацией к питанию организмами с твердым панцирем. Следует заметить, что у видов семейства Sparidae нижнечелюстным костям противостоит не непарный сошник, принимающий на себя основную долю нагрузки, как у зубаток, а обе межчелюстные кости, причем зубы на каждой нижнечелюстной кости приходятся непосредственно под зубами соответствующей межчелюстной кости. При подобном расположении зубов сила, раздвигающая челюсти при раздавливании моллюска, должна быть невелика, поэтому прикрепление жевательных мышц к неврокраниуму не является необходимым. Они прикрепляются, очевидно, лишь к нёбной дуге (Gregory, 1933 : 248—250, figs 122—124). При таком типе строения противоречия между увеличением обтекаемости тела и приспособлением к питанию организмами с прочным панцирем, по-видимому, не существует.
Тело и плавники рыбы в действии — весьма сложная система. Взаимодействие ее частей непрерывно изменяется в зависимости от скорости движения, его направления относительно основных осей тела рыбы и относительно центра Земли, его траектории, в зависимости от плотности и вязкости воды и т. д. Нет никакого сомнения в том, что высказанные выше соображения относительно характера движения и его влияния на форму тела у зубаток являются лишь самыми предварительными наметками для дальнейших исследований, в ходе которых многие из этих представлений могут в корне измениться. Имеется, однако, некоторая надежда, что основной вывод о возрастании средней скорости движения, в общем небольшой, в ряду одноразмерных особей Anarhichthys ocellatus — Anarhichas orientalis — Anarhichas lupus maris-albi — Anarhichas lupus lupus — Anarhichas minor — Anarhichas latifrons, а также у каждой из этих форм с возрастом, останется в силе. Дело в том, что изучение распределения и распространения атлантических зубаток, пусть тоже предположительно, но независимо, на основании других методов, приводит к тому же самому выводу. Увеличение средней скорости движения зубаток в указанном выше ряду, очевидно, связано с уходом на глубины, а конкретнее с уменьшением числа убежищ на дне, с увеличением протяженности кормовых и нерестовых миграций, с возрастанием роли подвижных организмов в питании. Предположение, что снижение удельного веса при отсутствии плавательного пузыря, с одной стороны, облегчает, а с другой — затрудняет выработку большей скорости движения, также представляется нам весьма вероятным.
Дополнение к разделу «форма тела, плавники». У пелагических мальков усеченность хвостового плавника и относительная мощность hypuralia в общем увеличивается в ряду Anarhichas latifrons — Anarhichas minor — Anarhichas lupus lupus. На данном этапе развития форма хвостового плавника тесно связана со степенью слияния непарных плавников: в том же ряду увеличивается разрыв между лучами анального плавника и лучами хвостового, а укорочение задних лучей спинного и анального плавников становится более заметным. Подвижность мальков одинаковых размеров, по-видимому, возрастает, что согласуется с характером питания, насколько о последнем можно судить по различиям в величине зубов. Во время перехода к питанию у дна ряд, в котором усеченность хвостового плавника увеличивается, меняется на обратный: Anarhichas lupus lupus — Anarhichas minor — Anarhichas latifrons.
«Змееобразное» движение, по устному сообщению О. Н. Киселева, наблюдавшего Anarhichas lupus lupus из гидростата, типично для этой рыбы и в естественной обстановке. Плавают Anarhichas lupus обычно над самым дном, на высоте 20—40 см от него, и редко на высоте 1—2 м. Отдыхая на грунте, рыба пригибает хвост к боку; иногда «посматривая», приподнимает голову. Отрывая моллюска от субстрата, зубатки извиваются, скручиваются. Часто двигаются вперед или пятятся назад по дну в наклонном положении, «прощупывая» дно лучами грудных плавников и разрывая грунт рылом. В связи с указанными особенностями поиска пищи покровы нижних лучей грудного плавника у Anarhichas lupus lupus сильно утолщены; сходное утолщение указанных лучей наблюдается у Anarhichas minor и Anarhichas latifrons, но степень его уменьшается, особенно у Anarhichas latifrons, и более всего у особей этого вида, меньших по размерам (60—70 см).
Кровеносная система.
В ряду Anarhichas lupus maris-albi — Anarhichas lupus lupus — Anarhichas minor — Anarhichas latifrons относительный вес сердца снижается (табл. 9).

Возрастной и половой изменчивости в этом индексе не обнаружено; его постоянство на всех стадиях роста, по Гессе (Hesse, 1921 : 257), вообще свойственно рыбам в отличие от других позвоночных.
Относительный вес сердца (без венозного синуса и луковицы аорты) у Anarhichas lupus lupus, исследованных Киттелем и Кэмпфе (3 экз.; Kittel и. Kampfe, 1952-1953 : 474), равнялся 0.07—0.08%, тогда как у Sebastes viviparus, трески и пикши в среднем — 0.09—0.10%. Киттель и Кэмпфе считают, что малый вес сердца Anarhichas lupus объясняется ее образом жизни, а именно питанием малоподвижными организмами. Однако в приведенном выше ряду зубаток питание подвижными организмами возрастает, а относительный вес сердца тем не менее снижается. Дело обстоит, надо полагать, гораздо сложнее, чем это нам представляется, так как на относительный вес сердца могут влиять, кроме характера плавательных движений, многие другие причины, в частности особенности строения околосердечной сумки и жаберного аппарата и т. д. (Пучков, 1954 : 45). Живучесть сердца зубаток изумительна. Сердце сокращается — при прикосновении к нему — иногда через несколько часов после того, как зубатка, вынутая из воды, перестает показывать признаки жизни. Даже вырезанное из тела и пробывшее до получаса на воздухе, сердце отвечает иногда сокращением на прикосновение, особенно к желудочку; на прикосновение к луковице аорты сокращением не отвечает.
По Криспу (Crisp, 1853 : 465), кровяные тельца зубаток не отличаются от кровяных телец многих других исследованных им рыб. Вес селезенки колеблется в пределах 0.12—0.32% от веса тела и в среднем немного повышается в ряду Anarhichas lupus maris-albi — Anarhichas lupus lupus — Anarhichas minor — Anarhichas latifrons (0.18, 0.22, 0.22, 0.23%; число экз., как и в табл. 9). Киттель и Кэмпфе (Kittel п. Kampfe, 1952—1953 : 474) нашли относительный вес селезенки у Anarhichas lupus lupus равным 0.11—0.21, в среднем 0.16% (3 экз.), у Sebastes viviparus, трески и пикши — в среднем 0.18, 0.13, и 0.10%. Указанные авторы предполагают, что с уходом рыб на глубины относительный вес их селезенки увеличивается. В самом деле, у зубаток относительный вес селезенки, по-видимому, в среднем несколько увеличивается с возрастанием глубин, на которых эти рыбы предпочитают держаться.
Покровы.
Все виды зубаток отличаются очень толстой и прочной кожей. Прочность и толщина кожи уменьшаются в ряду Anarhichas lupus — Anarhichas minor — Anarhichas latifrons, особенно у Anarhichas latifrons. Едва ли можно сомневаться в том, что прочность и толщина кожи зубаток объясняются их приуроченностью к каменистым грунтам. Не удивительно, что в указанном ряду кожа становится менее толстой и прочной: это происходит в силу перехода на мягкие грунты и возрастания оторванности от дна в связи со смещением на глубины.
Тело зубаток всегда покрыто очень толстым слоем слизи, имеющей специфический неприятный запах. Можно предполагать, что этот запах —
одно из средств обнаружения особей своего вида. На воздухе при понижении температуры слизь становится более вязкой. Возможно, и в воде происходит то же самое, и в таком случае вязкость слизи увеличивается с возрастанием вязкости воды, сохраняя значительное превосходство над последней. Как известно (Richardson, 1936 : 67), наличие пограничного слоя, более вязкого, чем основная масса жидкости, задерживает образование вихрей. Слизь является таким пограничным слоем между телом рыбы и водой и, следовательно, должна заметно влиять на скорость движения. Однако опыты Ричардсона по скорости падения мертвых рыб привели его к выводу, что слизь почти не влияет на эту скорость. Очевидно, у живых рыб слизь действительно не влияет на скорость при движении по инерции, но при плавательных движениях, надо полагать, возможность вихреобразования сильно возрастает и роль слизи проявляется гораздо отчетливее. Вряд ли случайно у зубаток, обладающих большой амплитудой плавательной волны, слой слизи отличается такой значительной мощностью.
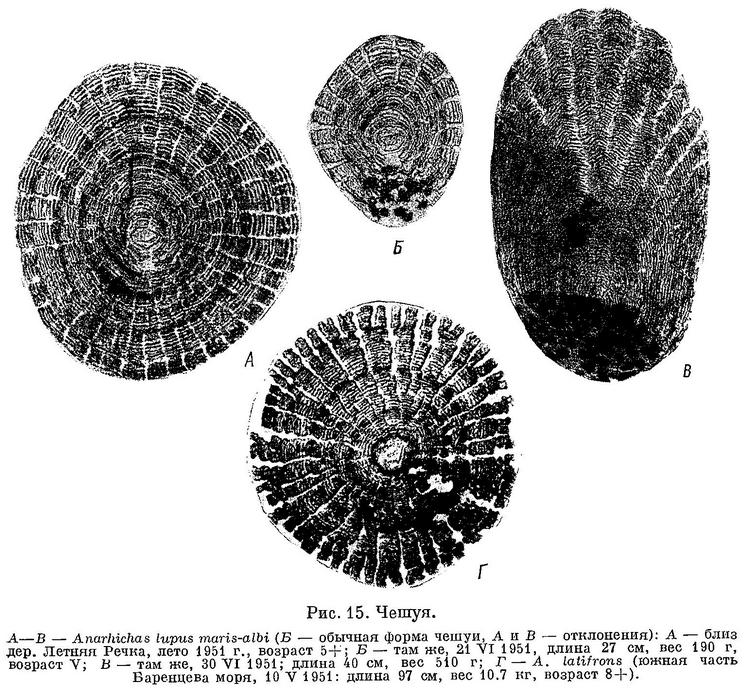
Чешуя очень мелкая и тонкая. На голове ее нет. На теле, особенно на брюхе, чешуйки сидят редко. На хвосте они мельче и сидят гуще, чем на туловище, но и здесь не соприкасаются друг с другом. Чешуйка имеет вид широкого полого конуса без основания с округленной вершиной, вокруг которой по бокам конуса располагаются концентрические кольца склеритов, разорванные радиальными каналами. Гораздо более широкий разрыв между склеритами, чем в области радиальных каналов, обычно покрытый пигментными клетками, заметен на каудальном секторе чешуи (рис. 15, Б). Величина указанного разрыва у всех видов зубаток сильно варьирует, а в среднем снижается в ряду Anarhichas lupus — Anarhichas minor — Anarhichas latifrons. Иногда его вообще не бывает (рис. 15, А). Этот разрыв несколько напоминает каудальный сектор ктеноидной чешуи (но лишен шипиков) особенно у одной видоизмененной чешуйки, найденной у Anarhichas lupus maris-albi (рис. 15, В).
Закладка чешуи происходит поздно: большинство сеголеток Anarhichas lupus lupus и Anarhichas lupus maris-albi длиной 6—7 см и сеголеток Anarhichas orientalis длиной 15 см имели на чешуе всего лишь по два склерита. Величина чешуек непостоянна: диаметр одной из них может до двух раз превышать диаметр другой, взятой с того же участка тела. Чешуйки, расположенные вдоль нижней ветви боковой линии, разделены более широкими промежутками, чем остальные, и несколько вытянуты дорзовентрально.
Форма чешуи у рыб должна быть тесно связана с характером плавательной волны. Этот вопрос, насколько мне известно, почти не затронут исследованиями. Как бы то ни было, чешуя у зубаток утончается и не перекрывается в связи с чрезвычайно большой гибкостью их тела, не только латеральной, но и дорзовентральной. Неправильно недооценивать последнюю, как это делает Гринберг (1950).
Окраска
Для всех зубаток характерно преобладание серых и бурых тонов в общем фоне окраски, причем интенсивность бурых наиболее изменчива, а с возрастом в общем увеличивается. Низ тела светлее боков и спины, хотя бы немного. Пестрота окраски ослабевает по направлению к хвосту и брюху. Радужина глаз серая, темно-серая или темно-бурая, с желтыми пятнами неправильной формы. Зубы вскоре после смены красные, затем постепенно теряют эту окраску, становясь желтоватыми, что зависит от изменений, происходящих в трабекулярном дентине (Lühmann, 1954а : 490). У Anarhichas lupus зубы теряют красную окраску, по-видимому, быстрее, чем у других атлантических видов.
У Anarhichas lupus maris-albi общий фон окраски серый с бурыми оттенками разной интенсивности. Бока и спина опоясаны буровато-коричневыми или темно-бурыми, почти черными, полосами, продолжающимися на спинной плавник. На боках их 9—13, на спине 8—12. Полосы на спине в той или иной мере сдвинуты по отношению к полосам на боках. Те и другие лучше видны в передней части тела, состоят из сливающихся между собой пятен с размытыми краями, что у взрослых выражено лучше. У молоди на спинном плавнике на продолжении полос обычно встречаются яркие черные пятна, не больше одного на продолжении каждой полосы. С возрастом пятна исчезают, начиная с задних; самым ярким и стойким всегда оказывается первое от головы пятно. Остальные плавники обычно без пятен и полос. У молоди между полосами на спине и в нижней части спинного плавника имеются пятна того же цвета, что и полосы, отделенные друг от друга и от полос белыми с желтоватым оттенком ленточками, особенно яркими у основания плавника. С возрастом эти пятна и ленточки сливаются в один общий фон. На основании спинного плавника и на спине ниже него, на полосах и между ними, у молоди бывают еще мелкие темные, часто почти черные, пятна, относящиеся, по-видимому, к поверхностной окраске; у одного из 2000 экз. они покрывали все тело, включая плавники. У взрослых особей эти пятна расплываются, бледнеют и сливаются с общим фоном. Интенсивность бурых тонов в окраске может заметно меняться в течение всего нескольких часов. В общем у взрослых общий фон и полосы темнее, полосы менее заметны, чем у молоди.
Покровительственный характер окраски вряд ли может вызвать сомнения. Поперечные полосы на боках и сдвинутые по отношению к ним такие же полосы на спине и спинном плавнике прекрасно имитируют разветвленные талломы фукусов, в зарослях которых молодь Anarhichas lupus maris-albi часто держится летом во время приливов. Пятна на спинном плавнике могут сойти за спорангии. По наблюдениям Фабрициуса (Fabricius, 1780 : 189), даже отдыхающая в расселинах скал зубатка (Anarhichas lupus lupus) производит телом волнообразные движения, что должно еще больше усиливать сходство ее полос с колеблющимися водорослями. Таким образом, приспособительная окраска сочетается в данном случае со специализированным поведением (Cott, 1940 : 433). Вне зарослей молодь может принимать однотонную окраску, так как промежутки между полосами темнеют сильнее, чем сами полосы. Окраска взрослых особей гораздо однороднее: по всей вероятности, они избегают зарослей.
Желтые пятна на радужине глаз становятся больше, если живую или недавно погибшую зубатку выставить на солнце.
У всех зубаток намечается один и тот же порядок возрастной изменчивости в окраске. Вначале появляются полосы, позднее наблюдается деление их на пятна, начинающееся с передней части туловища. Пятна в дальнейшем исчезают, сливаясь с темнеющим общим фоном. Покровительственная окраска достигается видоизменением какой-либо стадии, подавлением второй стадии (Anarhichas lupus), незавершенностью (Anarhichas lupus, Anarhichas latifrons) или полным выключением (Anarhichas minor, Anarhichthys ocellatus) конечной стадии.
Из вышеизложенного видно, что при всей своей изменчивости окраска зубаток отличается значительным постоянством, особенно в расположении полос и пятен на теле, которое давно служило и служит до сих пор хорошим систематическим признаком для определения видов зубаток.
Дополнение к разделу «Окраска». У пелагических мальков всех атлантических видов зубаток меланофоры наиболее рассредоточены на нижней стороне головы и на брюхе. Яркость окраски у мальков, меньших по размерам, несколько увеличивается по направлению к хвосту, а не к голове, как у больших по размерам; однако область hypuralia вначале остается свободной от пигмента и покрывается им лишь по мере роста мальков. Обнаружены следы трех продольных полос, расположенные вблизи оснований спинного и анального плавников и посередине тела. Мальки Anarhichas lupus lupus и Anarhichas latifrons окрашены довольно однообразно, тогда как мальки Anarhichas minor отличаются яркой поперечнополосатой окраской. Брюшина у мальков всех трех видов черная. Окраска глаз у живых — яркая голубоватая, как и у мальков морских окуней (по устному сообщению А. С. Бараненковой). У особей Anarhichas latifrons длиной 67 см и более радужина испещрена желтыми пятнами; с возрастанием размеров рыб относительная величина пятен уменьшается. Судя по этим изменениям, с увеличением размеров рыбы размах ее вертикальных миграций сокращается, что отражается и на окраске брюшины: последняя изменяется от темно-серой до чуть сероватой. У всех Anarhichas minor близких размеров брюшина светлая.
Органы чувств
Основное внимание в данном разделе уделено глазам и органам боковой линии; меньше, чем следовало бы, удалось поработать с органами слуха и обоняния. Изучая строение органов чувств и мозга, мы вступаем в такую среду, где особо остро чувствуется потребность непосредственного наблюдения над живыми рыбами, а также дополнения макроскопического исследования микроскопическим. Органы вкуса и осязания совершенно не затронуты именно вследствие того, что требуют главным образом гистологического исследования.
Глаза. Орбита у зубаток ограничена сверху frontale и хрящевым козырьком, расположенным над глазом вдоль краев этой кости, спереди — мезэтмоидом, снизу и сзади — окологлазничным кольцом, а также соединительнотканной перепонкой, натянутой между suborbitalia, парасфеноидом и нижним отростком frontale. Перепонка эта эластична; при открывании рта вследствие расслабления А3 она «проваливается» и глаза втягиваются внутрь орбит. Орбиты разделены межглазничной перегородкой, частью хрящевой, частью соединительнотканной. В ряду Anarhichas latifrons — Anarhichas minor — Anarhichas lupus — Anarhichas orientalis соединительнотканная часть перепонки увеличивается за счет хрящевой.
Положение глазного яблока во многом определяется степенью развития челюстного аппарата и его мышц. К задней части окологлазничного кольца прикрепляется поверхностная порция m. adductor mandibulae (A1); передняя часть кольца — praeorbitale — служит ограничителем, не позволяющим maxillare смещаться вверх при закрывании челюстей. Ширина межглазничного пространства, ввиду необходимости сохранения обтекаемой формы головы, до некоторой степени зависит от ширины заглазничного пространства, определяемой степенью и характером развития верхних отделов А3. Большая толщина А3 в области прикрепления A1 к окологлазничному кольцу создает наклон всего кольца вперед и вверх. Укорочение челюстей спереди, необходимое для увеличения силы их сжатия, приближает глаза к переднему краю головы. Значительная мощность челюстей спереди (praemaxillare) и снизу (dentale) и жевательных мышц сзади, при необходимости сохранения обтекаемости общих контуров головы, обусловливает сокращение орбиты спереди, а особенно снизу и сзади (Ланшина, 1928).
Глаза у зубаток расположены вблизи верхнего края головы, примерно в 2 раза ближе к ее переднему концу, чем к заднему. Диаметр радужины составляет 0.7—0.8 диаметра наружной части глазного яблока. Диаметр зрачка в среднем приблизительно равен половине диаметра радужины; при перемещении из полутьмы на яркий солнечный свет диаметр зрачка может увеличиться до 1.5 раз, что наблюдается некоторое время даже у погибшей зубатки (Anarhichas lupus maris-albi). Плоскость роговиц почти параллельна боковой плоскости окологлазничного кольца и наклонена вперед и вверх. Наклон этот настолько значителен, что наличие широкого поля бинокулярного зрения впереди и сверху головы вряд ли может ставиться под сомнение. Наличие его создает возможность точного расчета движений при помощи зрения. С другой стороны, наклон роговиц вперед и вверх сильно сокращает поле зрения снизу и сзади. Для зубаток, ведущих придонный образ жизни, это, по-видимому, не имеет большого значения, так как снизу опасность им вряд ли грозит.
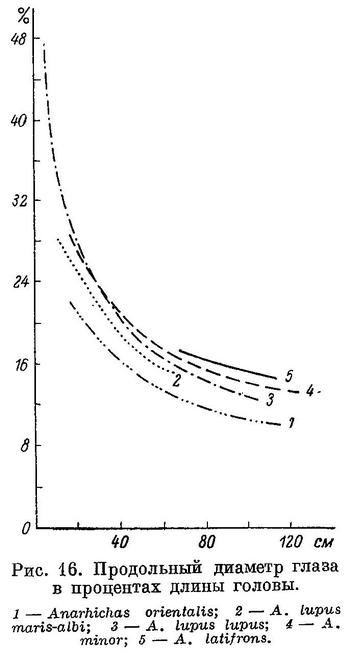
Возрастная изменчивость очень интенсивна и у всех видов сходна: в течение всей жизни относительные размеры глаз сильно уменьшаются (рис. 16), а угол наклона роговиц вверх и вперед возрастает. Этот процесс идет особенно быстро в начале онтогенеза, постепенно замедляясь к его концу.
У личинок Anarhichas lupus lupus, выходящих из икры, диаметр глаза составляет около половины длины головы и около 0.7 ее высоты через середину глаза. Плоскости роговиц почти параллельны друг другу. Глаза выдаются вбок дальше любой из точек тела. Их верхний край находится на уровне верхнего края головы, а передний — почти на уровне ее переднего края. Поля зрения должны быть очень обширными и при движении глаз, по-видимому, охватывают все пространство сверху, спереди, с боков и снизу головы, в значительной мере даже и сзади нее. По всей вероятности, зрение у личинок служит в основном для предупреждения о приближающемся враге, который может находиться и снизу личинок в то время, когда они поднимаются над дном (Mcintosh a. Prince, 1890 : 880): для зубаток такого размера могут быть опасны даже морские звезды — их будущая жертва. Челюсти и жевательные мышцы в это время развиты очень слабо, что и делает возможным наличие такого обширного поля зрения.
В дальнейшем происходит увеличение относительных размеров челюстей и связанных с ними мышц, сопровождающееся изменениями их формы, а также и формы неврокраниума. Эти процессы происходят особенно быстро в начале онтогенеза, постепенно замедляясь к его концу. Их связь с изменениями относительных размеров, формы и расположения глаз несомненна. Вполне очевидно, что при увеличении челюстей и связанных с ними мышц, при сужении заглазничного отдела черепа сохранение прежнего размера глаз относительно всей головы и прежнего положения плоскости роговиц невозможно без коренного нарушения обтекаемости тела; значение же последней в силу увеличения подвижности возрастает. В действительности уже на первом году жизни происходит быстрое сокращение относительных размеров глаз, а также поворот их роговиц вперед и вверх. Ввиду уменьшения относительных размеров глаз возможность расширения поля зрения с помощью их движений снижается. Поля зрения резко сокращаются снизу и сзади, но увеличивается возможность бинокулярного зрения вверх и вперед. Однако особо важно направление его вперед и вниз, а не вверх и вперед, что доказывается изменением формы наружных частей глаза, включая радужину и зрачок, из круглой в продолговатую, несколько вытянутую вперед и вниз. Отсюда следует, что у Anarhichas lupus бинокулярность зрения особенно важна при добывании пищи. Такая довольно редкая форма наружных частей глаза интересна в другом отношении: она показывает, что положение окологлазничного кольца, определяемое во многом мощностью и расположением челюстей и жевательных мышц, не обеспечивает надлежащего направления поля зрения при обычной круглой форме наружных частей глаза.
У Anarhichas minor глаза менее продолговаты, а у Anarhichas latifrons обычно круглые. Такое различие в форме глаза указывает на уменьшение роли зрения при добывании пищи со дна в ряду Anarhichas lupus — Anarhichas minor — Anarhichas latifrons.
Что касается Anarhichas orientalis, то удлинения наружных частей глаза у нее не наблюдается, по-видимому, по иной причине, чем у Anarhichas latifrons: наклон роговиц вперед настолько велик, что такое удлинение не является необходимым. Наклон роговиц вперед и вверх в общем возрастает в ряду взрослых особей Anarhichas latifrons — Anarhichas minor — Anarhichas lupus lupus — Anarhichas lupus maris-albi — Anarhichas orientalis.
Из вышеизложенного следует, что в указанном ряду возрастает роль зрения при добывании пищи со дна, но снижается предупреждающая роль при защите от врагов, нападающих снизу и сзади. Все это вполне согласуется с предпочтением зубатками меньших глубин и возрастанием привязанности ко дну, наблюдающейся в данном ряду.
В ряду Anarhichas latifrons — Anarhichas minor — Anarhichas lupus lupus — Anarhichas lupus maris-albi — Anarhichas orientalis среди особей одинаковой длины размер глаз относительноразмера всей головы уменьшается (рис. 16). Возможно, в том же ряду ухудшается и зрение при слабом освещении; однако для окончательного решения этого вопроса необходимо изучение внутреннего строения глаз, особенно сетчатки.
Строение мышц глаза у всех зубаток рода Anarhichas (относительно Anarhichthys сведений нет) очень сходно. Косые мышцы глаза прикрепляются к мезэтмоиду рядом с межглазничной перегородкой, внутрь от места прохождения через мезэтмоид обонятельного нерва. Верхняя, нижняя и внутренняя прямые прикрепляются к парасфеноиду в нижнем заднем углу глазниц. Наружная прямая входит в глубь миодома и прикрепляется к парасфеноиду вблизи его границы с basioccipitale. Свод миодома образован только prooticum, дно — только парасфеноидом. Basioccipitale никакого участия в образовании миодома не принимает. Таким образом, миодом зубаток имеет свои особенности и не подходит ни под один из типов, указанных Еремеевой (1928 : 115). Аллис (Allis, 1919 : 313) миодом такого типа, ограниченный областью prooticum, нашел лишь у Holostei и Crossopterigii. Вследствие наличия миодома наружная и внутренняя прямые мышцы становятся почти равными по длине. Верхняя и нижняя прямые и без того примерно одинаковы, как и косые мышцы.
У Anarhichas orientalis прямые мышцы глаза сильно удлинены, а глазница вырезана так, что край парасфеноида, ограничивающий ее сзади, располагается позади заглазничных отростков, тогда как у атлантических зубаток — примерно напротив них. У Anarhichthys ocellatus вырез этого края парасфеноида двойной. Выступ в его середине, по-видимому, отделяет наружную прямую от остальных мыщц глаза.
Боковая линия. Схема расположения каналов боковой линии, пор и сейсмосенсорных почек (генипор) у зубаток дана на рис. 18. Часть заглазничных пор, обычных у многих других Stichaeoidae, у зубаток исчезает вследствие «наплывания» на них верхних отделов «жевательных» мышц (Макушок, 1958 : 23). На схеме отмечено наиболее обычное число пор, от которого часто встречаются отступления. Так, в подглазничном канале нередко бывает не 8, а 9 пор, в надглазничном не 2, а 3; непарные поры надглазничной комиссуры и portio supratemporalis иногда бывают парными; в portio supratemporalis сверх того, кроме непарной, может быть не одна, а две парных поры; в предкрышечно-нижнечелюстном канале бывает изредка не 8, а только 7 пор; в отрезке канала боковой линии, проходящем во второй tabulare и костях плечевого пояса, — не 4, а только 3.

Число отверстий в костях для вхождения внутрь них каналов боковой линии гораздо постояннее. Вариации в числе пор происходят большей частью ввиду того, что каналы в местах прохождения между двумя соседними костями открываются наружу то одной, то двумя порами (frontale—nasale, dentale—praeoperculum) или даже тремя (между раrietalia); иногда пора может отсутствовать (posttemporale—supracleithrum). В подглазничном канале вариации в числе пор обусловлены колебаниями числа suborbitalia.
Число и расположение сейсмосенсорных почек еще менее постоянно, чем пор, но все же многие из них (отмечены на схеме крупными точками)
встречаются всегда в одних и тех же местах и примерно в том же числе.
Не удалось заметить никакого существенного различия между видами в расположении и числе пор и сейсмосенсорных почек, тем более в расположении и числе отверстий в костях, внутрь которых входят каналы боковой линии (правда, для Anarhichthys ocellatus мне известно — по рисункам и фотографиям черепов — только последнее). Заметно изменяется лишь диаметр каналов в костях для боковой линии, увеличиваясь в ряду Anarhichthys ocellatus — Anarhichas orientalis — Anarhichas lupus maris-albi — Anarhichas lupus lupus — Anarhichas minor — Anarhichas latifrons. Увеличение идет наиболее интенсивно от Anarhichas lupus к Anarhichas latifrons. Оно сопровождается возрастанием пористости костей и как-то связано с предпочтением этими видами больших глубин. Возможно, роль боковой линии в ориентировке движений увеличивается, компенсируя уменьшение роли зрения.
У всех видов рода Anarhichas на теле имеются две ветви боковой линии. Верхняя ветвь начинается сразу же за последней порой на supracleithrum и тянется вдоль спины до конца туловища или чуть далее. Нижняя начинается немного отступя от указанной поры и проходит вдоль средней оси тела почти до самого основания хвостового плавника. Американские исследователи (Jordan a. Gilbert, 1882 : 782; Jordan а. Evermann, 1898 : 2448, и др.) отрицают наличие боковой линии на теле Anarhichthys ocellatus, что, безусловно, требует проверки, так как наличие ее на теле Anarhichas orientalis ими также отрицается; более того, многие из них не находят боковой линии и у других видов зубаток, хотя ветви ее на теле Anarhichas lupus lupus были точно описаны еще Фабрициусом (Fabricius, 1780 : 138).
Ветви боковой линии на теле зубаток рода Anarhichas не имеют каналов и пор: каждая из них состоит из ряда отдельных сейсмосенсорных почек (Макушок, 1958 : 112). Эти почки очень плохо заметны не только у фиксированных, но часто и у свежих экземпляров находящихся на воздухе (у Anarhichas minor обычно совсем не видны). Однако у живых зубаток, в воде, они видны прекрасно. То же самое можно сказать и о сейсмосенсорных почках, расположенных на голове. Надо полагать, Уолтерс (Walters, 1955 : 306) исследовал фиксированных зубаток и именно поэтому не обнаружил боковой линии не теле крупных Anarhichas minor и Anarhichas latifrons. Ряд сейсмосенсорных почек, идущий к началу спинного плавника от верхней норы portio supratemporalis, Уолтерс считает дорзальной ветвью боковой линии и, таким образом, насчитывает на теле зубаток три ее ветви. Однако «дорзальная ветвь» очень коротка и ограничена в основном областью головы.
Носовые ямки. Носовые ямки у всех видов зубаток имеют по одному наружному отверстию, которое находится на конце кожной трубочки, возвышающейся над носовой полостью. Трубочки обычно направлены вбок, немного вперед и вверх. Они способны к самостоятельному движению и могут менять свое направление, обращаясь отверстиями вперед (у Anarhichas lupus lupus, — Low, 1813 : 187—188; мной наблюдалось у Anarhichas lupus lupus и Anarhichas minor). Этот рефлекс часто сопровождается раскрыванием рта и наблюдается лишь у только что пойманных зубаток.
Дно носовой полости покрыто кожными складками, лежит на связке, натянутой между nasale, мезэтмоидом и praeorbitale. Судя по большому объему обонятельных долей мозга (см. ниже), обоняние хорошо развито.
Дополнение к разделу «Носовые ямки». Наружное отверстие обонятельной трубочки у Anarhichas latifrons значительно уже, чем у Anarhichas minor и Anarhichas lupus lupus сходных размеров (рис. 41).
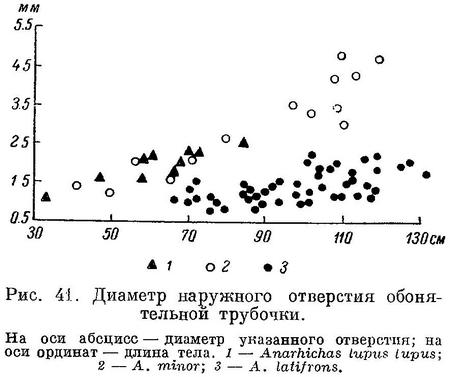
Внутреннее ухо. У Anarhichas lupus lupus sacculus и lagena широко отделены от utriculus (Retzius, 1881 : 66, taf. 10, figs 5, 6). Эту особенность Берг (1940а : 316) считает настолько важной, что выделяет ее как наиболее характерный признак семейства. В височном отделе черепа зубаток между pteroticum, epioticum и parietale имеется глубокая полость (очевидно, это подвисочная ямка, по терминологии Элиса, — Allis, 1909 : 204), ведущая к внутреннему уху и отделенная от него лишь тонкой костной перегородкой, образованной pteroticum. Сверху полость частично прикрыта первой tabulare. Такое строение черепа и внутреннего уха явно связано между собой и, может быть, определяется особенностями слуха. Интересно, что некоторые из только что пойманных зубаток издают иногда скрипящие звуки, но чем они издают их, не удалось установить. Сервисен (Sorensen, 1884 : 68) указывает, что лучи спинного плавника издают скрипящие звуки даже у мертвой Anarhichas lupus lupus, если их особенным образом подвигать (что несвойственно лучам других плавников), но сомневается, что живая зубатка сама способна производить такие движения.
Мозг
Строение мозга у всех представителей рода Anarhichas (мозг Anarhichthys ocellatus не исследовался) имеет очень много общего (рис. 19—20). Lobi olfactorii у всех сидячие. Corpora striata развиты сильно, складчатые. Эпифиз длинный и тонкий (на рисунках не изображен). Lobi optici округлые, крупные, поверхность их гладкая. Corpus cerebelli поднимается, постепенно утолщаясь с боков, выше верхнего края lobi optici, а затем изгибается вниз. Опускающаяся часть короче поднимающейся и не достигает продолговатого мозга. Eminentiae granulares небольшие, округлые. Valvula cerebelli довольно мала. Lobi vagales представляют собой лишь слабые вздутия по бокам ромбовидной ямки, a lobi acustici — впереди неё. Ромбовидная ямка длинная, превышает длину мозжечка. Зрительные нервы широкие и плоские, их перекрест происходит у взрослых особей впереди переднего мозга. Infundibulum несет направленный вперед, конусообразно заостренный гипофиз. Saccus vasculosus большой (Lissner, 1918 : 127-133).
В ряду Anarhichas latifrons — Anarhichas minor — Anarhichas lupus — Anarhichas orientalis происходит увеличение corpora striata относительно lobi optici (рис. 19). Напротив, относительные размеры lobi olfactorii снижаются. Сильно уменьшается и ширина обонятельных нервов: у Anarhichas latifrons они равны по ширине зрительным, у Anarhichas minor в 3 раза, а у Anarhichas lupus и Anarhichas orientalis в 4—5 раз уже последних.
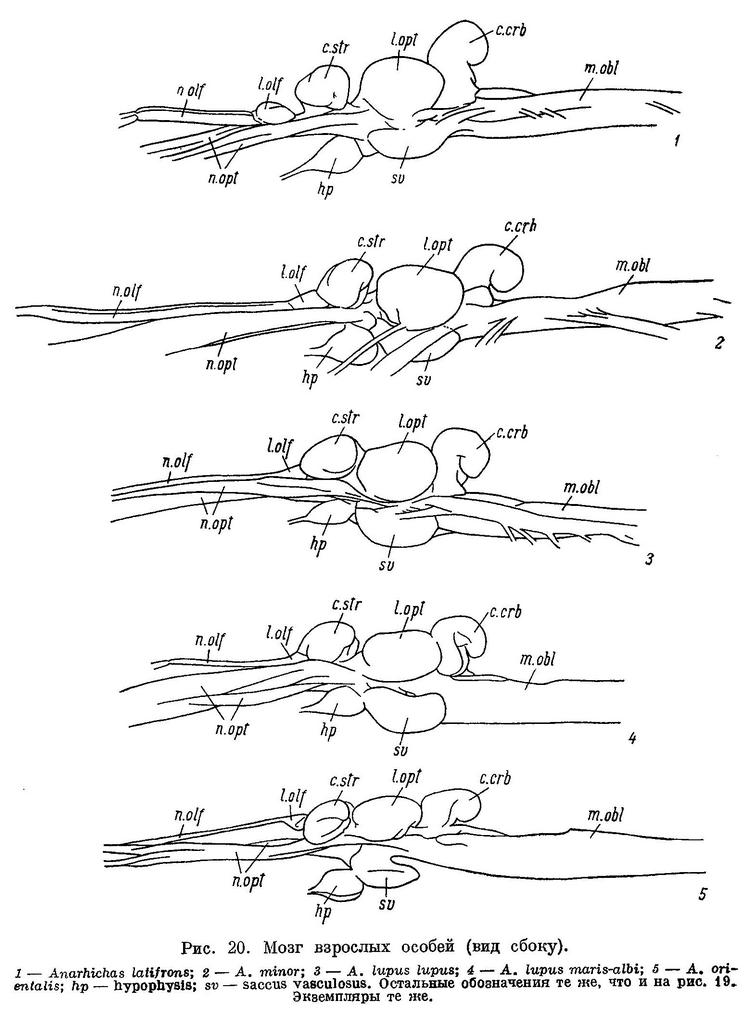
Приподнятость corpus cerebelli над верхним краем lobi optici постепенно уменьшается (рис. 20). Это происходит, очевидно, ввиду уменьшения крутизны верхнего профиля головы в области мозга. У Anarhichas lupus maris-albi тело мозжечка несколько короче и заметно уже, чем у Anarhichas lupus lupus.
Ромбовидная ямка у Anarhichas latifrons и Anarhichas minor широкая, у Anarhichas lupus и Anarhichas orientalis значительно уже. Увеличивается относительная длина участков зрительных нервов, налегающих друг на друга (на рисунках не показано, так как эти участки очень длинны, у Anarhichas lupus и Anarhichas orientalis в несколько раз превышают длину мозга). У Anarhichas lupus дорзальнее налегает левый зрительный нерв (2 экз.), у Anarhichas latifrons, Anarhichas minor и Anarhichas orientalis правый (по 1 экз.).
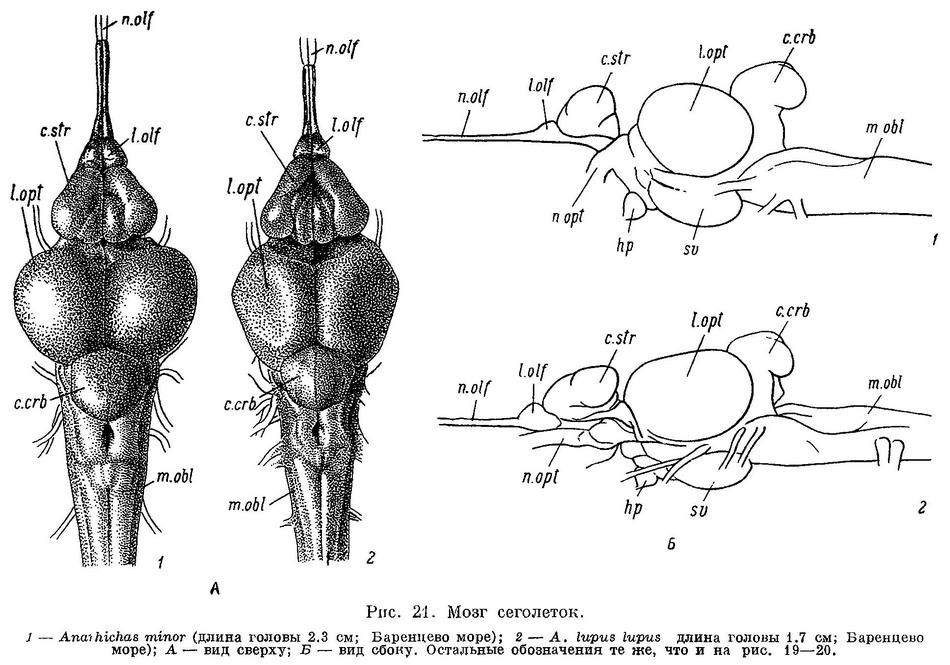
Мальки сильно отличаются от взрослых особей расположением перекреста зрительных нервов — не впереди переднего мозга, а под ним. Не возникает сомнений в том, что выдвижение хиазмы вперед вызывается сужением черепа в области прикрепления верхних отделов А3. У малька Anarhichas minor это сужение выражено слабее, мозг не отодвинут назад; зрительные нервы, отходя от промежуточного мозга к глазам, направляются сразу же вниз (рис. 21, Б, 1) — здесь и происходит их перекрест. У малька Anarhichas lupus lupus череп сужен сильнее, мозг отодвинут назад, зрительные нервы минуют край орбиты не сразу при отхожденни их от промежуточного мозга, а впереди него, примерно на вертикали lobi olfactorii, и только здесь могут опуститься вниз. Хиазма сдвигается вперед (рис. 21, Б, 2). Череп всех зубаток в области прикрепления верхних отделов А3 с возрастом становится уже. При этом мозг отодвигается от орбиты назад, зрительные нервы сближаются, располагаясь параллельно общему стволу мозга вплоть до края орбиты, где начинают расходиться и опускаться вниз. Хиазма выступает из-под переднего мозга, длина налегающих друг на друга участков зрительных нервов возрастает соответственно интенсивности сужения черепа.
У мальков ромбовидная ямка значительно короче и уже, чем у взрослых. Corpora striata впереди сужены, а не округлены. Lobi optici относительно других частей мозга развиты сильнее (рис. 21, А).
Размер мозга по отношению к общим размерам головы с возрастом уменьшается. Это уменьшение особенно велико при переходе от личинки к мальку, но продолжается в течение всей жизни (табл. 10).
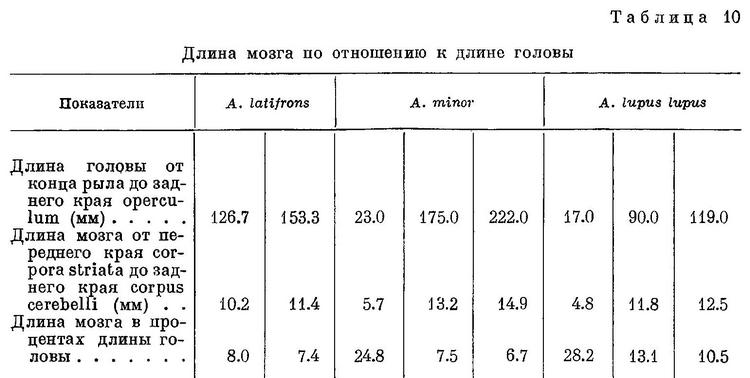
Уменьшение с возрастом размеров мозга относительно размеров головы, по всей вероятности, замедляется в ряду Anarhichas latifrons — Anarhichas minor — Anarhichas lupus.
Из рыб, исследованных Павловским и Курепиной (1953 : 134—161—182, рис. 28), мозг Blennius больше всего похож на мозг зубатки; особенно велико сходство с Anarhichas orientalis. Однако eminentiae granulares у последней развиты сильнее, ромбовидная ямка значительно длиннее и особенно велико должно быть различие в расположении зрительных нервов. По форме corpora striata на Blennius более всего похожи мальки зубаток.
Таково в настоящий момент наше знание морфологии зубаток, наше представление о связях между формой и функциями их органов. Вне всякого сомнения, завеса неизвестности, скрывающая бесконечную сложность указанных связей, пока еще только чуть приоткрыта. Тем не менее этого достаточно, чтобы ещё раз убедиться в том, что всякое изменение функции какого-либо одного органа, сопровождаясь изменением его формы не может не сказаться в той или иной степени на функциях и форме всех остальных органов. При этом возникает целый ряд противоречий, обусловленных приспособленностью органов к прежним функциям, их прежней согласованностью между собой. Более глубокое познание путей, по которым идет разрешение этих противоречий, может многое дать для понимания единства организма как целого и единства организма и среды, для правильной оценки биогенетического закона, значения рудиментарных органов и т. д. Безусловно исследования подобного рода во многом помогут и в деле направленного изменения организмов. Наибольшие результаты в этой области могут быть достигнуты при комплексном изучении взаимодействия органов, тканей, наконец клеток и межклеточных веществ в связи с их частной функцией и с общей жизнедеятельностью организма. В комплексе должны участвовать ученые различных специальностей — от разрабатывающих вопросы механики до занимающихся изучением высшей нервной деятельности.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ, РАССЕЛЕНИЕ И ЭВОЛЮЦИЯ
Распространение семейства зубаток амфибореальное: его представители обитают в северных частях Атлантического и Тихого океанов, будучи разобщены громадными пространствами Северного Ледовитого океана, не населенными зубатками, за исключением областей, соседних с Атлантическим океаном (рис. 22).

В северной части Тихого океана имеется два вида зубаток, резко отличающихся друг от друга по форме тела и принадлежащих к двум, различным родам: Anarhichas и Anarhichthys. Anarhichas orientalis встречается от северного побережья острова Хоккайдо до северной части Охотского моря и вдоль восточного побережья Камчатки по крайней мере до залива Корфа, а также у Курильских, Командорских и Алеутских островов, у островов Прибылова и к северу от последних до залива Нортон. Anarhichthys ocellatus обитает вдоль тихоокеанского побережья Америки от острова Кадьяк на юг до Сан-Диего. Таким образом, ареалы обоих видов соприкасаются, но не перекрещиваются: оба тихоокеанских вида как бы взаимно исключают друг друга. Оба вида обитают у берегов, предпочитая каменистые грунты.
Совершенно иное наблюдается в северной части Атлантического океана. Здесь имеется три вида зубаток, принадлежащих к тому же роду Anarhichas, что и дальневосточная зубатка. Все три вида довольно близки между собой по форме тела. Большая часть ареалов этих видов совпадает.
Далее других видов на юг проникает Anarhichas lupus — до северо-западных берегов Франции по европейскому побережью и до Лонг-Айленд-Саунд по американскому. Anarhichas lupus приурочена в основном к каменистым побережьям, но встречается и далеко от берегов, в открытом море. Особенно широко она распространена в мелководном и теплом Северном море, где другие виды зубаток очень редки. На север этот вид, очевидно, нe распространяется так далеко, как два других вида — Anarhichas minor и Anarhichas latifrons. Ареалы последних совпадают почти полностью, насколько это в настоящее время известно. На юг оба вида распространены в основном до Фарерских островов и Ново-Шотландских банок, на север — до западного побережья Шпицбергена и возвышенности Персея. Распространение зубаток вдоль северного побережья Канады изучено еще весьма слабо. Anarhichas minor всегда держится на несколько больших глубинах, менее привязана к твердым грунтам, чем Anarhichas lupus, a Anarhichas latifrons держится еще глубже, предпочитая мягкие илистые грунты. Приуроченность атлантических видов зубаток к различным местообитаниям подтверждается значительными различиями в их окраске. В одном и том же районе моря эти виды, очевидно, населяют различные биотопы и слабо сталкиваются между собой, чем, по-видимому, и объясняется совпадение их ареалов.
Ареал каждого из атлантических видов, возможно, непрерывен и от побережья Европы через порог Уайвилла Томпсона, Фарерско-Исландский порог, проливы Датский и Дэвиса простирается до побережья Америки. Связь между стадами, обитающими у побережья Европы, Фарерских островов, Исландии, Гренландии и Северной Америки, очевидно, наиболее тесна у Anarhichas latifrons, предпочитающей наибольшие глубины и совершающей, надо полагать, наибольшие передвижения. У Anarhichas minor эта связь, по-видимому, не столь велика, и наименее всего она должна быть выражена у Anarhichas lupus, предпочитающей наименьшие глубины и совершающей, судя по всему, наименьшие передвижения. Тем не менее у Anarhichas lupus lupus до сих пор не отмечено значительных морфологических отличий между особями, обитающими в различных частях ареала.
Как известно, североатлантическая фауна рыб и других групп животных ведет начало от более древней тихоокеанской. Однако из этого общего правила имеются весьма значительные исключения, в частности тресковые и сельдевые (Scheurmg, 1930 : 197, и др. — цит. по: Световидов, 1948 : 42; 1952 : 50). Таким же исключением, по предположению Шмидта (1948 : 111) и Андрияшева (1949 : 208), является и семейство зубаток.
Тресковые и сельдевые включают в себя большое количество родов и видов. Численное преобладание североатлантических видов и родов и отсутствие самобытности тихоокеанской фауны в данном случае доказывают североатлантическое происхождение указанных групп. Кроме того, у тресковых оно подтверждается вескими палеонтологическими доказательствами. Семейство же зубаток насчитывает очень небольшое число видов, а ископаемые остатки его представителей чрезвычайно скудны, что сильно затрудняет решение вопроса о месте возникновения и расселении семейства.
Преобладание североатлантических видов зубаток над северотихоокеанскими и, по-видимому, большая численность каждого из североатлантических видов по сравнению с тихоокеанскими, как будто указывают на атлантическое происхождение зубаток. Однако наличие эндемичного рода Anarhichthys в Тихом океане порождает сомнение в достоверности такого заключения. Различия в форме тела, в числе позвонков и лучей в спинном и анальном плавниках между этим родом и родом Anarhichas, представители которого имеются в обоих океанах, настолько велики, что подобное трудно встретить в каком-либо другом современном семействе рыб. Выделение Anarhichas latifrons в особый род Lycichthys (Gill, 1911 : 162—163), с которым трудно согласиться, только уравновешивает количество эндемичных родов в обоих океанах.
Результаты морфологического исследования зубаток могут до некоторой степени помочь в решении этого сложного вопроса.
Эндемичный тихоокеанский род Anarhichthys имеет чрезвычайно удлиненную угреобразную форму тела, большое число позвонков, лучей в спинном и анальном плавниках, а также слитые непарные плавники.
У представителей рода Anarhichas число позвонков и лучей в спинном и анальном плавниках сильно уменьшено, форма тела близка к торпедообразной, но сохранение большой и слабо увеличивающейся к хвосту амплитуды плавательной волны, очень большая боковая и дорзовентральная гибкость тела, редукция чешуи и брюшных плавников, частичная редукция мускулатуры хвостового плавника, надо думать, указывают на происхождений данного рода от рыбы с угреобразной формой тела. Редукция чешуи особенно характерна для рыб с угреобразной формой тела и редка у высокотелых рыб (Гринберг, 1950 : 452). Это предположение подтверждается также неполной обособленностью непарных плавников у представителей рода Anarhichas; вполне вероятно, что перепонки, соединяющие указанные плавники, являются остатками прежнего, гораздо более полного, их соединения.
Тип строения зубного аппарата, характерный для зубаток, как было видно из вышеизложенного, обусловливает форму головы, «треугольную» в поперечном сечении, узкую сверху и широкую снизу. Надо полагать, такой тип строения зубного аппарата мог сформироваться лишь у рыб, уже имевших подобную форму головы, т. е. у рыб с высоким удельным весом, лишенных плавательного пузыря. Это были придонные рыбы побережий (см. ниже), вероятнее всего, отличающиеся угреобразной формой тела, при которой несовершенство обтекаемости тела компенсируется большим коэффициентом полезного действия (Шулейкин, 1934 : 1182). Правда, более далекие предки зубаток, возможно, обладали формой тела, «обычной» для окунеобразных рыб. Они несомненно имели брюшные плавники, расположенные на груди; их чешуя, возможно, была ктеноидной. В наследство от дальних предков зубаткам достались тазовые кости и, может быть, разрыв между склеритами, иногда напоминающий задний сектор ктеноидной чешуи, лишенный зубчиков. Превращение формы тела этих предков в угреобразную или близкую к ней, надо полагать, являлось предысторией семейства зубаток, да скорее всего и не одних зубаток. Формирование основных черт семейства произошло, очевидно, уже после того, как непосредственные предки зубаток совершили указанное превращение. Приближение формы тела у рода Anarhichas к «торпедообразной», таким образом, явление вторичное. Едва ли можно сомневаться в том, что исходный вид, давший начало семейству, имел слабо дифференцированный зубной аппарат. Сочетание двух «примитивных» черт: очень удлиненной угреобразной формы, тела и довольно слабо дифференцированного зубного аппарата наблюдается лишь у молоди Anarhichthys ocellatus.
Anarhichthys ocellatus несомненно является прибрежным видом, а возможно, и самым прибрежным из видов зубаток. Происхождение же семейства от прибрежной формы очевидно, так как из всего семейства только два вида — Anarhichas minor и Anarhichas latifrons — обитают вдали от берегов, но и они несут явные следы происхождения от прибрежных предков. Расположение полос на теле и пятен на спинном плавнике у молоди Anarhichas minor настолько близко к таковому у молоди Anarhichas lupus, что не остается никакого сомнения в происхождении Anarhichas minor от поперечнополосатого, следовательно, прибрежного предка, общего с Anarhichas lupus (возникновение поперечнополосатой окраски, имитирующей бурые водоросли, на глубинах немыслимо). Группировку пятен на теле в полосы можно зачастую заметить и у взрослых особей Anarhichas minor. Широкие полосы на теле молоди похожи на крупные пятна, группировка пятен в полосы у взрослых выражена нерезко: сохранение некоторых примитивных черт, остатков окраски предка не мешает, по-видимому, приспособлению к новой обстановке у дна, лишенного растительности. Бледные пятна на теле Anarhichas latifrons, временами неясно группирующиеся в поперечные полосы, указывают, что и этот вид при своем формировании прошел тот же путь перехода от берегов на большие глубины, но зашел по этому пути еще дальше. Не только расположение пятен в виде полос, но и сами пятна являются у него остатками окраски предка.
Anarhichthys ocellatus, очевидно, наиболее теплолюбивый из видов зубаток, так как распространяется на юг вдоль побережья Калифорнии до Сан-Диего, т. е. до границы с тропической фауной, придерживаясь при этом довольно небольших глубин. Время нереста прибрежных зубаток, распространенных севернее, свидетельствует о их происхождении от теплолюбивого предка: Anarhichas orientalis и Anarhichas lupus maris-albi нерестятся летом. Наблюдения над Anarhichas lupus maris-albi показывают, что на определенном этапе развития гонад (II стадия и по крайней мере начало III) этому виду требуются относительно высокие температуры. Такой же вывод намечается и для зубаток, ушедших на глубины, а именно для Anarhichas minor.
Все эти данные склоняют нас к выводу о тихоокеанском происхождении семейства от вида, близкого по форме тела и распространению к Anarhichthys ocellatus. Для многих рыб с угреобразной формой тела характерен ночной образ жизни (Tsukamoto a. Yoshino, 1957 : 133). Возможно, он был свойствен и предку зубаток, сохраняясь в дальнейшем на протяжении всей истории семейства, несмотря на изменения в форме тела: прибрежные виды Anarhichas до сих пор в какой-то степени отличаются ночным образом жизни. Ночной образ жизни зубаток мог облегчить уход части особей на глубины, обусловивший образование видов, обитающих вдали от берегов.
Из представителей рода Anarhichas наиболее удлиненную форму тела, а также наибольшее число позвонков, лучей в спинном и анальном плавниках и малые размеры хвостового плавника имеет Anarhichas orientalis. Показательно, что по числу позвонков и лучей в указанных плавниках Anarhichas orientalis отличается от других представителей рода заметно сильнее, чем все они друг от друга. К тому же этот вид имеет сходную с Anarhichthys ocellatus, но резко отличающую его от других представителей рода Anarhichas продольнополосатую окраску у молоди. Подобное же явление наблюдается и в возрастной изменчивости некоторых признаков: например, размер головы относительно размеров всего тела у обоих тихоокеанских видов с возрастом увеличивается, а у атлантических — уменьшается (см. диагнозы видов). Сближение Anarhichas orientalis в признаках, наиболее резко отличающих данный вид от других представителей рода, с Anarhichthys ocellatus дает возможность полагать, что формирование рода Anarhichas произошло также в пределах Тихого океана.
Из атлантических зубаток по числу позвонков и лучей в спинном и анальном плавниках к Anarhichas orientalis наиболее близка Anarhichas latifrons. Anarhichas minor по очень многим признакам стоит ближе к Anarhichas lupus, чем к Anarhichas latifrons. Поэтому можно полагать, что обособление Anarhichas minor и Anarhichas lupus друг от друга произошло позднее, чем обособление Anarhichas latifrons от их общего предка. Подвиды Anarhichas lupus — Anarhichas lupus lupus и Anarhichas lupus maris-albi — настолько близки, что их еще более позднее образование не вызывает сомнений.
Как видно из вышеизложенного, предполагается, что древность происхождения уменьшается в ряду Anarhichthys ocellatus — Anarhichas orientalis — Anarhichas latifrons — Anarhichas minor — Anarhichas lupus lupus — Anarhichas lupus maris-albi. В этом же ряду закономерно уменьшаются различия в числе лучей в спинном и анальном плавниках, а также, за небольшим исключением, и в числе позвонков (табл. 7).
Возникает вопрос: почему при обсуждении филогении зубаток числу позвонков и лучей в спинном и анальном плавниках придается такое большое значение? Дело в том, что формирование позвонков и лучей плавников, прежде всего спинного и анального, у Anarhichas lupus lupus происходит еще в икринке и поэтому мало подвержено влиянию окружающей среды. В дальнейшем число их уже не изменяется в течение всей жизни, в отличие, например, от числа зубов, меняющегося с каждым годом. Большая консервативность числа лучей в спинном и анальном плавниках у Anarhichas lupus lupus подтверждается исключительным сходством колебаний указанных признаков как у личинок из одной и той же кладки, так и у взрослых особей из разных мест Баренцева моря — пределы колебаний, коэффициенты вариации и средние арифметические почти совпадают (Барсуков, 1955 : 715). Поскольку у других видов зубаток икринки по размерам и строению близки к таковым у Anarhichas lupus lupus (неизвестна лишь икра Anarhichthys ocellatus), можно предполагать, что формирование позвонков и лучей в спинном и анальном плавниках еще в икринке было свойственно зубаткам издавна и поэтому число указанных частей тела изменялось в филогенезе медленно.
Таким образом, довольно сходные по степени и характеру специализации зубного аппарата и связанных с ним структур Anarhichthys ocellatus, Anarhichas orientalis и Anarhichas lupus оказываются менее родственными между собой, чем сильно различающиеся в данном отношении Anarhichas lupus, Anarhichas minor и Anarhichas latifrons, из которых Anarhichas minor и особенно Anarhichas latifrons имеют к тому же более примитивное строение зубного аппарата. Однако близкое родство атлантических зубаток подтверждается не только числом позвонков и лучей в спинном и анальном плавниках, но и многими другими признаками, по которым эти виды образуют один и тот же ряд, где Anarhichas minor постоянно оказывается промежуточным звеном между Anarhichas lupus и Anarhichas latifrons. С другой стороны, Anarhichas orientalis, несмотря на сильную специализацию зубной системы, сохраняет некоторые примитивные черты, не свойственные Anarhichas lupus. К их числу относится длинное сухожилие, идущее от m. adductor mandibulae к переднему концу maxillare, имеющееся также у Anarhichas minor и Anarhichas latifrons, но отсутствующее у Anarhichas lupus. Anarhichas orientalis стоит ближе к Anarhichas latifrons и Anarhichas minor, чем к Anarhichas lupus, и по некоторым другим признакам: по расположению зрительных нервов, по форме opisthoticum (табл. XXI) и т. д. Форма заднего края глазниц, очень близкая у Anarhichas lupus, Anarhichas minor и Anarhichas latifrons, у Anarhichthys ocellatus, Anarhichas orientalis и Anarhichas lupus различна (табл. XVII), хотя по относительному размеру глаз последние три вида отличаются друг от друга не больше, чем первые. Нам представляется, что сходство между Anarhichthys ocellatus, Anarhichas orientalis и Anarhichas lupus в строении зубного аппарата и связанных с ним структур вызвано более параллелизмом развития, чем родственной близостью. Для окончательного решения этого вопроса безусловно потребуются более детальные исследования зубаток, в особенности их молоди, а также взрослых особей тихоокеанских видов.
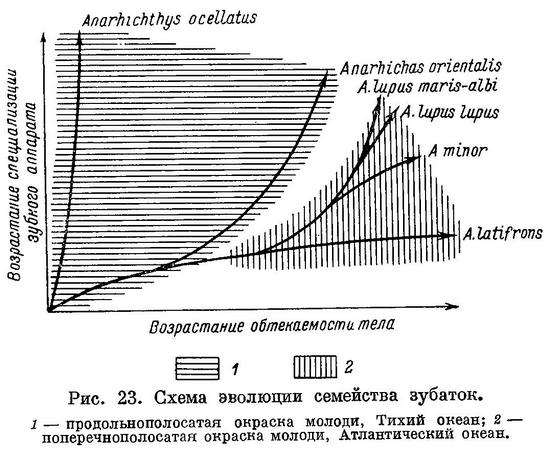
Обобщая все сказанное, можно следующим образом представить себе предварительную схему эволюции семейства зубаток (рис. 23). Исходный вид, давший начало семейству, имел угреобразную форму тела и слабо дифференцированный зубной аппарат. Подобно Anarhichthys ocellatus, он был теплолюбивым прибрежным тихоокеанским видом. В северной части ареала от него когда-то обособился другой вид, давший начало роду Anarhichas. По форме тела и числу позвонков этот вид был близок к Anarhichas orientalis, а по строению зубной системы — к Anarhichas latifrons. Он проник в Атлантический океан, вероятно, не позднее нижнего плиоцена — насколько об этом можно судить по находке клыкообразных зубов в Coralline Crag Англии (Newton, 1891 : 91—92, pl. X, fig. 4), и, следовательно, ранее предпредпоследней регрессии, предшествовавшей наступлению четвертичного периода (Линдберг, 1953 : 53). Эти зубы, судя по довольно слабой складчатости их оснований, могли принадлежать предку атлантических зубаток с примитивным строением зубов, но не современному виду Anarhichas lupus, которому их приписывает Ньютон (см. табл. VI). Известны также части черепов из London Clay, Sheppey, имеющие некоторое сходство с таковыми Anarhichas (Woodward, 1901 : 556); однако последнее Вудворт ставит под сомнение. Зуб из плиоцена Бельгии, якобы принадлежащий ископаемому роду Palanarhichas crassus (Le Hon H., Prelim. Mem. Poiss. Tert. Belg., 1871 : 10 — цит. по: Woodward, 1901 : 557), не описан. Нахождение зубов именно в Coralline Crag указывает, между прочим, на теплолюбивость предка современных атлантических зубаток, так как климат в это время был теплым и лишь позднее (Red Crag) произошло похолодание (Берг, 1934 : 69—78).
В Атлантическом океане окраска предка изменилась, стала поперечнополосатой, что было связано, возможно, с иным характером прибрежной растительности в местах обитания его молоди по сравнению с Тихим океаном. Постепенное смещение на глубины повлекло за собой обособление сначала Anarhichas latifrons, а затем Anarhichas minor, тогда как прибрежный вид дал современную Anarhichas lupus. Уход на глубины и образование указанных видов происходили, по-видимому, в северных частях ареала в ледниковый период, когда многие прибрежные формы не только рыб, но и других животных вынуждены были уйти на глубины (Шмидт, 1948, гл. VII; Гурьянова, 1938, 1951, и др.), так как прибрежье занималось ледниками на большом протяжении (сильные оледенения, очевидно, совпадали по времени с повсеместными трансгрессиями моря, — Линдберг, 1955 : 285 и сл.). В это же время, вероятно, выработалась способность Anarhichas lupus — прибрежного вида — обитать на больших глубинах по сравнению с тихоокеанскими видами.
Специализация зубной системы шла наиболее быстро у форм прибрежных и более южных вследствие того, что раковины моллюсков и панцири морских ежей на юге и у берегов в общем толще (Sverdrup, 1942 : 853—854), а количество моллюсков в общем больше, чем на севере и на глубинах. В связи с этим наибольшей специализации, как указывалось, достигла Anarhichthys ocellatus, наиболее южная из прибрежных форм, а внутри нее, по-видимому, самые южные калифорнийские особи; за ней следует Anarhichas orientalis — прибрежная форма, распространенная далее к северу. Несколько меньшая специализация достигнута Anarhichas lupus, опускающейся на большие глубины, нежели Anarhichas orientalis; затем слудет Anarhichas minor, редко встречающаяся у берегов, и, наконец, менее всего специализирован зубной аппарат у Anarhichas latifrons, наиболее глубоководной формы. Внутри вида Anarhichas lupus специализация зубной системы зашла дальше у Anarhichas lupus maris-albi, у которой весь нагульный период протекает у берегов, на небольших глубинах.
Чем больше была степень специализации к питанию организмами с прочным панцирем, тем меньше степень приближения к торпедообразной форме тела, так как при типе строения, характерном для зубаток, слабая специализация зубного аппарата предоставляет большие возможности для приобретения обтекаемой формы тела, чем сильная. Переход к обтекаемой форме тела был связан, вероятно, с выработкой большей скорости движения. Крупные размеры тела зубаток, возможно, облегчали эту выработку, так как поверхность трения по отношению к объему тела у крупных рыб меньше, чем у мелких. Это особенно важно на севере и на глубинах, где низкие температуры сильно повышают вязкость воды (Книпович, 1939 : 310), а последняя, очевидно, затрудняет выработку большей скорости движения у мелких рыб сильнее, чем у крупных. У зубаток же наиболее обтекаемую форму тела приобрели именно северные, а из последних — глубоководные формы. По-видимому, это было связано с увеличением протяженности миграций, а у глубоководных форм также с уменьшением числа убежищ на дне и с большей ролью нектонных организмов в питании.
Сильная специализация зубного аппарата ранее всего была достигнута, очевидно, Anarhichtys ocellatus, еще при угреобразной форме тела, сохранившейся и до сих пор. Позднее, уже после ухода части особей в Атлантический океан, произошла специализация у Anarhichas orientalis, еще позднее — у Anarhichas lupus. Вид, ранее ушедший на глубины Атлантического океана, Anarhichas latifrons, сохранил самый примитивный зубной аппарат, но приобрел при этом наиболее обтекаемую форму тела. Более поздний и не столь значительный уход на глубины Anarhichas minor обусловил среднее положение этого вида между Anarhichas latifrons и Anarhichas lupus.
Таким образом, эволюция предков зубаток в сторону специализации к питанию организмами с твердым панцирем, судя по всему, раньше всего началась у берегов, возможно, в южной части ареала. Особи с менее специализированным зубным аппаратом оттеснялись на север и на глубины, где, приспособляясь к новым условиям, вырабатывали большую подвижность за счет приближения формы тела к «торпедообразной». Зубаток, не изменивших своего строения, не сохранилось и сохраниться не могло. Особи с менее специализированным зубным аппаратом, оттеснявшиеся на север и на несколько большие глубины, попадая в прибрежья, еще не заселенные зубатками, претерпевали специализацию зубного аппарата в том же самом направлении, что и их предки, что и привело сначала к образованию Anarhichas orientalis, а затем Anarhichas lupus. Параллелизм развития этих, ныне высокоспециализированных форм и Anarhichthys ocellatus обусловливался как внешними, так и внутренними причинами (наличие организмов с твердым панцирем, слабо использующихся другими видами рыб; тип строения, обусловливающий тенденцию к питанию такими организмами). Говоря словами Чарлза Дарвина (Darvin, 1871 : 194), относящимися к приматам, «... организмы, имеющие общее происхождение и сходную конституцию, подверглись сходным влияниям, вызвавшим сходные изменения». («Many of these resemblances are more probably due to analogous variation, which follows. . . from co-descended organisms having a similar constitution and having been acted on by similar causes including variability».)
Оттеснение на север и на глубины особей со слабо изменяющимся зубным аппаратом первоначально, очевидно, приводило к образованию непрерывной или так называемой «клинальной» изменчивости (Huxley, 1938). Превращение непрерывной изменчивости в прерывистую означало образование видов. Для решения вопроса, как и почему происходило это превращение, необходимо: 1) гораздо более подробное изучение взаимоотношений между элементами внутренней среды (органы, ткани, клетки и т. д.) в их развитии; 2) изучение истории бассейнов и особенно истории обитающих в них биоценозов, членами которых являлись и являются зубатки. В первом случае мы имеем дело с закономерностями, внутренней среды, поставляющими материал для естественного отбора, во втором — с закономерностями внешней среды, определяющими направление этого отбора. Познание указанных закономерностей позволит ответить на вопрос, почему зубатки заселили в Атлантическом океане такие глубины, каких у себя на родине, в Тихом океане, они либо не смогли заселить, либо не смогли удержать. Возможно, изменения формы тела, совершившиеся у зубаток еще в северных частях Тихого океана, облегчили зубаткам, попавшим в Атлантический океан, заселение гораздо больших глубин, на которых может быть, отсутствовали конкуренты, имеющиеся или (и) имевшиеся в Тихом океане.
Переход к «торпедообразной» форме тела, по-видимому, сопровождался оттеснением угреобразной стадии на более ранние этапы онтогенеза, вплоть до ее исчезновения у личинок современных Anarhichas, выходящих из икры. Подобное «продвижение» вновь приобретенных особенностей к началу онтогенеза наблюдается в ходе специализации зубного аппарата; в частности, «пережим» на dentale, являющийся чертой специализации, у Anarhichas lupus lupus заметен уже у вылупляющихся личинок, хотя сами зубы находятся еще в «примитивном» состоянии. Тем не менее возрастные изменения формы тела у Anarhichas на протяжении всей жизни протекают в основном в том же самом направлении, что и у Anarhichthys: от более удлиненной формы тела к менее удлиненной.
Высказанное представление об эволюции и расселении семейства зубаток, очевидно, правдоподобнее всех других, которые можно вывести из имеющихся в настоящее время материалов по морфологии, биологии и распространению этих рыб. Однако некоторые факты трудно объяснимы с изложенной точки зрения. В частности, у молоди атлантических зубаток не обнаруживается никаких следов продольной полосатости, столь характерной для тихоокеанских. Требуются дополнительные гистологические исследования возрастной изменчивости окраски у личинок и мальков всех зубаток, чтобы выяснить, насколько данный факт противоречит высказанной гипотезе. Не исключена возможность, что возрастные изменения окраски проходили у древних атлантических зубаток через распадение продольных полос на пятна, затем через группировку пятен в поперечные полосы и новое распадение последних на пятна; в дальнейшем, в ходе развития, две первых стадии были вытеснены. Однако это предположение пока что почти ничем не обосновано.
Далее, у зубаток, возможно, имеется длительный инкубационный период, приходящийся на холодное время года, что не вяжется с представлением о их происхождении от теплолюбивых форм. Однако и этот период мог быть новоприобретением, при котором «теплолюбивые» стадии развития икры были сильно сокращены и сохранились лишь на определенном этапе ее развития (II и II—III стадии зрелости). К сожалению, размножение Anarhichthys ocellatus совершенно неизвестно.
Наконец, находка остатков представителя семейства зубаток в третичных отложениях юго-западной Африки (Paranarrhichas damesi, — Bohm, 1926 : 84, Texttabel В, Fig. 20, a, b) могла бы указывать на весьма широкое распространение зубаток в Атлантическом океане в это время, если бы имелись доказательства того, что несколько найденных конических зубов принадлежат именно зубатке. Автор указанной выше работы таких доказательств по существу не приводит.
Все перечисленные выше данные, противоречащие приведенной схеме происхождения и расселения зубаток, требуют проверки или дальнейшей разработки, как, впрочем, и многие из подтверждающих ее фактов. Понятно, что считать эту схему достаточно доказанной еще нельзя, но я надеюсь, что высказанные предположения окажутся интересными. Надеюсь, что они подтолкнут исследователей, для которых угревидная зубатка, а также личинки и мальки других видов, по счастью, окажутся более доступными, на подробное исследование прежде всего этих объектов и на критическую проверку высказанной гипотезы.
Вопрос о происхождении исходного вида, давшего начало семейству зубаток, т. е. о его отношении к другим семействам подотряда Blennioidei, мною не разбирался. Зубаток прежде сближали сначала с Zoarcidae (Cuvier et Valenciennes, 1836 : 472; Swainson, 1839 : 283), затем с Blenniidae (Bonaparte, 1846 : 7; Günther, 1861 : 208, и др.). Современные исследования (Макушок, 1958) показали, что семейство зубаток входит в состав надсемейства Stichaeoidae, явно имеющего тихоокеанское происхождение, что является еще одним доводом в пользу тихоокеанского происхождения зубаток. Правда, мое предположение о происхождении зубаток от предка с угреобразной формой тела В. М. Макушок отвергает. Он пишет: «В противоположность мнению В. В. Барсукова, Anarhichthys ocellatus представляется нам наиболее специализированным представителем семейства не только по строению зубного аппарата и черепа, но и по строению чрезвычайно удлиненного тела и слиянию спинного и анального плавников с сильно редуцированным хвостовым плавником». В. М. Макушок, очевидно, полагает, что общий предок зубаток имел форму тела, близкую к Anarhichas, но не к Anarhichthys. Однако он не вступает в дальнейшее обсуждение вопроса, не объясняет, почему в эволюции семейства должна была отсутствовать угреобразная стадия. Термин «специализация» применяется В. М. Макушком, судя по всему, к любому уклонению от предполагаемого исходного типа строения данного органа либо их системы. В более определенном значении этого слова специализация (клетки, ткани, органа, системы органов и т. д.) означает изменение, связанное с преимущественным развитием одной или немногих функций за счет ограничения других. Когда мы говорим о специализации зубного аппарата и тесно связанных с ним структур, то имеем в виду сужение их функций, происходящее в результате сужения круга основных пищевых объектов. Специализация в форме тела и плавников означает меньшую универсальность движений рыбы. Угреобразную форму тела, по-видимому, следует считать специализированной по сравнению с торпедообразной, на основании усложнения плавательной волны за счет ограничения функций парных и хвостового плавников. В таком случае Anarhichthys ocellatus действительно является наиболее специализированным представителем семейства, что, однако, не противоречит моему мнению о наибольшей близости этого вида (по форме тела) к ближайшим предкам семейства. Специализация последних, очевидно, еще не зашла так далеко, чтобы стать полностью необратимой. Приобретение предками зубаток угреобразной формы тела не привело еще к редукции грудных и утрате хвостового плавников, но было достаточным для утраты брюшных плавников, слияния непарных плавников, редукции чешуи (у большинства Stichaeoidae чешуя налегающая, — Макушок, 1958 : 14) и т. д. (см. выше). Следует отметить, что степень редукции осевого скелета хвостового плавника у Anarhichthys ocellatus нам неизвестна; не исключена возможность, что в данном отношении Anarhichthys ocellatus пошел дальше, чем его предки, однако этот вопрос остается открытым. У Anarhichas число свободных хвостовых позвонков, т. е. задних позвонков (с уростилем), между отростками которых не внедряются опорные элементы спинного и анального плавников, значительно уменьшено (2—4) по сравнению с большинством Stichaeoidae (4—6 и более). Уменьшение числа этих позвонков обычно сопутствует переходу к угреобразному движению (Макушок, 1958 : 38). Увеличение числа основных лучей хвостового плавника у Anarhichas, слияние эпаксиальных hypuralia и срастание их с уростилем В. М. Макушок считает вторичным явлением. Надо полагать, правильнее было бы считать таковым же и приближение степени слияния перепонок непарных плавников к типу, наиболее обычному для Stichaeoidae. Все эти изменения могли быть обусловлены вторичным приближением к «торпедообразной» форме тела.
Выше уже указывалось, что колючки спинного плавника у Anarhichas постепенно утолщаются к заднему концу плавника, теряя гибкость, тогда как у Anarhichthys все они гибкие. В. М. Макушок установил, что среди Stichaeoidae не наблюдается переходов от форм, несущих только жесткие колючки, к формам, у которых колючки утолщаются к заднему концу плавника, а переходы к последним от форм, имеющих лишь гибкие колючки, существуют (Alectriinae). У личинок Anarhichas lupus lupus различия в толщине колючек еще не выражены и все колючки гибкие. Казалось бы поэтому, что строение спинного плавника у Anarhichthys ближе к исходному типу, однако Макушок (1958 : 32, 53) придерживается противоположного мнения. Изучение онтогенеза спинного плавника у Lumpenus показало, что данный плавник является гомологом лишь переднего, колючего спинного плавника, появившегося в онтогенезе позднее заднего, мягкого; считается, что Anarhichas в этом отношении сходен с Lumpenus (Емельянов, 1948 : 315—321). Однако это еще ничего не говорит о путях перехода от твердых колючек к гибким и о вариациях в их взаимном расположении. В связи с этим изучение онтогенеза плавников у Anarhichas и Anarhichthys, не говоря уже о других Stichaeoidae, приобретает большой интерес.
Следует также остановиться на различии Anarhichas и Anarhichthys в строении мягких лучей анального плавника, отмеченных В. М. Макушком (1958 : 113) и введенным им в диагнозы родов: эти лучи у Anarhichthys состоят из небольшого количества (не более 5) длинных члеников, тогда как у Anarhichas — из очень большого количества коротких члеников; самые задние лучи у Anarhichthys не ветвятся, у Anarhichas — ветвятся. У Anarhichas lupus lupus длиной 45 см и Anarhichas minor длиной 54 см, просмотренных мною, лучи анального плавника действительно состоят из нескольких десятков члеников, диаметр которых значительно превышает длину. Вблизи основания луча границы между члениками незаметны, но по направлению к концу луча четкость границ увеличивается, очевидно, вследствие уменьшения различия между диаметром членика при закладке и его диаметром к моменту вылова рыбы. У Anarhichas lupus lupus длиной 7.5 см и Anarhichas minor длиной 10.5 см лучи анального плавника состоят из 8—11 члеников, длина которых значительно превышает диаметр, и только у последних 2—3 члеников эти измерения становятся примерно равными. Лучи анального плавника у личинки Anarhichas lupus lupus длиной 2.5 см, выходящей из икры, еще не членисты. Следовательно, рост лучей анального плавника у этих двух видов зубаток (а возможно, и у других) происходит вначале в основном за счет удлинения члеников, позднее — в основном за счет их добавления. Так что и в данном случае у Anarhichas, возможно, наблюдаются остатки прежнего типа строения лучей анального плавника, сохраняющегося у Anarhichthys и на более поздних стадиях развития. Экземпляр Anarhichthys ocellatus, имевшийся в нашем с В. М. Макушком распоряжении, является еще довольно молодым, и членистость лучей анального плавника у взрослых особей данного вида, по всей вероятности, не столь сильно отличается от таковой же у рода Anarhichas. Все сказанное выше относительно членистости лучей анального плавника справедливо и в отношении разветвленности последних: у личинки Anarhichas lupus lupus самые задние лучи еще не ветвятся, у Anarhichas lupus lupus длиной 7.5 см и Anarhichas minor длиной 10.5 см их разветвление только намечается.
Дополнение к главе «Географическое распространение, расселение и эволюция». Новые данные по образу жизни молоди свидетельствуют о том, что связь между отдельными частями ареала у каждого из атлантических видов зубаток может осуществляться не только путем миграций взрослых особей, но и путем переноса течениями пелагических мальков. Правда, есть основания предполагать, что эти мальки переносятся обычно на небольшие расстояния, однако в отдельных случаях течения могут уносить их очень далеко от мест выклева.
Наблюдения над окраской пелагических мальков атлантических зубаток позволили обнаружить у них следы продольных полос, отмеченных у молоди тихоокеанских зубаток на более поздних стадиях развития. Оказалось, что поперечная полосатость у Anarhichas lupus lupus появляется позднее, чем следы продольных полос. У Anarhichas minor поперечная полосатость возникает раньше, чем у Anarhichas lupus lupus: возможно, окраска молоди общего предка этих видов, возникшая как средство подражания слоевищам бурых водорослей, у Anarhichas minor, претерпев лишь незначительные изменения, преобразовалась в расчленяющую окраску, причем сроки ее появления, возможно, сдвинулись на более ранние стадии онтогенеза. Мальки Anarhichas latifrons наименьших размеров, судя по степени слияния непарных плавников и форме хвостового, стоят ближе к тихоокеанским зубаткам, чем мальки Anarhichas minor, а тем более мальки Anarhichas lupus lupus. Все это хорошо согласуется с приведенной выше схемой эволюции зубаток.
Жизнь зубаток, после того, как желточный мешок рассосется, состоит из двух основных этапов, характеризующихся особым типом питания: 1) питание в толще воды организмами без прочного панциря, различной подвижности; 2) донное питание малоподвижными организмами с прочным панцирем. У Anarhichas latifrons первый этап сильно растянут и продолжается почти до начала полового созревания. Предки зубаток, вероятнее всего, отличались сходной растянутостью первого этапа, но проходили его не над большими глубинами на севере, а возле берегов на юге. У Anarhichas minor, а еще более у Anarhichas lupus, первый этап сокращен, что наблюдается, судя по всему, и у тихоокеанских зубаток. Сокращение первого этапа у этих видов происходило, по-видимому, в общем параллельно возрастанию специализации к питанию организмами с прочным панцирем.
БИОЛОГИЯ
Ввиду крайней незначительности сведений о биологии тихоокеанских зубаток данный раздел поневоле посвящен главным образом атлантическим зубаткам; его целью является сравнение видов зубаток для выяснения общих черт в биологии семейства и различий между видами. Биология каждого вида в отдельности подробно изложена в «Систематической части», где в основном и цитируется использованная литература.
Распределение по глубинам. Оба тихоокеанских вида (Anarhichthys ocellatus и Anarhichas orientalis) и один из атлантических (Anarhichas lupus) прибрежные формы; из них Anarhichas lupus lupus встречается на самых больших глубинах (до 435 м), еще глубже распространяется Anarhichas minor (до 550 м), а особенно Anarhichas latifrons (до 936 м). Смещение на большие глубины в ряду Anarhichas lupus — Anarhichas minor — Anarhichas latifrons наблюдается во всех частях их ареалов, где эти виды встречаются вместе (табл. 11).

Предельные глубины установлены в основном по данным тралового лова. Поэтому минимальные глубины, по-видимому, не всюду отражают действительное положение вещей, особенно для Anarhichas lupus lupus. Точно так же и максимальные глубины могут быть неточными, так как трал во время подъема может захватить зубатку, держащуюся не у дна, а в толще воды, что скорее всего можно ожидать для Anarhichas latifrons. Тем не менее наибольшие уловы тралом достаточно достоверно свидетельствуют о том, что всюду Anarhichas lupus lupus предпочитает меньшие глубины, чем Anarhichas minor, а последняя — меньшие, чем Anarhichas latifrons (Барсуков, 1957 : 45). Это подтверждается также ловом на яруса.
Как правило, все зубатки в северных частях ареала летом держатся на меньших глубинах, чем зимой, а в южных частях, возможно, на больших. Сроки выхода на наименьшие глубины в одних и тех же местах у разных видов различны, как и у одного и того же вида в разных частях ареала. Одной из важных причин этих различий является, по-видимому, отношение зубаток к температуре.
Различия в распределении по глубинам между молодью Anarhichas lupus lupus, Anarhichas minor и Anarhichas latifrons выражены резче, чем между взрослыми особями этих видов.
В Баренцевом море все три вида на северных окраинах Западного желоба придерживаются больших глубин, чем в южной части моря, и еще больших — близ материкового свала, в прилегающих к Баренцеву морю частях Норвежского и Гренландского морей.
Судя по максимальным траловым уловам в Баренцевом море, плотность поселения зубаток со смещением на глубины снижается. Anarhichas lupus, по всей вероятности, образует наибольшие скопления, но вследствие того, что они слабо охватываются промыслом, средние уловы ее очень низки. Слабый охват этих скоплений объясняется тем, что Anarhichas lupus предпочитает каменистый грунт, неудобный для траления. Поэтому уловы Anarhichas minor значительно превышают уловы Anarhichas lupus lupus, хотя Anarhichas minor держится, по-видимому, более разреженно. Anarhichas latifrons никогда не образует таких скоплений, как Anarhichas minor. Скопления всех зубаток, возможно, никогда не принимают характера стай.
Отношение к свету. По наблюдениям ряда авторов и опросным данным, прибрежные формы зубаток (Anarhichas lupus и Anarhichas orientalis, относительно Anarhichthys ocellatus сведений нет) избегают яркого освещения, активны у берегов и в аквариуме только ночью, а днем отстаиваются в расселинах скал и других укрытиях.
Отношение к давлению. Все атлантические зубатки могут выдерживать значительные и довольно резкие колебания давления, так как извлеченные тралом с больших глубин (300—400 м) долго остаются живыми на палубе. Эта способность связана, в частности, с отсутствием у них плавательного пузыря.
Отношение к грунту. Все прибрежные виды (Anarhichas lupus, Anarhichas orientalis и Anarhichthys ocellatus) предпочитают каменистые грунты. Anarhichas minor обычно ловится на илистых и илисто-песчаных грунтах, но очень часто при этом трал поднимает со дна и камни. Anarhichas latifrons предпочитает илистые грунты. Таким образом, со смещением на глубины привязанность зубаток к твердым грунтам уменьшается. Однако все зубатки нерестятся, надо полагать, на твердых грунтах.
Отношение к температуре. Географическое распространение атлантических зубаток указывает на большую теплолюбивость Anarhichas lupus по сравнению с другими видами, что подтверждается также наблюдениями над зависимостью траловых уловов зубаток от придонной температуры (Барсуков, 1957 : 46). В указанной работе на рис. 12, А не по вине автора допущена грубая ошибка: кривая, обозначенная цифрой 2, в действительности принадлежит Anarhichas latifrons, а цифрой 3 — Anarhichas minor; в остальном обозначения правильны. Ряд искажений текста, допущенных при редактировании, слишком велик для того, чтобы здесь на нем останавливаться. Наиболее холодолюбивым видом является Anarhichas minor. Anarhichas latifrons, очевидно, несколько более теплолюбива; однако неизвестно, каких температур придерживается молодь Anarhichas latifrons, а также и взрослые особи в тех местах, где обитает молодь. Anarhichas lupus и Anarhichas minor в определенный период созревания половых продуктов, по-видимому, придерживаются более высоких температур, чем обычно. Вероятно, то же самое наблюдается и у Anarhichas latifrons.
Отношение к солености. Anarhichas lupus выдерживает меньшие солености, чем Anarhichas minor и Anarhichas latifrons, что особенно проявляется в Белом море, где другие виды зубаток отсутствуют. Судя по тому, что у Мурманского побережья — одного из наиболее опресненных участков Баренцева моря — Anarhichas minor скапливается дальше от берегов, чем Anarhichas lupus lupus, a Anarhichas latifrons — еще дальше (последняя, кроме того, не входит в Кольский и Мотовской заливы, как остальные), можно полагать, что в ряду Anarhichas lupus lupus — Anarhichas minor — Anarhichas latifrons требовательность к солености возрастает. Зубатки — типичные морские рыбы, и даже наименее требовательный в отношении солености вид Anarhichas lupus в эстуариях не встречается.
Зависимость от течений. Течения оказывают очень сильное влияние на жизнь зубаток. Anarhichas lupus lupus в северных частях арела обитает лишь у берегов, омываемых теплыми течениями. Основные концентрации Anarhichas minor и Anarhichas latifrons в Баренцевом море, как, вероятно, и в других частях их арелов, связаны с районами полярного фронта, т. е. местами стыка холодных и теплых течений. Роль этих течений в расселении личинок и мальков атлантических зубаток, по-видимому, меньше, чем у многих других придонных видов рыб, проходящих ранние стадии развития в толще воды. Приливо-отливные течения, вероятно, имеют большое значение при выборе зубатками мест нереста, так как донной икре зубаток, откладываемой в виде одной большой кладки, надо полагать, требуется хорошая аэрация и твердый грунт. Передвижения взрослых зубаток и во время нагула, по-видимому, каким-то образом также связаны с приливо-отливными течениями (Барсуков, 1957 : 42). Зубатки в этом отношении, возможно, не представляют ничего особенного: оказывается, уловы морского окуня тоже зависят от приливо-отливных явлений (Бараненкова, 1958 : 41).
Миграции. Изучены мало. Лундбек предполагает, что зубатки в Баренцевом море, сходно с другими промысловыми рыбами, совершают миграции с запада на восток и обратно, причем далее всего на восток они: продвигаются в сентябре—октябре (Lundbeck, 1932 : 18). С этим предположением следует согласиться, с той оговоркой, что в отношении Anarhichas lupus lupus хорошо известен лишь подход к берегам и отход на глубины; протяженность ее передвижений, как предполагают, мала. Распространение беломорской зубатки и распределение зубаток в Баренцевом море позволяют предполагать, что в ряду взрослых особей Anarhichas lupus maris-albi — Anarhichas lupus lupus — Anarhichas minor — Anarhichas latifrons протяженность миграций, особенно нерестовых, возрастает (Барсуков, 1957 : 47). По всей вероятности, молодь совершает менее значительные миграции, чем взрослые особи.
Враги. Атлантических зубаток, чаще Anarhichas minor и Anarhichas latifrons, находят в желудках полярной акулы (Somniosus microcephalus), что отмечается и в Баренцевом море (Книпович, 1902 : 304; собственные наблюдения), и у побережья Америки (Bigelow a. Schroeder, 1948 : 520). В желудках трески изредка встречается молодь Anarhichas lupus lupus, еще реже — молодь Anarhichas minor и Anarhichas latifrons; молодь Anarhichas minor, кроме того, однажды была найдена в кишечниках сайды.
Паразиты. У зубаток Баренцева и Белого морей Ю. И. Полянским, С. С. Шульманом и Р. Е. Шульман-Альбовой найдено 34 вида паразитов: 15 у Anarhichas latifrons, 12 у Anarhichas minor, 19 у Anarhichas lupus lupus и 21 у Anarhichas lupus maris-albi. 5 видов паразитов — Neophasis lageniformis, Steringophorus furciger, Diphterostomum microacetabulum, Lepidophillum steenstrupii, Clavellodes rugosa — поражают все 3 вида баренцевоморских зубаток; все эти виды паразитов, кроме Clavellodes rugosa, найдены и у беломорской Anarhichas lupus maris-albi. 2 вида паразитов обнаружены лишь у Anarhichas latifrons, 15 — только у Anarhichas lupus (из них 5 только у Anarhichas lupus lupus и 8 лишь у Anarhichas lupus maris-albi), у Anarhichas minor не оказалось паразитов, не обнаруженных у других видов зубаток. Интересно, что 2 вида паразитов — Myxoproteus elongatus и Lecithaster gibbosus — найдены у Anarhichas latifrons и у Anarhichas lupus maris-albi, но не обнаружены у Anarhichas minor и Anarhichas lupus lupus. Из других районов Атлантики исследованы нью-фаундленские Anarhichas latifrons, Anarhichas minor и Anarhichas lupus lupus на содержание нематод в мышцах: нематоды обнаружены не были (Templeman, Squires a. Freming, 1957 : 867). Работы по паразитофауне тихоокеанских зубаток мне неизвестны.
Питание. Краткие сообщения о питании зубаток, разбросанные в многочисленных работах, содержат в большинстве случаев лишь качественный анализ. Гораздо чаще отмечается сходство отдельных видов в питании, а именно преобладание иглокожих, моллюсков и ракообразных над другими объектами, чем различие. Для уточнения этих различий собственные наблюдения, наблюдения Полярного научно-исследовательского института морского рыбного хозяйства и океанографии (ПИНРО) и литературные данные сведены в табл. 12, примерно отражающую значение важнейших групп пищевых организмов в питании атлантических видов. Данные по питанию Anarhichas minor — большей частью, а по питанию Anarhichas lupus maris-albi целиком получены на основании собственных наблюдений. В питании беломорской зубатки рыба, икра пинагора, трава и водоросли не учитывались, так как эти объекты она в большинстве случаев хватала, уже будучи в ловушке. Наблюдения над тихоокеанскими зубатками единичны и поэтому в таблицу не включепы.
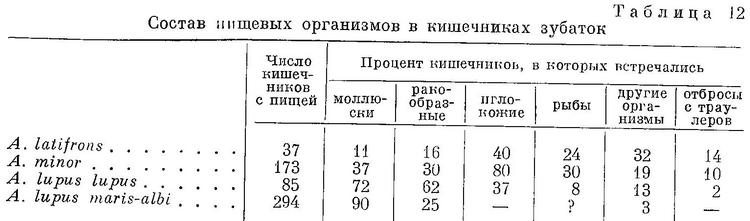
Из табл. 12 видно, что в ряду Anarhichas latifrons — Anarhichas minor — Anarhichas lupus lupus — Anarhichas lupus maris-albi быстро возрастает роль моллюсков в питании, а вплоть до Anarhichas lupus lupus — и роль ракообразных (из которых преобладают раки-отшельники и крабы). Иглокожие играют большую роль в питании Anarhichas latifrons и особенно Anarhichas minor. Роль иглокожих в питании Anarhichas lupus lupus, вероятно, несколько преувеличена, так как предпочитаемые ею каменистые грунты, где иглокожих мало, не охватываются тралением и Anarhichas lupus lupus здесь ускользает от наблюдений. В кишечниках Anarhichas lupus maris-albi, интенсивно питавшейся летом 1951 г. у берегов на каменистом грунте, не было обнаружено ни одного иглокожего (дер. Летняя Речка). В губах Гридинской и Чернореченской этот подвид использует и иглокожих, но, по-видимому, в небольших количествах. В отношении питания рыбой вопрос остается неясным, так как мелкая рыба, обнаруженная в кишечниках зубаток, может быть не поймана ими, а подобрана уже мертвая, выброшенная с траулеров вместе с другими отбросами (головы, кишечники, позвоночники трески и пикши). Какую-то часть рыбы зубатки, безусловно, ловят. Встречаемость в кишечниках отбросов с траулеров снижается в ряду Anarhichas latifrons — Anarhichas minor — Anarhichas lupus lupus, что может косвенно указывать и на снижение питания рыбой в естественных условиях. Правда, в табл. 12 отмечена повышенная встречаемость рыбы в кишечниках Anarhichas minor по сравнению с Anarhichas latifrons. Дело, однако, в том, что наблюдения над питанием пятнистой зубатки были произведены мной в значительной мере в районе большого скопления мойвы; в обычное время эта встречаемость заметно ниже. Разнообразие питания в общем уменьшается в ряду Anarhichas latifrons — Anarhichas minor — Anarhichas lupus lupus — Anarhichas lupus maris-albi.
Рядом авторов замечено, что у моллюсков, которыми питается Anarhichas lupus lupus, раковины в общем толще, чем у тех, которыми питается Anarhichas minor, а тем более Anarhichas latifrons. По-видимому, в ряду Anarhichas latifrons — Anarhichas minor — Anarhichas lupus lupus нарастает способность использования не только моллюсков с более толстой раковиной, но и крабов с более прочным панцирем, морских ежей с более мощным скелетом и т. п. Точных наблюдений над этим еще нет. При их проведении необходимо учитывать возрастную изменчивость зубаток и сравнивать лишь особей более или менее одинакового размера.
Дополнение к главе «Биология», раздел «Питание». Пелагические мальки атлантических зубаток, по данным Т. К. Сысоевой, питаются разнообразными планктонными организмами, пелагической молодью донных и придонных животных, а также икрой и личинками рыб. В питании придонной молоди Anarhichas lupus lupus длиной 7—19 см, по данным И. Я. Пономаренко, моллюски играют довольно значительную роль, но они совсем не отмечены у молоди Anarhichas minor длиной 9—14 см. В кишечниках последней, кроме иглокожих, обычных для обоих видов, встречена рыба и Themisto abyssorum (Boeck): по-видимому, у молоди Anarhichas minor привязанность ко дну меньше, чем у молоди Anarhichas lupus lupus сходных размеров, пищу которой составляют исключительно донные организмы.
Размножение. Исследовано весьма слабо. Время и места нереста точно не установлены, образ жизни личинок и мальков почти совершенно не изучен, а продолжительность инкубационного периода вообще неизвестна. Все это обусловлено в основном, по-видимому, привязанностью нереста зубаток к местам, мало доступным для траления. Большие затруднения доставляет отсутствие надежного определителя кладок икры, личинок и мальков наименьших размеров. До недавнего времени указанные затруднения усугублялись отсутствием шкалы для определения стадий зрелости зубаток и полным незнанием продолжительности этих стадий.
Шкала для определения стадий зрелости самок Anarhichas lupus maris-albi была разработана автором ранее (Барсуков, 1953 : 1212); с небольшими изменениями, основанными на немногочисленных наблюдениях над другими видами зубаток, она приводится ниже (табл. 13). Эта шкала может применяться для определения стадий зрелости самок всех зубаток: различия между видами в диаметре икринок, в их окраске, в отношении веса гонад к весу тела, естественно, наблюдаются и будут еще уточняться, но, надо полагать, не окажутся столь значительными, чтобы сделать шкалу непригодной для какого-либо из видов; абсолютно ничего неизвестно лишь в отношении Anarhichthys ocellatus. В дальнейшем стадии зрелости самок будут приводиться в тексте по данной шкале, а исключения — оговариваться.

Развитие икринок у самки III стадии идет очень равномерно: колебания размеров икринок, взятых из разных частей яичника, поразительно малы (при самых крупных икринках не превышают 0.2—0.3 мм), окраска одинакова. Часть икринок, обычно очень небольшая, гибнет, не достигая IV стадии: гибнут, вероятно, отстающие в росте. Их остатки рассасываются медленно, часто сохраняясь вплоть до VI стадии. Переход икринок из III в IV стадию, по-видимому, происходит очень быстро, сопровождаясь рядом изменений, подробно описанных Фултоном и Милроем (Fulton, 1898 : 122; Milroy, 1898 : 149) не только для Anarhichas lupus lupus, но и для некоторых других рыб. На IV стадии зрелости диаметр всех икринок в яичниках одной и той же самки остается почти одинаковым. При переходе из III стадии в IV увеличенные икринки располагаются отнюдь не всегда в задней части яичника: иногда вдоль спинной стороны его, иногда даже в передней части. О порционности нереста у зубаток не может быть и речи. Мелкие белые икринки, находящиеся в яичниках при всех стадиях зрелости, включая III и IV (кроме I), являются генерацией для нереста следующего года (или ряда лет).
Длительность стадий зрелости можно наметить лишь приблизительно. I и II стадии длятся несколько лет каждая (последняя по крайней мере до первого нереста), III стадия — несколько месяцев, III—IV и IV стадии несколько дней, IV стадия — по-видимому, от одного до нескольких месяцев. Единичные крупные икринки, оставшиеся после нереста, возможно, иногда могут сохраняться в яичниках вплоть до начала развития икринок для следующего нереста. Так, Фултон (Fulton, 1891а : 254) описывает яичники Anarhichas lupus lupus, в которых 28 V 1880 среди мелких икринок (0.8—0.9 мм) и более крупных (2.1—2.2 мм) находилась одна очень крупная икринка (6.1 мм). Очевидно, она осталась от предыдущего нереста, поскольку предположить переход из III стадии в IV при столь малом диаметре икринок (2.1—2.2 мм) трудно. У Anarhichas lupus maris-albi этот переход происходит при диаметре икринок 3.5—5.5 мм.
Поскольку II стадия у впервые созревающих самок зубаток длится более одного года, размеры и возраст созревания должны определяться лишь непосредственно перед нерестом, во время нереста или сразу после него, когда все зрелые самки, нерестящиеся в данном году, находятся в III, IV или VI стадиях зрелости, но не во II стадии.
Не встречено ни одного самца зубаток с отношением веса семенников к весу тела большим, чем 0.27%. В среднем вес семенников у исследованных мной самцов Anarhichas minor и Anarhichas latifrons составлял около 0.10% веса тела, у незрелых самцов Anarhichas lupus maris-albi — также около 0.10%, у зрелых — около 0.16%. Весьма малые размеры семенников, вероятно, связаны с кучным единовременным откладыванием икры самками, облегчающим оплодотворение (McIntosh a. Masterman, 1897 : 202), а может быть, и с какими-либо другими его особенностями. Фултон (Fulton, 1892 : 240) считает, что у рыб с демерсальной икрою разница в размерах между яичниками и семенниками обычно бывает меньше, чем у рыб с пелагической икрою; зубатки представляют собой весьма резкое исключение из этого правила.
Дополнение к разделу «Стадии зрелости». В марте 1959 г. на банке Копытова обнаружен самец Anarhichas latifrons с весом семенников 0.54% веса тела — для самцов зубаток это наиболее высокий индекс зрелости из наблюдавшихся до сих пор. Текучих самцов зубаток, как и текучих самок, никто еще не наблюдал. У Anarhichas latifrons в яичниках самок III стадии икринки переходных размеров перестают встречаться при гораздо большем диаметре крупных икринок (около 4.5 мм), чем у Anarhichas lupus maris-albi (2.2 мм); окраска крупных икринок лимонно-желтая, кремовая или оранжеватая.
Созревание. Половой состав. Прежде чем созреть, самка, по-видимому, должна достигнуть определенной степени упитанности: у самок III стадии вес печени по отношению к весу тела больше, чем у самок II стадии и самцов, пойманных в то же время (известно для Anarhichas minor и Anarhichas lupus; обычно больше и вес тела (без веса гонад) при той же длине (Anarhichas lupus maris-albi).
Есть основания предполагать, что в определенный период развития гонад зубатки должны обитать при более высоких температурах, чем обычно. У самок Anarhichas lupus maris-albi это прежде всего период перехода из II в III стадию, а возможно, и значительная часть III стадии, а также II стадия в то время, когда нерестующие в данном году особи проходят III стадию.
В ряду Anarhichas lupus maris-albi — Anarhichas lupus lupus — Anarhichas minor — Anarhichas latifrons размеры, при которых впервые созревают самки, увеличиваются. Самки Anarhichas lupus lupus в Северном море созревают при больших размерах, чем в Баренцевом (табл. 14).

Темп роста молоди повышается в том же ряду (см. ниже).
Самцы зубаток, по-видимому, созревают при несколько меньших размерах, чем самки. Однако средние размеры у самцов немного больше, чем у самок. Самцы обычно преобладают над самками в количестве.
Плодовитость самок вследствие крупного размера икринок невелика. Под плодовитостью самок понимается количество икринок, выметываемых в данном году. Поэтому плодовитость зубаток определялась мной, как и Сахно (Маслов, 1944 : 156), только у самок III и IV стадий зрелости и подсчитывались лишь крупные икринки; мелкие белые икринки, по числу примерно равные крупным и представляющие собой генерацию для следующего нереста, не подсчитывались. В ряду Anarhichas lupus maris-albi — Anarhichas lupus lupus — Anarhichas minor — Anarhichas latifrons абсолютная плодовитость возрастает соответственно увеличению размеров зрелых самок вплоть до Anarhichas minor, но у Anarhichas latifrons вновь снижается (табл. 15).

Относительная плодовитость в указанном ряду, по-видимому, уменьшается.
Восстанавливаемость стада зубаток, очевидно, обеспечивается не столько большим количеством икринок, личинок и мальков, сколько их повышенной выживаемостью. В силу большой величины икринок личинки выходят крупными и сформированными (Расс, 1935 : 599). Исключительная равномерность развития икринок обеспечивает большой процент выхода личинок (Морозов, 1951 : 464). В связи с этим интересна отмеченная выше гибель отстающих в росте икринок у самок III стадии.
Икра, личинки и мальки. Размеры зрелых икринок у Anarhichas lupus обычно колеблются от 5.5 до 6.5 мм; на Баренцевом море — до 7 мм. Диаметр зрелых икринок у Anarhichas minor (по Рассу, 1941 : 31) сходен с диаметром их у Anarhichas lupus, a у Anarhichas latifrons заметно больше (7.25—8.0 мм).
Велики ли видовые различия в характере кладок, в строении и образе жизни личинок и мальков, сказать очень трудно, поскольку надежного определителя кладок икры, личинок и мальков не существует, к тому же и находки их редки. О видовой принадлежности кладки пока можно говорить лишь в том случае, если она найдена в районе, где нерест других видов исключен или мало вероятен. Личинок и мальков можно определять по числу лучей в плавниках, однако этот метод определения до сих пор еще не применялся.
У Anarhichas lupus lupus кладка имеет форму правильного шара значительных размеров, в котором икринки очень крепко склеиваются между собой частями оболочки. Между ними остаются промежутки, необходимые для аэрации. Надо думать, икринки, находящиеся в центре кладки, очень плохо аэрировались бы, если бы в промежутках между ними не наблюдалось постоянного движения воды. Следовательно, кладка должна выметываться там, где имеются постоянные и довольно сильные придонные течения. Такие течения всегда существуют в местах с каменистым грунтом, где Anarhichas lupus lupus и нерестится. Судя по всему, кладки Anarhichas minor и Anarhichas latifrons похожи на кладки Anarhichas lupus и откладываются тоже на каменистый грунт. По устному сообщению А. С. Бараненковой, кладка какой-то зубатки была найдена внутри большой листовидной губки.
Охраняется ли отложенная икра, достоверно неизвестно. Исчезновение из уловов самцов Anarhichas lupus maris-albi сразу после нереста позволяет предполагать такую охрану, по крайней мере у Anarhichas lupus maris-albi.
Личинки Anarhichas lupus lupus, вылупившиеся из икры, пробывшей несколько дней в аквариуме, оказались очень мелкими (11 —12 мм) и слабо сформированными. Вылупление на такой ранней стадии в море вряд ли является нормальным, но все же величина и сформированность личинок, вылупившихся из различных кладок, заметно варьирует, что может быть вызвано колебаниями гидрологических условий, различиями в размере икринок и т. д. Личинки из одной и той же кладки, напротив, почти не различаются ни по величине, ни по сформированности. Максимальные размеры вылупляющихся личинок Anarhichas lupus lupus в Баренцевом море 25 мм. Минимальные размеры уже вылупившихся личинок, добытых вне связи с кладкой, в Баренцевом море 19 мм, у берегов Шотландии и Исландии 17—18 мм, у берегов Северной Америки 21 мм.
Личинки и мальки зубаток длиной 17—44 мм ловятся как у дна, так и у самой поверхности моря. Судя по всему, эти личинки и мальки принадлежат исключительно Anarhichas lupus lupus. Молодь такого размера у Anarhichas minor и Anarhichas latifrons пока, по существу, неизвестна. Более того, молодь Anarhichas latifrons длиной менее 6 см никем не указана, а менее 60—65 мм не описана. Мальки Anarhichas lupus lupus длиной более 44 мм ловятся только у дна. Anarhichas orientalis по образу жизни личинок и мальков, возможно, напоминает Anarhichas lupus lupus. У Anarhichthys ocellatus молодь, по-видимому, пелагическая.
Дополнение к разделу «Икра и личинки». Диаметр зрелых икринок, оставшихся в яичниках после нереста, у самок Anarhichas minor длиной 65, 107 и 110 см равен 6.1, 6.3 и 6.7 мм, у самок Anarhichas latifrons длиной 108 и 119 см — 6.0 и 6.5 мм (март 1959 г.). По-видимому, диаметр зрелых икринок у всех атлантических видов зубаток в общем сходен, у самок же одинаковой длины уменьшается в ряду Anarhichas lupus — Anarhichas minor — Anarhichas latifrons.
Личинки зубаток, вылупившиеся из икры, по данным А. С. Бараненковой и Н. С. Хохлиной, вначале, по-видимому, держатся у дна и лишь по истечении некоторого времени начинают подниматься в толщу воды. Наевшись здесь, мальки, возможно, опускаются на дно (если позволяют глубины), где переваривают пищу в состоянии пониженной активности — судя по аквариальным наблюдениям А. С. Бараненковой за мальком Anarhichas lupus lupus длиной 24 мм (отчет за 47-й рейс э/с «Персей-2»). Глубины, над которыми вылавливаются мальки, возрастают в ряду Anarhichas lupus lupus — Anarhichas minor — Anarhichas latifrons: 72—290 (большинство мальков), 265—455 и 470 — > 1000 м, соответственно. Мальки Anarhichas lupus lupus довольно часто поднимаются к самой поверхности, что у мальков других атлантических видов, очевидно, наблюдается реже. Минимальные размеры мальков, добытых в толще воды, у баренцевоморской Anarhichas lupus lupus равны 19 мм, у Anarhichas minor — 24 мм, у Anarhichas latifrons — 25 мм; максимальные — 47 мм (37 экз.), и 60 мм (13 экз.) и 26 мм (2 экз.), соответственно. Постепенность перехода от пелагического питания к придонному возрастает в ряду Anarhichas lupus — Anarhichas minor — Anarhichas latifrons, особенно резко у Anarhichas latifrons: даже у очень крупных особей этого вида питание в толще воды, судя по ничтожной стертости старых зубов, иногда преобладает над донным питанием.
Сроки нереста и инкубационный период. Сроки нереста достаточно точно определены лишь для Anarhichas lupus maris-albi, которая отнерестилась в 1951 г. в основном в июле и первой половине августа. Прибрежный тихоокеанский вид Anarhichas orientalis нерестится, по-видимому, также летом. Anarhichas lupus lupus в Северном море и у побережья Исландии нерестится с июля—августа по февраль. Сроки нереста, таким образом, удлинены к зиме по сравнению с беломорским подвидом, что может быть связано с нерестом части особей Anarhichas lupus lupus на глубинах, обогреваемых теплыми течениями. Anarhichas minor в Баренцевом море и у побережья Исландии, по некоторым данным, нерестится летом. У Anarhichas latifrons также намечается летний нерест, но, возможно, сроки его очень растянуты. Известно, что температура сильно влияет на развитие гонад и нерест у рыб. Учитывая большую сложность изменений температуры на глубинах обитания Anarhichas lupus lupus, Anarhichas minor и Anarhichas latifrons в связи с течениями, рельефом дна и т. д., а также различную способность указанных видов к миграциям, имеющиеся сведения нельзя считать достаточными для точного определения сроков нереста. Материалы для этого малы, особенно в отношении Anarhichas latifrons. К тому же, если ряд наблюдений делает возможным предположение о потребности зубаток в более высоких температурах на определенном этапе развития гонад, то ничего не известно о температурах, при которых происходит сам нерест
Судя по тому, что кладки зубаток с развивающимися или вылупляющимися эмбрионами гораздо чаще находят от конца зимы до конца весны, чем летом или осенью, можно полагать, что инкубационный период длится несколько месяцев. Молодь размером от 17 до 44 мм в большинстве случаев ловится весной, что подтверждает высказанное предположение. Однако имеются и противоречащие ему факты. Так, одна из кладок зубатки, с развивающимися эмбрионами, была обнаружена 23 VII 1901 (Collett, 1905 : 113—115), а одна из личинок какого-то вида зубаток, вероятно Anarhichas lupus lupus, в ноябре (Collett, 1879 : 58).
Места нереста. Anarhichas lupus maris-albi, по-видимому, нерестится недалеко от берегов, так как самки IV стадии зрелости, по всей вероятности весьма непродолжительной, редко, но все же попадаются в ставные ловушки, поставленные у самого берега, на глубине нескольких метров. Кладки икры Anarhichas lupus lupus найдены на глубине 9—11 (Шотландия) и 78—84 м (Мурманское побережье). По всей вероятности, глубины, на которых происходит нерест, нарастают в ряду Anarhichas lupus — Anarhichas minor — Anarhichas latifrons. В Баренцевом море в указанном ряду намечается сдвиг нерестилищ на запад и увеличение в связи с этим нерестовых миграций. Нерестилища Anarhichas lupus lupus находятся, по-видимому, почти во всех местах основных скоплений взрослых особей, от Лофотенских островов и западного склона Медвежинской банки до Восточно-Прибрежного района; нерестилища Anarhichas minor — лишь в северо-западной части моря, Anarhichas latifrons — только на материковом свале (стр. 126, 136, 153, 167—168). Можно предполагать, что в связи с большой протяженностью нерестовых миграций Anarhichas minor и Anarhichas latifrons нерестятся не ежегодно. У Anarhichas lupus maris-albi нерест ежегоден.
Флюктуации установлены (по материалам ПИНРО) у баренцевоморских Anarhichas minor и Anarhichas latifrons. «Волны» поколений «урожайных» лет на кривых Сунда (рис. 24) у пятнистой и синей зубаток располагаются очень сходно. Учитывая различие в темпе роста, можно предполагать, что оба «урожайных» года, сказывающихся на этих кривых, у синей и пятнистой зубаток совпадали. Это означает, что какие-то общие для обоих видов условия определяли успешность их нереста и выживания молоди. В связи с этим интересно западное побережье Шпицбергена, где все виды зубаток имеют в среднем наименьшие размеры по сравнению с другими районами Баренцева моря и прилегающих к нему частей Атлантического океана. Только у Anarhichas lupus столь же малые средние размеры наблюдаются еще в районе Лофотенских островов. Предстоит выяснить, действительно ли два указанных района являются районами скопления молоди, так как снижение средних размеров может быть вызвано также скоплением тугорослых взрослых особей. У баренцевоморской Anarhichas lupus lupus флюктуации, судя по всему, выражены еще сильнее, чем у Anarhichas minor и Anarhichas latifrons, однако закономерных «волн» на кривых Сунда не получается, надо полагать, вследствие малого числа наблюдений. Возможно также, что баренцевоморская популяция Anarhichas lupus lupus делится на ряд обособленных стад, колебания численности которых не совпадают по времени. Колебания численности Anarhichas lupus lupus в Северном море, по-видимому, также велики (Lundbeck, 1936 : 102); велики они и в Белом море — у Anarhichas lupus maris-albi, судя по рассказам рыбаков.
Возраст и темп роста. Определение возраста зубаток производилось мной, как и Сахно (Маслов. 1944 : 155), по кольцам на чешуе. Отчетливость годовых колец на чешуе в общем снижается в ряду Anarhichas lupus maris-albi — Anarhichas lupus lupus — Anarhichas latifrons, но все же и у Anarhichas latifrons они достаточно хорошо заметны. Определение возраста контролировалось подсчетом числа склеритов в годовых приростах; оно колеблется от 4 до 13, чаще всего от 5 до 7 (у Anarhichas lupus и Anarhichas minor). В некоторых случаях определение возраста по чешуе контролировалось определением его по костям (hypurale, metapterigoid, позвонки). Кольца на костях видны лучше, чем на чешуе, однако сбор и обработка костей требуют большего времени. Чешуя зубаток имеет еще одно преимущество перед костями: четкость колец на ней при хранении зубаток в формалине и спирте не теряется. Шапочкообразная форма чешуи зубаток делает ее неподходящим объектом для обратного расчисления темпа роста. Новый прирост на чешуе баренцевоморской Anarhichas lupus lupus появляется примерно в феврале—мае, у баренцевоморской Anarhichas minor — в апреле—июне. У зубаток Белого и Баренцева морей темп роста увеличивается в ряду Anarhichas lupus maris-albi — Anarhichas lupus lupus — Anarhichas minor — Anarhichas latifrons (рис. 25). В данном ряду в связи с увеличением нагульных миграций, а также в связи со смещением на глубины, где сезонная смена температур менее резка, по всей вероятности, увеличивается продолжительность нагульного периода, что может явиться одной из причин увеличения темпа роста.
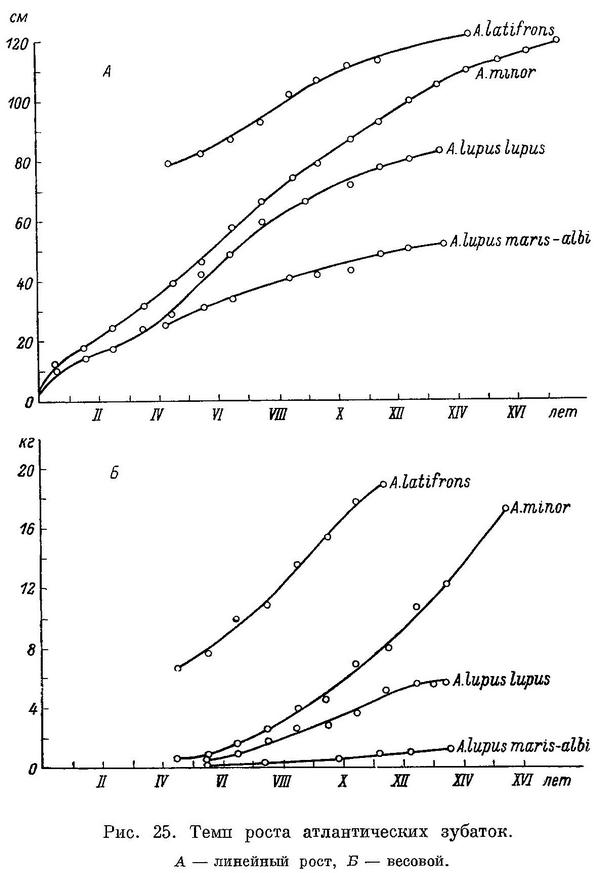
Следует отметить, что методику определения возраста по чешуе и костям зубаток нельзя считать достаточно разработанной и дающей точные результаты. Нужна дальнейшая разработка этой методики и проверка ее путем мечения зубаток. Люман (Lühmann, 1954b : 322) предполагает, что на позвонках Anarhichas minor могут часто появляться дополнительные кольца из-за несовпадения сроков смены зубов и нереста — несовпадения, которого будто бы не наблюдается у Anarhichas lupus lupus. Но в таком случае по темпу роста Anarhichas minor должна была бы превышать Anarhichas lupus lupus еще больше, чем это отмечено, между тем Люман почему-то считает, что оба вида растут одинаково.
Не исключена возможность сильных различий в темпе роста между отдельными поколениями одного и того же вида зубаток в одном и том же районе.
ПРОМЫСЕЛ
Специального промысла зубаток не существует. Большая часть атлантических зубаток вылавливается попутно, при промысле трески, пикши и других донных рыб, в тралы (обычно в количестве нескольких экземпляров за подъем) и на яруса. При ярусном лове трески прилов чаще составляет пятнистая зубатка (Anarhichas minor), при ярусном лове палтуса — синяя (Anarhichas latifrons).
Полосатая зубатка (Anarhichas lupus) в Северном море вылавливается, кроме того, снюрревадами. Она изредка попадает в сети, а также в сельдяные кошельковые невода (Smitt, 1893 : 235). У берегов Северной Америки ловится и на удочку (Bigelow a. Welsh, 1925 : 374). На Белом море добывается вместе с другими рыбами ставными ловушками и очень редко закидными неводами; любители бьют ее острогой у берегов. Подобный промысел существовал также у берегов Гренландии (Fabricius, 1780 : 139) и, по-видимому, у берегов Норвегии (Bloch, 1784 : 22; Donovan, 1802, pl. XXIV, и др.).
Дальневосточная зубатка (Anarhichas orientalis) изредка попадает в ставные и закидные невода, а у берегов Аляски добывается на крючки (Turner, 1886 : 93). Anarhichthys ocellatus довольно часто ловится вдоль тихоокеанского побережья Америки в лососевые ловушки, тралы, сети и случайно на крючки (Clemens a. Wilby, 1946 : 171).
Общий улов зубаток в северной части Атлантического океана в 1906— 1910 гг. составлял около 50 тыс. ц в год. К 1913—1914 гг. в связи с развитием промысла у берегов Исландии улов возрос до 110—120 тыс. ц и в 1920—1924 гг. оставался примерно на том же уровне. В 1925—1938 гг. он достигал 150—200 тыс. ц, а в 1948—1954 гг. — 300—400 тыс. ц (Bull. Statistique, 1906—1956).
В Баренцевом море в 1905—1913 гг. добывалось всего 1—2 тыс. ц зубаток в год. К 1930 г. уловы возросли до 36 тыс. ц. В 1931—1938 гг. здесь вылавливалось 37—57 тыс. ц, из них нашими траулерами 16—39 тыс. ц (табл. 16). У островов Медвежьего и Шпицбергена в это время уловы колебались от 4 до 17 тыс. ц. В Баренцевом море преобладает пятнистая зубатка, составляя 64—84%, а в среднем 78% от общего улова зубаток. Основная масса зубаток вылавливается в западных и центральных районах с февраля по июль. Наибольший процент к общему улову (до 20 и более) наблюдается в феврале, наибольшие уловы на час траления (в среднем около 70 кг) — в марте и апреле. Осенью и в начале зимы, когда ловят большей частью в юго-западных и северо-восточных районах, а также на Канинско-Колгуевском мелководье, уловы на час траления резко снижаются. Все зубатки держатся обычно разреженно. Все же в центральных районах, на Канинской банке, на северном склоне Канинско-Колгуевского мелководья иногда попадает до тонны и несколько более пятнистых зубаток за подъем трала. Зубатки составляют 1.3—5.5% нашего общего годового вылова. В довоенные годы наблюдалась тенденция к снижению этого процента (Маслов, 1944 : 156), что объясняется развитием промысла в северо-западной части моря, где средний размер, а тем более вес пятнистой зубатки меньше, чем в южной, а также в других районах, где зубаток вообще мало. Ярусные уловы зубаток в Баренцевом море значительно уступают траловым: в 1938 г. из общего улова 32 940 ц только 1243 ц было выловлено судами прибрежного промысла.
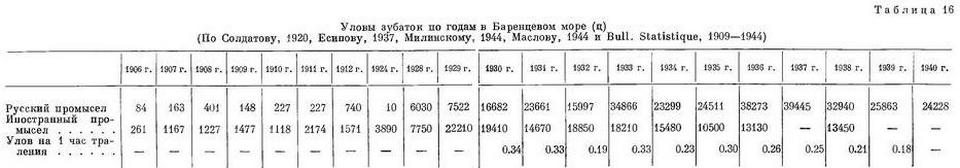
Сведений об общем улове зубаток на Карельском берегу Белого моря нет; во всяком случае, он очень невелик по сравнению с уловами в других морях. Обитающая здесь полосатая зубатка ловится у берегов с мая по сентябрь, причем в течение этого периода уловы постепенно снижаются.
В Северном море в 1904—1922 гг. (исключая 1915—1919 гг.) вылавливалось 20—28 тыс. ц полосатой зубатки в год, а в 1923—1954 гг. (исключая 1939—1946 гг.) — 12—19 тыс. ц. В первом случае это составляло 0.16—0.29% к общему улову всех рыб, во втором — 0.9—0.19%. Уловы зубатки обычно возрастали с января по апрель—июнь, а затем снижались (Thompson, 1905 : 345, fig. 2). В проливах Скагеррак и Каттегат уловы в 1904—1954 гг. колебались от 0.3 до 3 тыс. ц.
Возле Фарерских островов уловы зубаток (в основном полосатой) в 1904—1922 гг. составляли 2—11 тыс.ц в год, в 1923—1927 гг. — 1—8 тыс. ц. В 1928 г. улов достиг 28 тыс. ц. В 1929—1938 гг. добывалось 2—6 тыс. ц, в 1947—1954 гг. — 1—1.5 тыс. ц. Наибольшие уловы зубатки наблюдались здесь с февраля по июнь—июль (Thompson, 1908 : 272—274).
У берегов Норвегии в 1914—1928 гг. добывалось 0.1—0.4 тыс. ц в год, в 1929—1938 гг. — 1—7 тыс. ц, в 1947—1954 гг. — 5—34 тыс. ц.
Больше всего добывается зубаток у берегов Исландии. Преобладает полосатая зубатка, составляющая около 90% общего улова зубаток (Травин, 1952 : 173). Зубатка была важным объектом исландского рыболовства еще в XVIII в., особенно на западном побережье. «В последние тяжелые годы [зубатка] сохранила жизнь многим людям, не только в западных фиордах, но и на южном и западном побережьях — счастье, что ее не покупают иноземцы» — так писал Олафсен (Olafsen, 1774 : 315) в то время. Ныне большая часть вылова зубаток у побережья Исландии приходится на долю иностранных судов. В 1906—1910 гг. здесь вылавливалось 15—18 тыс. ц в год. К 1913 г. уловы возросли до 75 тыс. ц. В 1920—1925 гг. они составляли 71—83 тыс. ц, в 1926—1938 гг. — 93—142, в 1948—1954 гг. — 173—251 тыс. ц. Наблюдались два максимума уловов — в апреле и сентябре (Thompson, 1908 : 272—274).
У берегов Гренландии в 1928—1938 гг. вылавливалось 0.2—1.7 тыс. ц в год. В 1947—1954 гг. уловы неуклонно возрастали и к 1954 г. достигли 28 тыс. ц.
У берегов Северной Америки перед второй мировой войной вылавливалось 11—13 тыс. ц полосатой зубатки в год (Есипов, 1949 : 612).
Мясо пятнистой, полосатой и дальневосточной зубаток обладает высокими вкусовыми и питательными качествами. Употребляется в свежем, копченом, соленом и вяленом виде. Мясо синей зубатки (Anarhichas latifrons) ввиду его студенистости менее ценно. Вкус мяса зубаток, как и других рыб, зависит от того, какими организмами они питались перед тем, как быть выловленными. Более того, исландцы утверждали, что вкус мяса зубатки будто бы портится, если пойманную зубатку положить на траву, поэтому ее клали на берегу только на камни (Olafsen, 1774 : 25).
Икра зубатки очень вкусна, особенно через день-два после посола. Печень зубатки считается у скандинавских рыбаков деликатесом (Smitt, 1893 : 235).
Головой, плавниками и костями исландцы кормили овец и особенно коров, которые давали при этом хорошее молоко (Faber, 1829 : 75). Желчь зубаток употреблялась вместо мыла (Olafsen, 1774 : 314); по Фаберу, она ядовита. На Мурмане приходилось слышать, что мясо зубатки, сваренное или изжаренное в свежем виде, может вызвать легкое отравление. Вероятно, такие случаи наблюдаются при неаккуратной разделке, когда желчь попадает в пищу.
Шкуру зубаток выделывают и изготовляют из нее легкую обувь, сумочки, переплеты для книг, портсигары и т. д. В Гренландии еще в XVIII в. из шкуры зубаток делали сумки для сбора ягод (Fabricius, 1780 : 139). Изделия из этого материала экспортируются оттуда до сих пор. Особенно ценится шкура пятнистой зубатки. В 1939 г. у гренландцев было скуплено 27 тыс. ее шкур (Jensen, 1948 : 121). Эскимосы на Аляске закладывают шкуры дальневосточной зубатки между швов непромокаемой одежды и обуви: при намокании шкура набухает и туго стягивает швы (Turner, 1886 : 94).
При заготовке на промысловых судах зубатки разделываются на пласт с продольным разрубом головы. Методы обработки шкур зубаток даны в брошюре Гельман (1933 : 28—30).
Запасы зубаток, возможно, недостаточно используются промыслом. Преобладающие и максимальные размеры зубаток в уловах на Баренцевом море в настоящее время остаются по существу такими же, какими они были в начале этого века, во времена Экспедиции для научно-промысловых исследований у берегов Мурмана (ЭНПИМ), хотя траловый промысел с тех пор неизмеримо вырос. В Белом море большую часть уловов составляют зрелые особи полосатой зубатки, нерестующие в данном году. Самые старые из них достигают не меньшего возраста, чем в Баренцевом море.
Отсутствие стайности у зубаток делает сомнительными перспективы значительного увеличения их уловов с помощью современных орудий лова. Необходима разработка специальных орудий лова на основе детального изучения биологии зубаток. Это особенно относится к полосатой зубатке, которая в Баренцевом море, по-видимому, гораздо менее затрагивается промыслом, чем пятнистая. Нужна разработка средств привлечения рыбы в ловушку, притом со значительным радиусом действия.
Из органов чувств перспективным в этом отношении может оказаться не только слух, но и обоняние: по наблюдениям Книповича (1902 : 378—379), в одних и тех же местах на яруса попадает гораздо больше пятнистой и синей зубаток, чем в трал, особенно, если на крючки насажена порезанная на куски рыба.
Дополнение к главе «Промысел». В 1949—1958 гг. наши суда вылавливали в Баренцевом море 43—73 тыс. ц зубаток, а вместе с иностранными траулерами 61—109 тыс. ц (табл. 28).
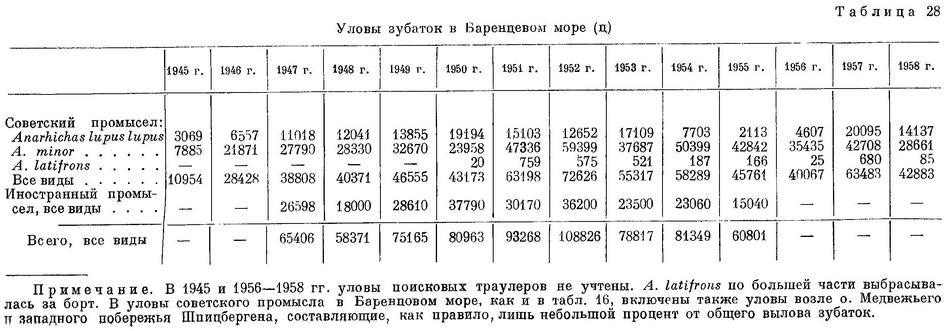
Мурманский рыбокомбинат заготовляет шкуры лишь тех зубаток, которые идут на холодное копчение. В капиталистических странах спрос на шкуры зубаток в последнее время резко снизился; напротив, филе из пятнистой зубатки получило большой спрос на американском рынке, что вызвало резкое увеличение добычи зубаток у берегов Гренландии — с 3520 ц в 1951 г. до 35100 ц в 1956 г. (P. М. Hansen og F. Hermann, Fisken og havet ved Grønland, København, 1953 : 82; P. M. Hansen, Ann. Biol., XIII, 1958 : 53).
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Цель систематической части — дать возможно более полную сводку современных знаний по систематике, распространению и биологии отдельных видов зубаток.
Семейство зубаток представляет собой монолитную группу, отдельные виды которой имеют много общих черт в строении, что, по возможности, отражено в диагнозе семейства. К сожалению, диагнозы родов остались разработанными гораздо слабее из-за недостаточной изученности угревидной зубатки (Anarhichthys ocellatus) и невозможности получения ее для самостоятельного исследования — отчасти это сказалось и на диагнозе семейства.
Роды зубаток так резко обособлены друг от друга, что затруднения в определении их современных представителей возникнуть не могут.
Виды рода Anarhichas определяются уже не столь легко. Их сходство в строении тела велико (особенно у Anarhichas lupus и Anarhichas minor), причем возрастные изменения происходят по большей части параллельно. Тем не менее на каждом этапе виды хорошо различаются по целому комплексу признаков, не имеющих переходов. Не следует лишь забывать, что с возрастом этот комплекс претерпевает изменения, в ходе которых одни из признаков заменяются другими.
Реальность указанных видов не вызывает сомнений. Зубаток, видовая принадлежность которых неясна, мною не встречено. Среди лиц, часто сталкивавшихся с зубатками, также не имеется разногласий по этому поводу. Опытные рыбаки прекрасно различают виды зубаток без всяких определительных таблиц и никогда не дробят ни один из них на несколько видов, чем иногда грешили ихтиологи, работавшие с ограниченным и притом фиксированным материалом. Определительные таблицы рассчитаны главным образом на людей, незнакомых или мало знакомых с зубатками.
Видовые различия, прекрасно улавливаемые на глаз человеком, имевшим дело с несколькими видами зубаток, нередко бывает трудно отразить в точных цифрах и выражениях. В таких случаях чаще всего употребляются промеры длин отдельных частей тела относительно его общей длины или длины других частей. Эти промеры привязываются к хорошим ориентирам — более или менее четким граням на теле рыбы (иначе трудно обеспечить точность, простоту и сравнимость промеров, производящихся разными лицами). И в подавляющем большинстве случаев при подобного рода промерах видовые различия в форме тех или иных частей тела не выделяются, а наоборот, смазываются. Например, Anarhichas minor хорошо отличается от Anarhichas lupus тем, что сошниковый ряд зубов у первой не простирается назад далее нёбных рядов, как это наблюдается у последней (табл. I, 2, 3). Тем не менее встречаются отдельные особи Anarhichas minor, у которых сошниковый ряд несколько выдается назад за нёбные, но так незначительно по сравнению с Anarhichas lupus, что имевшему дело с обоими видами спутать их невозможно. Однако в том случае, когда у человека перед глазами лишь один вид, подобное определение — «незначительно выдается» — никак не может его удовлетворить. Выразить же это различие достаточно точными и простыми промерами очень трудно. Так, если мы выразим длину сошникового ряда через длину нёбных рядов, обязательно появятся переходы между видами в силу того, что сошниковый ряд зубов у обоих видов может выдаваться в обратную сторону — вперед — то больше, то меньше, к тому же нёбные ряды по отношению к нему могут образовать то больший, то меньший угол.
Не помогают и просчеты числа зубов. Различий в зубном аппарате, не связанных переходами, между обоими видами немало, как это было видно уже из морфологического описания. Но просчеты, как и промеры, выражают эти различия очень бледно и зачастую искаженно. К примеру, Anarhichas lupus maris-albi отличается от Anarhichas lupus lupus по числу зубов больше, чем Anarhichas lupus lupus от Anarhichas minor: у взрослых особей Anarhichas minor всех зубов в среднем около 96 (86—116), Anarhichas lupus lupus — около 84 (70—94), Anarhichas lupus maris-albi — около 60 (54—72); на нёбных костях у взрослых Anarhichas minor около 23 зубов (15—27), у Anarhichas lupus lupus — около 19 (14—20), у Anarhichas lupus maris-albi — около 11—12 (8—14). Между тем по форме и расположению зубов (табл. I, 2—4) виды различаются безусловно значительно сильнее, чем подвиды, и притом, как отмечено, в ряде признаков не имеют переходов. Да и странно ожидать, что произвольно взятые соотношения длин частей тела, как и просчеты числа сходных частей, могут дать достаточно полное выражение всему объемному многообразию форм указанных частей, а тем более выразить сущность этого многообразия. Морфометрия, будучи разрабатываема вне связи с функцией, как это часто наблюдается, лишается смысла. Что касается определения, то несравненно лучшим пособием, чем промеры и просчеты, являются фотографии, а особенно рисунки самих рыб и отдельных частей их тела.
При первых определениях не следует удовлетворяться одной лишь определительной таблицей. После нее нужно проверить определение по диагнозу вида. При этом нельзя придавать абсолютного значения ни одному из приведенных признаков, отдельно взятому: очень немногие из них постоянны для рыб всех размеров, да и для одноразмерных особей при увеличении числа наблюдений пределы колебаний признака могут расшириться. Определение следует строить на комплексе признаков. Приобретение достаточного навыка позволит в дальнейшем пользоваться для определения одной лишь окраской и общей формой тела. Таким же путем можно определить и фиксированных рыб, но в данном случае Anarhichas lupus и Anarhichas minor легче различать по зубам. Поэтому зубаток следует фиксировать с открытым ртом: у зафиксированных с закрытым раскрыть его, не повреждая, зачастую невозможно.
Археологи и палеонтологи, интересующиеся видовыми различиями в строении черепа современных видов зубаток, найдут их в диагнозах и на рисунках (табл. I—XXII).
Естественно, все диагнозы могут бесконечно дополняться и совершенствоваться. Нам представляется, что по любой части тела, в частности по любой кости, можно определить вид зубаток — все зависит только от труда, затраченного на поиски признаков, общих для особей данного вида, но без переходов отличающих их от особей других видов. Существовать же такие признаки должны в любой части тела, различаясь лишь по степени выраженности.
Сем. ANARHICHADIDAE — ЗУБАТКИ
Anarhichae Pallas, Zoogr. Rosso-Asiat., III, 1811 : 76.
Gobiadae (частью) Richardson, Fauna Boreali-Amer., III, 1836 : 95—96..
Gobioidae (частью) Wilson, Introd. nat. hist. fish., 1838 : 193.
Zoarcidae (частью) Swainson, Nat. hist. fish., amph., rept., II, 1839 : 283.
Blenniidae (частью) Bonaparte, Catal. Metod., III, 1846 : 7, 68—69.
Anarrhichadidae Gill, Canad. Natur. Geol., (2), II, 1865 : 247—252. — Gill, Proc. U. S. Nat. Mus., XXXIX, 1911 : 158. — Regan, Ann. Mag. Nat. Hist., (8), X, 1912 : 267.
Anarhichadidae Jordan a. Evermann, Fish. N. a. M. Amer., III, 1898 : 2343, 2445. — Берг, Система рыб, 1940 : 316. — Макушок, Тр. Зоолог, инст. АН СССР, XXV, 1958 : 3, 6, 13—18, 20—23, 25—28, 30, 32, 34—35, 41, 43, 45—48, 50—52, 54, 56, 58—59, 63, 67, 112—113, 122, рис. 33.
Anarhichadidae + Anarrhichthyidae Jordan, Classif. fish., 1923 : 235.
Зубы челюстей и нёба приспособлены к разрыванию и раздавливанию организмов с твердым панцирем (моллюски, ракообразные, иглокожие). Зубы на сошнике, нёбных костях и в задней части нижнечелюстных конические или бугорковидные, реже клыкообразные (у молоди). Основную нагрузку несут зубы сошника и часть внутренних зубов нижнечелюстных костей. Зубы сошника при закрытом рте располагаются над промежутком между нижнечелюстными костями, прогнутыми в этой области внутрь. Межчелюстные кости и передние части нижнечелюстных вооружены клыковидными зубами. Зубы на всех перечисленных выше костях, как правило, расположены в два продольных ряда. Глоточные зубы мелкие, клыкообразные. Верхнеглоточные зубы расположены на трех площадках. Цоколи почти всех зубов слившиеся. Замещающие зубы развиваются внутри цоколей. Смена зубов одновременная, ежегодная, но у молоди, по-видимому, поочередная.
Череп за глазами, в области прикрепления верхних отделов внутренних порций жевательных мышц, сужен. Все три порции m. adductor mandibulae имеют места прикрепления на неврокраниуме. Frontalia слиты в одну непарную кость. Parietalia разделены посредством supraoccipitale. Opisthotica малы. Sphenotica отодвинуты от глазницы, снабжены мощным гребнем. Нет базисфеноида. Suborbitalia (4—7) образуют жесткое костное полукольцо от praeorbitale к frontale. На парасфеноиде мощный киль для прикрепления мышцы, приводящей нёбную дугу. С крыльями парасфеноида сочленяются frontale, alisphenoideum, sphenoticum и prooticum. Миодом ограничен областью prooticum; basioccipitale не принимает участия в его образовании. Свод миодома, образованный горизонтальными отростками prooticum, костный. Между parietale, pteroticum и epioticum имеется глубокая впадина, ведущая к лабиринту, но отделенная от него тонкой костной перегородкой, которую образует pteroticum. Sacculus и lagena широко отделены от utriculus (известно для Anarhichas lupus lupus). Нет шипов на крышечных костях. Есть два tabularia с каждой стороны. Кроме того, по В. М. Макушку (1958 : 53), имеются парные tabularia parietalia и непарная tabulare supraoccipitale (она названа temporale supraoccipitale вследствие описки), срастающиеся с соответствующими костями. Передняя половина боковой поверхности позвонков вогнутая, задняя — выпуклая. Туловищные позвонки, кроме одного—двух передних, с парапофизами, к которым, начиная с третьего позвонка, причленяются ребра, доходящие до последнего туловищного позвонка. Большая часть сведений о строении осевого скелета заимствована из работы Макушка (1958); из них сведения относительно скелета хвостового плавника относятся лишь к роду Anarhichas. Число свободных хвостовых позвонков взято из приложения II к статье Макушка (1958 : 123); в диагнозе семейства Макушком (1958 : 112) ошибочно указаны несколько иные числа. Парапофизарные перемычки отсутствуют. Поперечные костные мостики, соединяющие парапофизы туловищных позвонков и отделяющие спинную аорту от почек. Epipleuralia начинаются с первого позвонка (первое epipleurale окостеневает поздно) и оканчиваются на туловищных позвонках. Первая гемальная дуга и первый гемальный отросток обычно на одном и том же позвонке. Первая невральная дуга не срастается с телом позвонка. Переднее interneurale внедряется между остистыми отростками первого и второго позвонков. Каждый остистый и каждый гемальный отросток несут обычно не более одной basalia, а последняя — не более одного плавникового луча. Между остистыми отростками 2—3 и гемальными отростками 3—4 последних хвостовых позвонков basalia не внедряются. Одно эпаксиальное (дорсальное) hypurale; срастается с телом уростиля. Гипаксиальное hypurale, прободеяное отверстием хвостовой вены, hypurale предпоследнего позвонка и uroneurale сидят свободно. Hypurale minimale отсутствует. Scapula и coracoideum широко разделены хрящом; radialia 4, не удлиненные, из них 2 средних сидят на хрящевой перемычке. Postcleithrum отсутствует. Posttemporale не вильчатое. Задние концы тазовых костей соединены, передние расходятся, прилегая к внутренней поверхности нижней части cleithrum соответствующей стороны, с которой соединены связками. Все лучи грудного плавника (18—24) сидят на radialia, кроме 1—2 верхних, сидящих на scapula. Основных лучей хвостового плавника (сидящих на уростилярных hypuralia) 14—15. Лучи грудного и хвостового плавников дважды-трижды ветвятся, как и несколько самых задних лучей анального плавника. Остальные лучи последнего не ветвятся; в начале плавника одна недоразвитая колючка. Все лучи спинного плавника представлены колючками — или гибкими на протяжении всего плавника, или утолщающимися и отвердевающими по направлению к заднему краю плавника.
Тело умеренно или сильно удлиненное, сжатое с боков, покрыто очень, мелкими и тонкими, погруженными в кожу чешуйками, не соприкасающимися друг с другом. Голова сверху сжата с боков, снизу широкая, лишена чешуи. Усиков нет. Рыло тупое, рот большой, ротовая щель ограничена межчелюстной и челюстной костями в равной мере. Верхняя челюсть слабо выдвижная; есть хрящевое rostrale. Обонятельные ямки имеют лишь по одному отверстию, расположенному на конце кожной трубочки. Жаберные отверстия широко разделены межжаберным промежутком. Жаберная щель широкая. Кожная перемычка, соединяющая верхние края operculum и supra cleithrum, образует сифон жаберной крышки. Лучей жаберной перепонки (6) 7 (8). Ложножабры есть. Спинной плавник один, тянется от затылка до хвостового плавника или сливается с ним. Анальный плавник примерно вдвое ниже спинного, простирается от ануса почти до хвостового плавника или сливается с последним. Грудные плавники большие, округлые, посажены низко. Брюшных плавников, нет. В каналах боковой линии на голове число пор варьирует (рис. 18).
Каналы боковой линии на теле отсутствуют: сейсмосенсорные точки сидят открыто в виде верхней и средней ветвей. Короткий и широкий пищевод без резкой границы переходит в сильно растяжимый, хорошо обособленный от кишки желудок. Кишечник и анус очень широкие. Имеется 2 бугорковидных рудимента пилорических придатков. Плавательного пузыря нет. Печень большая, с 2 или 3 долями. Сердце небольшое, не более 0.2% веса тела. Перекрест зрительных нервов происходит не под передним мозгом, а на значительном расстоянии впереди него. Семенники даже в период нереста очень малы, не более 0.6% веса тела. Икра крупная, донная. Свойственны ли перечисленные в данном абзаце признаки Anarhichthys ocellatus, предстоит еще выяснить.
Крупные рыбы, до 1.5—2.5 м длины, распространенные в северных частях Атлантического и Тихого океанов. 5 видов, принадлежащих к 2 родам: Anarhichthys и Anarhichas. Различия между родами настолько велики, что их рассматривают как отдельные подсемейства, а иногда даже как семейства. Последнее явно преувеличено; деление же семейства зубаток на подсемейства практически не имеет смысла, поскольку каждое из них включает в себя лишь по одному современному роду, а ископаемые пока достоверно не известны.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
(Указанные в таблице признаки годны для определения особей всех возрастов — с момента выхода из икринки и до достижения максимальных размеров.)
1 (2). Тело сильно удлиненное: наибольшая высота тела не менее 9 раз в его длине. Спинной и анальный плавники слиты с хвостовым. Позвонков, лучей в спинном, а также анальном плавниках более 200. (Известны пока что особи с числом позвонков менее 250, а не более, как это ошибочно отмечено Макушком (1958 : 113); ниже Макушок (1958 : 123) приводит правильные данные.) 1. Anarhichthys.
2 (1). Тело умеренно удлиненное: наибольшая высота тела меньше 9 раз в его длине. Спинной и анальный плавники обособлены от хвостового. Позвонков, лучей в спинном, а также анальном плавниках менее 100. 2. Anarhichas.
В самое последнее время получено сообщение о находке Anarhichas lupus вблизи г. Генуя (октябрь 1958 г., глубина 80 м, Е. Tortonese, Doriana, II, 94, 1958 : 1—4, 1 fig.) поистине поразительное, так как полосатая зубатка до сих пор не встречалась южнее Бискайского залива; даже в проливе Ламанш и Бристольском заливе она очень редка (М. Graham, Sea fisheries, London, 1956 : 45). He менее удивительно включение Anarhichas latifrons в список рыб Аляски, правда, произведенное без указания на место поимки (N. Wilimovsky, Fisher. Bes. Labor., U. S. Fish. a. Wildl. Serv., 1958; цит. по: V. Walters, Copeia, 1, 1959 : 82). Из списка неиспользованной литературы мною получены работы следующих авторов: Buckland 1881; Вørup; Canestrini (о зубатках ничего не пишет); Ditlevsen (Anarhichas lupus близ Фарерских о-вов на каменистом и песчаном грунтах; характер местообитания определяется наличием пищи; на северном побережьи Исландии только в фиордах); Galløe (Исландия; Anarhichas lupus в больших количествах на глубине 37 м, кишечники набиты мидиями); Gemzoe (Anarhichas lupus; Флрерские о-ва, Исландия; найдена в кишечнике белокорого палтуса длиною 185 см; пойман экз. длиною 120 см); Grøndal; Guomundsson; Hørring (Anarhichas lupus близ Фарерских о-вов на каменистом грунте, связана с зарослями ламинарий; питается в основном крупными двустворчатыми моллюсками); Johansen 1903 (Anarhichas lupus, Исландия; обычная длина 30—40 дюймов); Johansen 1906 (исландские рыбаки украшают жилища сушеными головами зубаток; вяленая Anarhichas lupus — национальное блюдо); Lohnberg (Anarhichas lupus, золотистая вариация окраски); Lundbeck 1892, 1893; Mohr (Исландия; Anarhichas lupus предпочитает каменистый грунт; Anarhichas minor повсюду ловится на яруса, используется как наживка на крючки, по вкусу лучше Anarhichas lupus); Oddson; Otterstrøm; Saemundsson. Зубатки упоминаются также в следующих работах: P. М. Hansen, Beretn. vedr. Grønland, 1, 1956 : 53; 1, 1957 : 49—51, figs 1, 2 (Гренландия; ярусные уловы Anarhichas lupus незначительны; длина и вес Anarhichas minor снижаются по направлению к северу); 1, 1958 : 62, 65 (Anarhichas latifrons, большие уловы на яруса на восточном побережье Гренландии; длина и вес Anarhichas minor в уловах — молодь по мере роста продвигается на юг, к местам нереста; добыта в рингтрал пелагическая молодь всех трех видов зубаток); В. Saemundsson, Handb. Seefischer. Nordeur., VII, 4, 1930 : 27—28; J. Schmidt, Skr. udg. Komm. Havunders., 1, 1904 : 18, 25; G. Weitzel, F. Strecker, U. Roester, A. Fretzdorf u. E. Buddecke, Hoppe-Seyler's Zeitschr. f. Physiol. Ghemie, 295, 1953 : 105, Tab. 1—3, Abb 3 (содержание цинка и инсулина в островках Лангерганса и экзокринной ткани поджелудочной железы Anarhichas minor); М. Junker, Arch. f. Fischerei-wissensch., 7, 3, 1956 : 251 (содержание витамина А в филе Anarhichas lupus и Anarhichas minor).
Рыбы северных морей СССР. А.П.Андрияшев 1954 г.
Подотряд Blennioidei (Jugulares, частью)
XXXIII. Сем. ANARHICHADIDAE — ЗУБАТКОВЫЕ
Anarhichadidae (или Anarrhichadidae) Gill, Proc. U. S. Nat. Mus., XXXIX, 1911 : 158. — Regan, Ann. Mag. Nat. Hist., (8), X, 1912 : 267. — Берг, Система рыб, 1940 : 316.
Anarhichadidae + Anarrhichthyidae Jordan, Classif. Fishes, 1923 : 235.
Blennioidei с весьма специализированным зубным вооружением, приспособленным для раздавливания твердой пищи (иглокожие, моллюски, крупные ракообразные). Тело умеренно или сильно удлиненное, покрыто очень мелкой, погруженной в кожу чешуей. Рот большой, верхняя челюсть не выдвижная. В передней части челюстей сидят клыковидные зубы, с боков конические или бугорковидные. На сошнике и нёбных также конические или бугорковидные зубы, расположенные в 2 ряда. Жаберные отверстия широко разделены межжаберным промежутком. Жаберных лучей 7—8. Ложножабры есть. Спинной плавник один, длинный, состоит из гибких нечленистых лучей, последние же его лучи обычно колючие. Анальный плавник также длинный. Брюшных плавников нет. Плавательный пузырь и пилорические придатки отсутствуют. Череп (рис. 110) за глазами сужен.
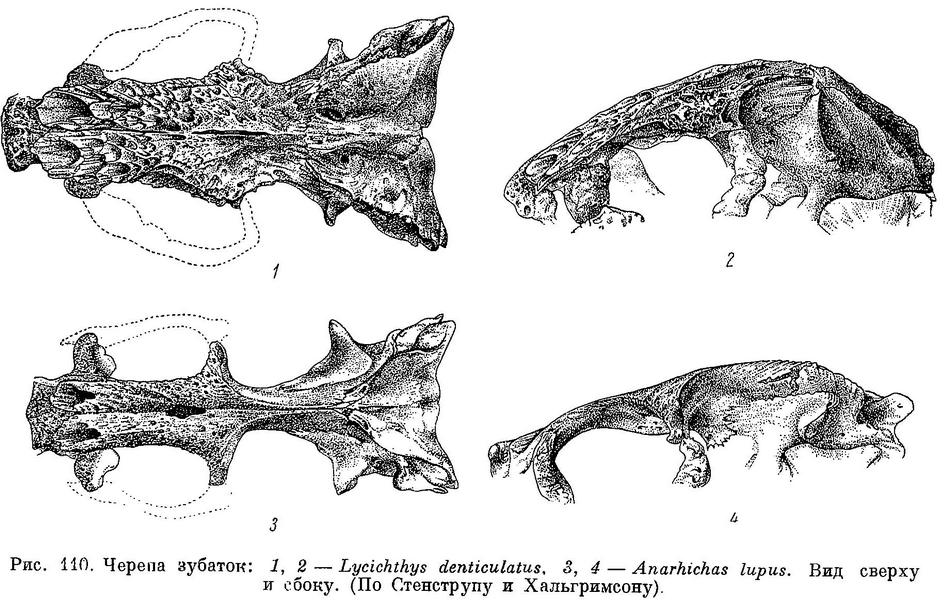
Parietalia разделены посредством supraoccipitale. 6 подглазничных косточек образуют костное полукольцо от lacrimale к frontale. Нет шипов на крышечных костях. Posttemporale не вильчатое. Scapula и coracoideum широко разделены хрящом: radialia 4, не удлиненные. Postcleithrum отсутствует. Берг (1940 : 316) обращает внимание на большие отличия зубаток от других Blennioidei в строении лабиринта, в котором sacculus и lagena широко отделены от utriculus. Икра донная, очень крупная.
Различают 2 подсемейства: Anarhichadinae (Lycichthys Gill., Anarhichas L.; позвонков 74—81) и Anarrhichthyinae (Anarrhichthys Ayres; позвонков около 250).
Крупные рыбы (до 1.5—2.5 м длины), распространенные в северных частях Атлантического и Тихого океанов. В северных морях 2 рода с 4 видами. Определительная таблица для удобства определения дана для видов обоих родов вместе.

ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ LYCICHTHYS И ANARHICHAS
1 (2). Длина сошникового ряда зубов заметно короче нёбного ряда (рис. 111, 1). Длина грудного плавника менее 12% L. Ткани туловища рыхлые, водянистые. Окраска однотонно темная или с неясными крупными пятнами. 1. Lycichthys denticulatus (Krøyer).
2 (1). Длина сошникового ряда зубов равна длине нёбного ряда или значительно превышает ее. Длина грудного плавника более 12% L. Ткани туловища плотные.
3 (4). В спинном плавнике более 80, в анальном более 49 лучей. Сев. часть Тихого океана. 3. Anarhichas orientalis Pall.
4 (3). В спинном плавнике не более 80, в анальном менее 49 лучей. Сев. часть Атлантического океана и сопредельные воды.
5 (6). Сошниковый ряд зубов простирается назад значительно дальше нёбных рядов (рис. 111, 3). В грудном плавнике чаще 18—20 лучей. На боках тела темные поперечные полосы. Позвонков чаще 74—76. 2. Anarhichas lupus L.
6 (5). Сошниковый ряд зубов простирается назад почти так же далеко, как и нёбные ряды, или слегка заходит за них (рис. 111, 2). В грудном плавнике чаще 21—22 луча. На спине и боках тела отчетливые темные пятна. Позвонков чаще 78—79. 1. Anarhichas minor Olafs.