(Oncorhynchus Suckley, 1861) Род Онкоринхусы, Онкоринхи, Тихоокеанские лососи, Genus Oncorhynchus Suckley, 1861 (Pacific salmon, Pacific trout) 14 видов
Синонимы:
Hypsifario Gill, 1862
Onchorhynchus
Onchorrhychus
Onchorynchus Suckley, 1861
Parasalmo Vladykov, 1972
Материал подготовил Фактистов И. Е., 2021 г. Сайт www.fishbiosystem.ru
Новый род Oncorhynchus впервые описал в 1861 году американский учёный Джордж Сакли (Джордж Сакли — George Suckley, 1830 — 1869, американский врач и естествоиспытатель, известный как исследователь территорий Вашингтона (Washington) и Орегона (Oregon) в 1850-х годах, учился в Колледже врачей и хирургов (College of Physicians and Surgeons), ныне Колумбийский университет (Columbia University), в 1851 году получил степень доктора медицины, впоследствии работал хирургом в больнице Нью-Йорка, в апреле 1853 года был назначен помощником хирурга и естествоиспытателем Тихоокеанской железной дороги (Pacific Railroad Survey), в 1856 году ушёл в отставку, чтобы заниматься естествознанием, опубликовал несколько работ по естественной истории Северо-Запада Тихого океана, описал несколько новых таксонов, в его честь названо 2 вида рыб).
Онкоринхусы, Онкоринхи, Тихоокеанские лососи (Oncorhynchus) — род лучепёрых рыб из отряда лососеобразных (Salmoniformes Bleeker, 1859) семейства лососёвые (Salmonidae Jarocki or Schinz, 1822), обитающих в естественных водоемах, впадающих в Тихий океан и в прилегающих морях. Типовой вид — Oncorhynchus gorbuscha (Walbaum, 1792).
Название рода Oncorhynchus происходит от древнегреческих слов ογκος (onkos) — «крюк», «изгиб», «кривой» и ρυγχος (rhynchos) — «рыло», «морда», «нос», и относится к крючковатым челюстям самцов («kype») в брачный период (вторичный половой признак).
Несколько позднемиоценовых (около 7 миллионов лет назад — Mya) окаменелостей из Айдахо (Idaho), похожих на форель, которые были найдены в отложениях озера Кларкия (Clarkia Lake), по-видимому, принадлежат роду Oncorhynchus. Наличие этих видов в глубине суши показало, что до начала плиоцена (5—6 млн лет назад) представители рода Oncorhynchus присутствовали не только в водах Тихого океана. То есть, линии радужной форели (rainbow trout — Oncorhynchus mykiss) плюс красногорлого лосося (cutthroat trout — Oncorhynchus clarkii) и тихоокеанских лососей разошлись до начала плиоцена. Следовательно, раскол между Oncorhynchus и Salmo (атлантический лосось) должен был произойти задолго до плиоцена. Предполагаемые даты восходят к раннему миоцену (около 20 млн лет назад).
Один ископаемый вид, отнесенный к этому роду, Oncorhynchus rastrosus (см. фото), саблезубый лосось (синоним Smilodonichthys), представляет собой вид длиной около 3 м, известный в окаменелостях от позднего миоцена до плейстоцена. Oncorhynchus rastrosus обитал вдоль тихоокеанского побережья Северной Америки, в пресных водоёмах и древних реках Калифорнии, доказательством служат окаменелые останки Oncorhynchus rastrosus, которые находят учёные-палеонтологи и археологи в штатах Калифорния, Орегон, Вашингтон. Oncorhynchus rastrosus имел мелкие зубы и многочисленные жаберные тычинки, что свидетельствует о его рационе питания (планктонные организмы). Два зуба, росших примерно на месте клыков млекопитающих, были у Oncorhynchus rastrosus резко увеличены в размерах, достигая длины в 2—3 см, и имели бивнеобразную или саблеобразную форму.
По поводу зубов Oncorhynchus rastrosus и его связи с современными лососями ответила профессор Калифорнийского университета США, доктор палеонтологических наук Джулия Санки: «Зубы из пресноводных отложений не только крупнее, но также и более загнуты, а кончики этих зубов более изношены и тупы. Кроме того, костные основания более крупнее. Зубы из прибрежных морских отложений меньше, прямее, острее, и их корни меньше. Это подтверждает нашу гипотезу о том, что у Oncorhynchus rastrosus происходили онтогенетические изменения между морским и пресноводным этапами жизни, как у современных лососей сегодня». По мнению ученых, Oncorhynchus rastrosus использовал свои бивнеобразные зубы для схваток и демонстрации друг другу во время нерестового периода, происходившего в древних реках Северной Америки. Длина взрослого самца достигала 3 метров, а масса — до 170—177 кг. Но известны случаи нахождения останков и крупнее, до 200 кг.
Видообразование среди Oncorhynchus было изучено в течение многих десятилетий, и семейное «дерево» (см. рисунок) ещё не полностью разработано для лососей Тихого океана. Исследования митохондриальной ДНК (mtDNA) были завершены для различных видов тихоокеанской форели и лососевых, но результаты не обязательно согласуются с исследованиями окаменелостей или молекулярными исследованиями. По общему мнению, родословные кеты, горбуши и нерки разошлись в последовательности после других видов.
Тихоокеанские лососи имеют продолговатые, сжатые с боков тела, чешуя средних размеров или мелкая (до 220 поперечных рядов). Рот большой, конечный; нижняя челюсть сочленяется с черепом позади вертикали заднего края глаза. Верхняя челюсть достигает заднего края глаза или заходит за него. На переднем конце нижней челюсти у половозрелых самцов нет соединительнотканного крючка, входящего в выемку между передними концами praemaxillaria. У молодых, сплывающих из рек в море, зубы хорошо развиты; у взрослых особей, начинающих входить из моря в реки, зубов на челюстях нет, но в соединительной ткани по краям челюстей имеются не прикрепленные к челюстям, еще непрорезавшиеся зубы. Такие же неприкреплённые зубы развиваются на сошнике, нёбных и языке. Заглазничные косточки достигают переднего края praeoperculum. Киля на спинной части хрящевого черепа нет. В анальном плавнике (10) 12—16 ветвистых лучей. Позвонков 63—71. Икра крупная (5—7 мм). Для икрометания тихоокеанские лососи входят в реки, после нереста гибнут. К моменту наступления нереста рыбы сильно изменяются — тело сплющивается с боков, челюсти удлиняются и изгибаются, на челюстях, сошнике, нёбных и языке вырастают крупные крючковидные зубы, кожные покровы утолщаются и окрашиваются в яркие цвета (красный, зеленый, черный и др.).
Распространение (карта): северная часть Тихого океана и в прилежащей части Северного Ледовитого океана. Их ареал простирается от Берингии (в настоящее время это название распространяется на территории, окружающие Берингов пролив, Чукотское и Берингово моря. Включает в себя часть Чукотки и Камчатки в России, а также Аляски в США) на юг, примерно до Тайваня на западе и Мексики на востоке. В Северной Америке некоторые подвиды Oncorhynchus clarkii обитают в Скалистых горах (Rocky Mountains) и Большом бассейне (Great Basin), в то время как другие происходят из Рио-Гранде (Rio Grande) и западных притоков бассейна реки Миссисипи (Mississippi River Basin), которые впадают в Мексиканский залив, а не в Тихий океан. Несколько видов Oncorhynchus были завезены (интродуцированы) в чужеродные водоёмы по всему миру, создавая самостоятельные (самоподдерживающиеся) дикие популяции.
Радужная форель (Oncorhynchus mykiss) — наиболее широко интродуцированный вид этого рода. Радужная форель, чавыча (Oncorhynchus tshawytscha) и кижуч (Oncorhynchus kisutch) создали дикие самоподдерживающиеся популяции в Великих озерах, а чавыча и в Новой Зеландии (известная там как квиннат — «quinnat», королевский — «king» или весенний лосось — «spring salmon»). Аквакультура чавычи, кижуча и радужной форели — основные отрасли промышленности в Чили и Австралии. Чавычи из Чили были выпущены в реки Аргентины, а в Патагонии уже были промыслы кижуча, нерки и радужной форели.
Шесть тихоокеанских лососей Oncorhynchus (см. рисунок) проходные виды и погибают после нереста (семеларность — semelparity) (см. рисунок). На миграцию могут влиять паразиты. Заражённые особи могут стать слабыми и, вероятно, у них сокращается продолжительность жизни. Заражение паразитами вызывает эффект, известный как выбраковка, при котором у инфицированной рыбы меньше шансов завершить миграцию. Проходные формы Oncorhynchus mykiss, известные как стальноголовый лосось (Steelhead), являются итеропарными (iteroparity — не погибают после нереста). Прибрежные лососи Кларка (Oncorhynchus clarkii clarkii) считаются полупроходным, поскольку они проводит некоторое время в океане, обычно гораздо ближе к своему естественному месту обитания, чем его проходные родственники.
Общее сокращение популяции тихоокеанских лососей началось в середине XIX века. В результате западной экспансии и развития США, по оценкам экспертов, популяции лосося в бассейне реки Колумбия (Columbia River) к 1933 году сократились до 20% от уровня, существовавшего до 1850 года. В 2008 году подсчитали, что запасы тихоокеанских лососей на северо-западе Тихого океана составляли менее 10% от их численности до 1850 г. Во многих из оставшихся промыслов лосося преобладает выращенный в инкубаториях лосось, а не дикий. Многие изолированные подвиды тихоокеанской форели, особенно радужная форель — Oncorhynchus mykiss и красногорлый лосось — Oncorhynchus clarkii, сократились в своих естественных ареалах, как и многие местные популяции или отдельные сегменты популяций анадромных форм стальноголового лосося (Oncorhynchus mykiss). В результате этого ряд популяций, видов или подвидов Oncorhynchus государственными, федеральными или международными властями были занесены в список находящихся под угрозой исчезновения или в качестве «видов, вызывающих особую озабоченность». Два подвида Oncorhynchus clarkii считаются вымершими. Уменьшение объясняется широким спектром причин — переловом, утратой и деградацией среды обитания, искусственным размножением, зарыблением и гибридизацией с интродуцированными неместными видами или конкуренцией с ними. Например, желтопёрый лосось Кларка (Oncorhynchus clarkii macdonaldi) вымер в результате интродукции в его естественные воды неместной радужной форели.
В августе 2020 года исследователи сообщили, что повсеместное сокращение размеров тихоокеанских лососей привело к значительным потерям для экосистем и людей, что, по их оценкам, связано с рядом факторов, включая изменение климата и конкуренцию с растущим числом диких и заводских лососей.
Снижение численности дикого лосося из-за чрезмерного вылова рыбы оказало большее давление на рыбоводные заводы, которые должны увеличить производство и восстановить запасы дикого лосося для обеспечения рыболовства. Проблема в том, что инкубаторы никогда не могут полностью воспроизвести среду обитания дикого лосося, проблема, которая часто приводит к физиологическим и поведенческим различиям между диким лососем и лососем, выращенным в инкубаториях. Эти различия часто являются результатом генетических изменений, связанных с инбридингом, искусственным отбором и естественным отбором, а также из-за различного давления окружающей среды, действующего больше на заводских рыб, чем на дикие популяции. Из-за избирательного подхода к размеру промысла в пользу более крупной рыбы со временем наблюдается уменьшение среднего размера взрослых особей лосося. Более мелкий лосось составляет большую часть оставшихся особей, продолжающих популяцию, и проблемы возникают, когда эта выращенная в заводских условиях рыба вводится в дикие популяции. В отличие от дикого лосося, в инкубаториях отбирается более крупный лосось, который, как правило, намного крупнее дикого лосося. В результате лосось, выращиваемый в инкубаториях, имеет тенденцию превосходить дикого лосося в борьбе за пространство, еду и другие ресурсы. Некоторые виды лосося в инкубаториях проявляют хищное поведение по отношению к дикому лососю, потому что они вырастают намного крупнее. Независимо от того, наблюдается ли хищничество, естественное социальное взаимодействие нарушается из-за выпуска большого количества выращиваемого на заводе лосося, где дикие популяции малочисленны, потому что лосось в питомниках, естественно, более склонен к агрессивному поведению.
Рыбы открытого океана. Н.В.Парин 1998 г.
Мезоэпипелагические рыбы прибрежного происхождения, которые встречаются в открытых водах только в период нагула, по размножаются у берегов или даже в пресных водах, составляют очень характерный элемент населения эпипелагиали. Они нередко образуют значительную биомассу, в некоторых районах соизмеримую с биомассой постоянных обитателей эпипелагиали или даже превышающую ее. Типичными представителями этой группировки являются, в частности, настоящие (род Salmo) и тихоокеанские (Oncorhinchus) лососи. Представители последнего рода — кета, горбуша, нерка, кижуч, чавыча, сима — эндемичны для бассейна северной части Тихого океана и в период нагула являются наиболее массовыми рыбами поверхностного слоя на огромной акватории — от Берингова моря до 40° с. ш. Тихоокеанские лососи образуют многочисленные репродуктивные стада, нерестящиеся в реках дальневосточного побережья Азии (на юг до п-ова Корея и Японии; жилая форма нерки есть даже на о-ве Тайвань) и Северной Америки (до Калифорнии). Ареалы таких стад в открытом океане в значительной степени перекрываются, и между 165° в. д. и 155° з. д. даже азиатские и американские лососи встречаются совместно. Длительность морского периода жизни у разных видов неодинакова — от 1,5 лет у горбуши до 4—7 лет у чавычи, наиболее крупного вида, иногда достигающего 50 кг. Они питаются среднеразмерными и крупными планктонными животными — ракообразными, крылоногими моллюсками, молодью кальмаров и рыб. Границы морских ареалов тихоокеанских лососей претерпевают значительные сезонные изменения, однако даже в зимний период область их распространения не простирается за пределы субарктических вод.
Хорошо известно, что границы ареалов эпипелагических рыб, принадлежащих к нектону (т. е. способных к активным горизонтальным перемещениям) , определяются главным образом температурой воды. Связь распространения с температурными условиями показана для многих рыб — акул, лососей, летучих рыб, тунцов, марлинов и др. Температурный диапазон встречаемости в океане лососей рода Oncorhynchus полностью укладывается в пределы 2,3—12°, причем наиболее холодолюбивым видом является нерка, обычная при 2,3—9°, а наиболее теплолюбивым — кижуч, предпочитающий температуру 7—12°. Подобные примеры, количество которых может быть многократно увеличено, вполне убедительно демонстрируют значение температуры воды как главного фактора, лимитирующего географическое распространение нектонных рыб.
Значительные перемещения многих нектонных рыб в пределах своих ареалов, связанные с периодическими нерестовыми, нагульными и зимовальными миграциями, приводят к существенным изменениям границ распространения отдельных видов в разные сезоны. Особенно сильно меняется распространение в океане у тех рыб, которые обитают в водах, подверженных заметным сезонным изменениям температуры, — у сайры, лососей, некоторых тунцов и акул и т. п.
Иллюстрированная энциклопедия рыб. Ст.Франк 1983 г.
Виды рода тихоокеанских лососей Оncorhynchus живут в бассейне северной части Тихого океана, где они представлены 6 видами кета, горбуша, нерка, чавыча, кижуч и сима. Наиболее крупным из них является чавыча (Oncorhynchus tschawytscha), достигающая 1.5 м длины и 45 кг веса. Все тихоокеанские лососи после нереста в реках погибают.
Словарь названий пресноводных рыб СССР. Г.У.Линдберг и А.С.Герд 1972 г.
4.4.0. Oncorhynchus Suckley, 1861 — Тихоокеанские лососи.
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
Тихоокеанские лососи (Oncorhynchus),
как показывает название, обитают в
бассейне Тихого океана. У представителей
этого рода в анальном плавнике от 10 до
16 ветвистых лучей, чешуя средних
размеров или мелкая, икринки крупные и
окрашены в красно-оранжевый цвет. Это
проходные рыбы, нерестующие в
пресных водах Азии и Северной Америки и
нагуливающиеся в море. Известно 6 хорошо
различающихся видов (кета, горбуша,
чавыча, красная, кижуч и сима). Все
тихоокеанские лососи мечут икру лишь раз
в жизни, погибая после первого нереста.
Еще первооткрыватель Камчатского
полуострова Владимир Атласов сообщал в
своей «скаске»: «А рыба в тех реках в
Камчатской земле морская, породою
особая... И идет той рыбы из моря по тем
рекам гораздо много и назад та рыба в
море не возвращается, а помирает в тех
реках и в заводях».
В морской период жизни тихоокеанские
лососи нагуливаются во всей северной
части Тихого океана вплоть до фронта
теплого течения Куро-Сиво, включая
Японское, Охотское и Берингово моря.
В это время они не образуют больших
скоплений и держатся в верхних слоях
(обычно до 10 м глубины). Пища их
разнообразна; чаще всего встречаются в
желудках мелкие пелагические рыбы и их
молодь, ракообразные, пелагические
крылоногие моллюски, молодь кальмаров,
черви, реже медузы и мелкие гребневики.
Тело лососей в это время покрыто
серебристой, легко опадающей чешуей, зубов
на верхней и нижней челюстях нет. Зиму
они проводят на юге, в зоне фронта Куро-Сиво. С наступлением весны океан
оживает: как только повышается температура
верхних слоев, в них обильно
развиваются микроскопические водоросли, к
поверхности поднимаются и начинают
интенсивно размножаться и расти
разнообразные пелагические животные. Эта зона
обильного развития жизни перемещается
от фронта Куро-Сиво на север и северо-восток, по мере того как прогревается вода.
Вслед за ней движутся лососи, все время
находясь в полосе, богатой кормовыми
ресурсами. Этим и объясняется их быстрый
рост в море. Перемещаясь за пищей,
тихоокеанские лососи доходят до устьев рек
северотихоокеанского побережья США,
Канады, Аляски и всего дальневосточного
побережья Азии до Южной Кореи и Японии.
Здесь их стада разделяются. Те, которые
не идут на нерест в этом году, после
откорма с наступлением осеннего
похолодания воды начинают обратную миграцию
на юг. Половозрелые начинают
нерестовую миграцию — путешествие без
возврата, устремляясь в реки, где они
родились и где им суждено, отложив икру,
погибнуть. Неизвестен ни один случай
переживания дальневосточными лососями
нереста, и этим они отличаются от всех
других лососевых. Замечательно, что
лососи, по-видимому, находят ту реку,
в которой они родились. Причины этого
не разгаданы до конца. Есть
предположения, что в открытом море они
ориентируются по солнцу, луне, возможно,
ярким созвездиям, а у берегов «узнают»
воду «родной» реки, отличая тончайшие
особенности ее химического состава с
помощью органов обоняния и вкуса.
Впрочем, эта загадка еще ожидает
разрешения. Внешний вид лососей, входящих
в реки, меняется. У них появляется
«брачный наряд»: тело, бывшее в море
вальковатым, уплощается, на челюстях,
сошнике, нёбе и языке появляются
сильные крючковатые зубы. Сами челюсти,
особенно у самцов, искривляются, на
спине вырастает горб, кожа становится
толстой и грубой, в нее врастает чешуя.
Серебристая окраска исчезает, и в коже
появляется пигмент, окрашивающий ее
в черный, малиновый или лилово-красный
цвет. У самок признаки брачного наряда
выражены слабее, чем у самцов.
Причины возникновения брачного
наряда не изучены. Одни исследователи,
согласно теории полового отбора Чарлза
Дарвина, предполагают, что атрибуты
брачного наряда привлекают самок,
выбирающих «красивейшего» самца, другие
видят в них приспособления, полезные
рыбам в условиях рек. Есть мнение, что
брачный наряд лососей — явление
атавистическое, возврат к предковому типу;
мнение это основано на поверхностном
сходстве в окраске тела и озублении
челюстей у половозрелых рыб и мальков.
Наконец, не исключается возможность,
что брачный наряд обусловлен побочным
действием гормонов, так как во время
интенсивного созревания гонад активно
работают железы внутренней секреции,
особенно гипофиз. Какая из точек зрения
ближе к истине, покажет будущее.
Во время миграции от устьев рек к
местам нерестилищ лососи не питаются,
существуя исключительно за счет запасов,
накопленных в мышцах. Они крайне
истощаются во время пути. Поднимаясь на
1200 км по Амуру, Уссури и р. Хор, кета
теряет более 75% накопленной в море
энергии. Количество жира в мышцах
снижается от 10% до долей процента,
уменьшается также количество сухого вещества,
мясо становится водянистым и дряблым.
Желудок и кишечник съеживаются,
печень перестает вырабатывать желчь,
ферменты, расщепляющие белки,
желудком не выделяются. Все это время рыбы
выполняют огромную работу, поднимаясь
вверх по течению рек, нередко бурных,
изобилующих перекатами, порогами и
водопадами. Установлено, что водопады
высотой в метр и даже более
преодолеваются лососями сравнительно легко.
Рекордсмен в этом отношении чавыча,
поднимающаяся по р. Юкон до озера Беннета и
Карибу — Кроссинг (около 4000 км).
Относительно кеты есть расчеты,
показывающие, что суточный расход энергии для
самцов составляет 25 810 и для самок 28 390
калорий на килограмм живого веса.
Нерестовая миграция лососей при их
большой численности оставляет
неизгладимое впечатление. Вот как описал ее
первый ученый, исследовавший
Камчатку, С. П. Крашенинников: «Все рыбы
на Камчатке идут летом из моря в реки
такими многочисленными рунами, что
реки от того прибывают и, выступя из
берегов, текут до самого вечера, пока
перестанет рыба входить в их устья».
Описание Крашенинникова относится к 1737—1741 гг., и до начала нашего века его
нельзя было считать преувеличенным.
В настоящее время численность
тихоокеанских лососей сильно сократилась и
нерестовый ход уже не представляет столь
грандиозного зрелища.
Все тихоокеанские лососи закапывают
оплодотворенную икру в грунт, поэтому
нерестятся они в местах, где дно не заилено,
покрыто галькой или гравием, нередко
там, где бьют подводные ключи. Самка,
сопровождаемая одним или несколькими
самцами, держится головой против
течения и энергичными движениями
хвостового стебля разбрасывает грунт. Икра
откладывается в образовавшуюся яму,
и самец поливает ее молоками. Между
самцами во время нереста происходят
непрерывные стычки. Часть икринок
остается неоплодотворенной, многие уносятся
течением и поедаются пресноводными
рыбами. Выметав икру, самка забрасывает
яму галькой. Образуется бугор, под
которым икринки проходят развитие и
вышедшие из икры личинки находятся до
рассасывания желточного мешка.
После нереста начинается массовая
гибель производителей. Наиболее
истощенные погибают уже на нерестилище,
другие относятся течением и гибнут по
дороге к устью. Дно и берега рек
покрываются мертвой рыбой (ее у нас на
Дальнем Востоке называют сненкой). На этот
обильный корм собирается множество
ворон, чаек и самых разных зверей, вплоть
до медведей.
Мальки, как только рассосется желточный
пузырь, выходят из бугра и сплывают вниз
по течению, питаясь мелкими водными
беспозвоночными и упавшими в воду
насекомыми. У одних видов они не
задерживаются долго в реке, у других речной
период растягивается до одного-двух лет.
Иногда часть самцов достигает половозрелости в реке, имея очень небольшие
размеры; такие карликовые самцы могут
принимать участие в нересте. Наконец,
некоторые виды образуют настоящие жилые
пресноводные формы, не выходящие в
море. Подобные формы вообще
распространены в семействе лососевых.
Рыбы Японского моря и сопредельных частей Охотского и Желтого морей. Часть 2. (Acipenseriformes — Polynemiformes). Г.У.Линдберг, М.И.Легеза 1965 г.
1. Род ONCORHYNCHUS SUCKLEY, 1861 — ТИХООКЕАНСКИЕ ЛОСОСИ
Oncorhynchus Suckley, Ann. Lyceum Nat. Hist., New York, 7, 1861 : 313 (subg. тип: Salmo scouleri Suckley = Salmo gorbuscha Walbaum).— Берг, Рыбы пресных вод, 1, 1948 : 169. — Nеavе, Trans. Royal Soc., Canada, 52, (3), 5, 1958 : 25—39 (происхождение рода Oncorhynchus).
Рот большой, нижняя челюсть сочленяется с черепом позади вертикали заднего края глаза. Этим признаком отличается от малоротых лососевых — Brachymystax и Coregonus, у которых нижняя челюсть сочленяется с черепом впереди или под названной вертикалью. Верхнечелюстная кость длинная, у взрослых хватает до заднего края глаза (или еще далее); в части, образующей край челюсти, не имеет изгиба. Заглазничные косточки хватают до praeoperculum. Зубы у молодых по сравнению с сигами сильно развитые (у взрослых Oncorhynchus перед входом в реку зубы еще зачаточны и прорезаются уже в реке). В анальном плавнике 10—16 ветвистых лучей. Чешуя умеренно мелкая или мелкая (до 220 поперечных рядов). Яйца крупные. Позвонков 63—71. (Берг, 1948 : 169—171).
Северная часть Тихого океана и от Берингова пролива на запад до Лены, а на восток до Маккензи.
6 видов. В водах Японского моря 4 вида и в сопредельных с ним водах еще 2 вида. Туводные формы, не выходящие в море, рассматриваются как производные от проходных видов, но в некоторых случаях — как самостоятельные виды. Недавно описан новый вид Oncorhynchus iwame Kimura et Nakamura, 1961, близкий к Oncorhynchus rhodurus Jordan et McGregor, 1925 (Kimura a. Nakamura, 1961 : 69—74, fig. 3).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ РОДА ONCORHYNCHUS (По Бергу (1948 : 172) с дополнениями.)
1 (10). Чешуя сравнительно крупная, не более 150—160 поперечных рядов.
2 (9). Жаберных тычинок на первой дуге не более 27.
3 (8). Жаберных лучей не более 15, обычно меньше.
4 (7). Пилорических придатков менее 100.
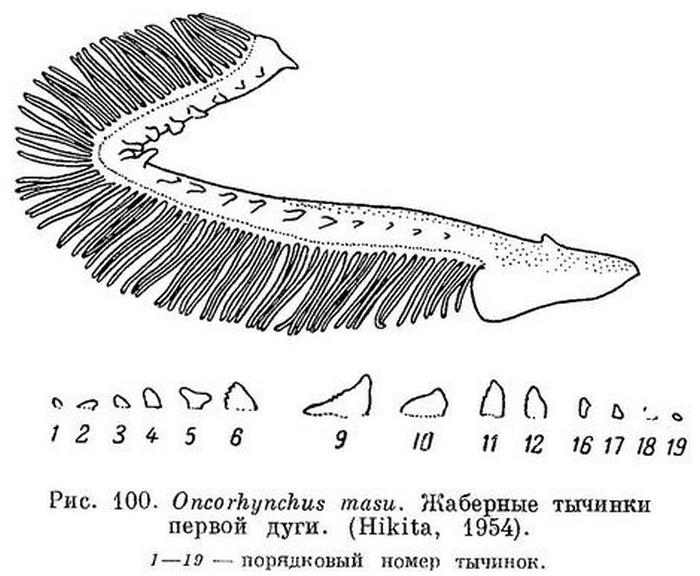
5 (6). У половозрелых в реке на теле появляются более или менее ясные, темные поперечные полосы, выше и ниже полос обычно довольно крупные черные пятна. A III—IV 11—14 (15), l. l. 130—140, жаберных лучей 11—15, жаберных тычинок, очень коротких (рис. 100) 18—22, пилорических придатков 35—76. Хвостовой плавник без пятен. Характер жаберных тычинок у мазу и кижуча показан на рисунках у Хикиты (Hikita, 1954 : 195. рисунок на стр. 198). 1. Oncorhynchus masu (Brevoort).
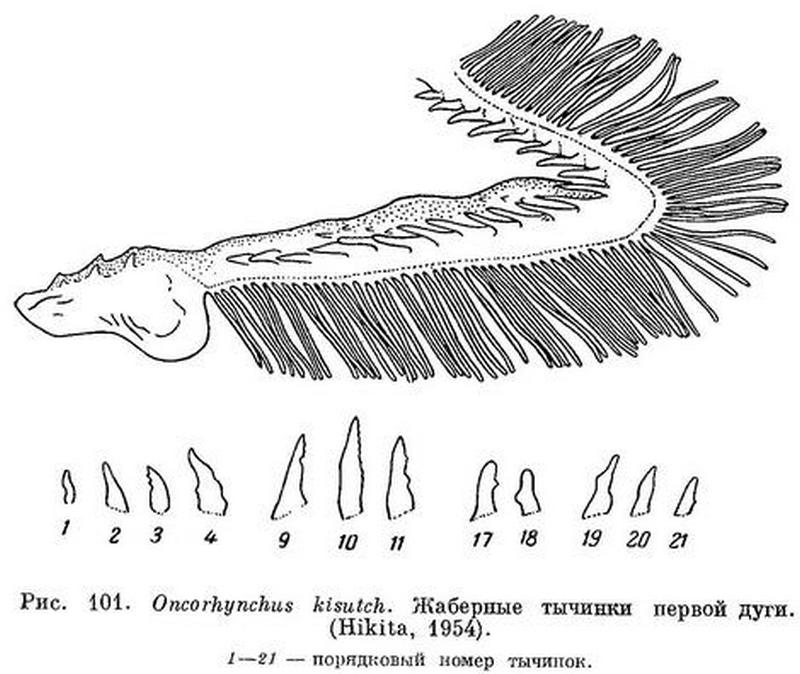
6 (5). У половозрелых на теле темных поперечных полос и крупных, округлых пятен нет (мелкие круглые пятнышки есть). А IV—V 12—14, l. l. 120—140, жаберных лучей 13—14, жаберных тычинок заметно удлиненных (рис. 101) 19—23, пилорических придатков 45—81. Верх хвостового плавника с пятнами. 2. Oncorhynchus kisutch (Walbaum).
7 (4). Пилорических придатков 135—185. А III 12—15, l. l. 125—150, жаберных лучей (11) 12—15, жаберных тычинок 19—25. Хвостовой плавник без пятен. 3. Oncorhynchus keta (Walbaum).
8 (3). Жаберных лучей 15—19, А III 15—16, l. l. 135—155, жаберных тычинок 23—27, пилорических придатков 140—185. Хвостовой плавник с многочисленными мелкими пятнами. 4. [Oncorhynchus tschawytscha (Walbaum)].
9 (2). Жаберных тычинок 30—40. A III—IV 12—16, l. l. 130—144, жаберных лучей 13—16, пилорических придатков 75—95. Хвостовой плавник без пятен. 5. [Oncorhynchus nerka (Walbaum)].
10 (1). Чешуя мелкая, 177—240 поперечных рядов. Жаберных тычинок 26—33, жаберных лучей 10—14, А II—IV 12—16. Хвостовой плавник с крупными пятнами. 6. Oncorhynchus gorbuscha (Walbaum).
Рыбы северных морей СССР. А.П.Андрияшев 1954 г.
1. Род ONCORHYNCHUS SUCKLEY — ТИХООКЕАНСКИЕ ЛОСОСИ
Oncorhynchus Suckley, Anil. Lyceum Nat. Hist. New York, VII, 1861 : 313 (subgenus; тип: Salmo scouleri Rich. = Salmo gorbuscha Walb.). — Берг, Рыбы пресн. вод СССР, I, 1948 : 168.
Рот большой, конечный; нижняя челюсть сочленяется с черепом позади вертикали заднего края глаза. Верхняя челюсть достигает заднего края глаза или заходит за него. На переднем конце нижней челюсти у половозрелых самцов нет соединительнотканного крючка, входящего в выемку между передними концами praemaxillaria. У молодых, сплывающих из рек в море, зубы хорошо развиты; у взрослых особей, начинающих входить из моря в реки, зубов на челюстях нет, но в соединительной ткани по краям челюстей имеются не прикрепленные к челюстям, еще непрорезавшиеся зубы. Такие же неприкрепленные зубы развиваются на сошнике, нёбных и языке. Заглазничные косточки достигают переднего края praeoperculum. Киля на спинной части хрящевого черепа нет. В анальном плавнике (10) 12—16 ветвистых лучей. Позвонков 63—71. Икра крупная (5—7 мм).
6 видов в северной части Тихого океана и в прилежащей части Сев. Ледовитого океана. Для икрометания входят в реки, после нереста гибнут. К моменту наступления нереста рыбы сильно изменяются — тело сплющивается с боков, челюсти удлиняются и изгибаются, на челюстях, сошнике, нёбных и языке вырастают крупные крючковидные зубы, кожные покровы утолщаются и окрашиваются в яркие цвета (красный, зеленый, черный и др.). Морской период жизни слабо изучен. Чрезвычайно ценные промысловые рыбы, уловы которых на Дальнем Востоке ежегодно составляют до 150 млн шт.
1 (2). Чешуя мелкая, 177—240 поперечных рядов. Хвостовой плавник без серебристого налета, весь покрыт темными пятнами. Жаберных
тычинок на 1-й дуге 26—33. Жаберных лучей 10—14. 2. Oncorhynchus gorbuscha (Walb.).
2 (1). Чешуя крупнее, не более 150—160 поперечных рядов. Хвостовой плавник без пятен или на нем есть мелкие темные пятнышки (в последнем случае между лучами С есть серебристый налет). Серебристый налет между лучами хвостового плавника хорошо заметен на свежих экземплярах, но после фиксации пропадает (то же, естественно, относится и к цвету мяса).
3 (8). Жаберных тычинок не более 27. Между лучами хвостового плавника (особенно у основания С) есть серебристый налет.
4 (5). Жаберных лучей 15—19. Ветвистых лучей в анальном плавнике 15—16. На спине мелкие темные х-образные пятнышки; весь хвостовой плавник покрыт мелкими округлыми пятнышками. Жаберных тычинок 23—27. (Чавычу от кижуча легко отличают по цвету «дёсен»: у кижуча мякоть, в которой сидят челюстные зубы, светлая (белая), а у чавычи вся ротовая полость, включая и «дёсны», всегда темная (F. В. Hagerman, Calif. Fish a. Game, v. 37, 1, 1951 : 53).) 3. Oncorhynchus tschawytscha (Walb.).
5 (4). Жаберных лучей 11—15. Ветвистых лучей в А 12—15. На хвостовом плавнике нет темных пятнышек или они расположены преимущественно по его верхнему краю. Жаберных тычинок 19—25.
6 (7). Пилорических придатков 45—81. На спине и в верхней части хвостового плавника мелкие темные округлые пятнышки. Хвостовой плавник усечен или со слабой выемкой. Мясо красного цвета. 5. Oncorhynchus kisutch (Walb.).
7 (6). Пилорических придатков 135—185. На спине и плавниках нет темных пятнышек. Хвостовой плавник с заметной выемкой. Мясо бледное, оранжевато-розовое. 1. Oncorhynchus keta (Walb.).
8 (3). Жаберных тычинок 30—40. На хвостовом плавнике нет серебристого налета. На теле и плавниках нет темных пятен. Пилорических придатков 75—95. 4. Oncorhynchus nerka (Walb.).
Рыбы пресных вод СССР и сопредельных стран. Л.С.Берг 1948 г.
Род ONCORHYNCHUS SUCKLEY. — ТИХООКЕАНСКИЕ ЛОСОСИ
Oncorhynchus Suckley, Ann. Lyceum Nat. Hist. New York, VII, 1861, p. 313 (subg.; тип: Salmo scouleri = Oncorhynchus gorbuscha).
Hypsifario Gill, Proc. Acad. Nat. Sc. Philadelphia (1862), 1863, p. 330 (тип: Salmo kennerlyi = Oncorhynchus nerka).
Oncorhynchus Чернавин, 1918, 1921, 1923.
Рот большой, нижняя челюсть сочленяется с черепом позади вертикали заднего края глаза. Верхнечелюстная кость длинная, у взрослых хватает до заднего края глаза (или еще далее); в части, образующей край челюсти, не имеет изгиба. Заглазничные косточки хватают до praeoperculum. Мезэтмоид в виде треугольника. Киля на спинной части хрящевого черепа нет. Рыло хрящевого черепа спереди заострено, не раздвоено. В теменной части хрящевой череп у половозрелых сверху без отверстий. У молоди Oncorhynchus keta и Oncorhynchus masu, а также у половозрелых карликовых самцов Oncorhynchus masu, эти отверстия имеются (А. Я. Таранец, Изв. Тихоокеанск. инст. рыбн. хоз. и океаногр., XII (1937), 1933, стр. 10). Зубы у молодых, по сравнению с сигами, сильно развитые, по сравнению с взрослыми (но не нерестующими) Oncorhynchus развиты слабо. В брачном наряде имеет на челюстях большие зубы, снабженные костными основаниями, коими они прирастают к костям челюстей. На переднем конце нижней челюсти у половозрелого самца нет соединительнотканного крючка, входящего в выемку между передними концами praemaxillaria. В анальном плавнике 10—16 ветвистых лучей. Чешуя умеренно мелкая или мелкая (до 220 поперечных рядов). Яйца крупные. Позвонков 63—71.
Северная часть Тихого океана, откуда входят в реки; на юг до южн. Кореи, Японии и (пресноводная форма) Формозы и зал. Монтри (южнее Сан-Франциско); прилегающая к Берингову проливу часть Ледовитого моря (на запад до Лены, на восток до р. Мэккензи) (J. Dymond и V. Vladykov (Proc. V Рас. Sc. Congr. 1933, Toronto, 1934, p. 3743), говорят, что имели в своих руках Oncorhynchus sp. (Oncorhynchus keta?) из Большого Невольничьего озера в системе р. Мэккензи.). Некоторые живут постоянно в пресной воде. Шесть видов.
У молоди Oncorhynchus, сплывающей из рек в море, хорошо развиты зубы. У Oncorhynchus, входящих из моря в реку и еще не имеющих ясных признаков брачного наряда, ни на верхней, ни на нижней челюсти нет зубов, но в соединительной ткани по краям челюстей имеются не прикрепленные к челюстям еще не прорезавшиеся или едва прорезавшиеся зубы. Такие же неприкрепленные зубы имеются на сошнике, на нёбных и на языке (Чернавин). В реке тело рыбы сплющивается с боков, у самцов образуется горб (у одних большой, у других меньший), челюсти удлиняются (Особенно сильно удлиняются praemaxillaria у самца.) и изгибаются; на челюстях, сошнике, нёбе и на языке появляются сильные крючковатые зубы; особенно сильно изгибаются челюсти у самцов. Затем сильно увеличиваются глоточные зубы. Кожа сильно утолщается, и на ней появляется яркий пигмент — черный или лилово-красный; чешуя становится почти незаметной. Серебристая, блестящая окраска исчезает. Во все тремя миграции к нерестилищам рыба в реке ничего не ест. Различия между самцами и самками в брачном наряде заключаются в следующем: у самок или совсем не появляется горба или появляется горб лишь слабо развитой; рыло и челюсти удлиняются гораздо меньше, зубы слабее, тело не уплощается так сильно, окраска менее интенсивна.
Последовательность появления лососей в р. Камчатке в 1927 г. была такова:
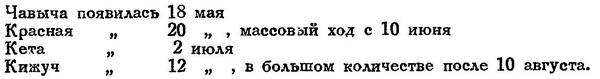
Соотношение пород лососей в уловах у нас на Дальнем Востоке и в Америке (Соединенные Штаты) видно из следующей таблички, заимствованной у В. К. Бражникова (1920) и показывающей число штук в миллионах:

Последовательность вхождения этих видов в наши воды такова: чавыча, красная, сима, горбуша, кета, кижуч.
Разные виды рода Oncorhynchus приурочены преимущественно к определенным рекам. Так, кета занимает заметное место в Анадыре и в Амуре, красная — в р.Камчатке, горбуша — в р. Большой и в Амуре, сима — в р. Тумнин.
Все представители рода Oncorhynchus мечут икру раз в жизни, а затем погибают. Об этом писал еще Крашенинников (1755, т. I, ч. 2, гл. VIII): «во всех родах камчатских лососей сие достойно примечания, что они в реках и родятся и издыхают, а возрастают в море, и что по однажды токмо в жизнь свою икру и молоки пускают».
По словам И. И. Кузнецова (1928, стр. 109), кета может скрещиваться с горбушей, красной и кижучем. На Амуре был произведен опыт скрещивания горбуши и летней кеты. На Камчатке названному автору однажды пришлось видеть, «как огромный самец кеты, отогнав самцов красной, несколько дней нерестовал один с самкой красной; в другом месте, наоборот, самка кеты весь процесс нереста закончила с самцом красной» (стр. 165).
Врагами лососей являются на Амуре мальма, ленок, хариус и почти все виды гольянов, пожирающие их икру во время нереста, на Камчатке — мальма (и ее речная форма), микижа, кунджа и хариус. На Камчатке особенный вред приносят мелкие гольцы (Salvelinus), остающиеся в реках круглый год; они подбирают ту икру, которая выносится течением из гнезд, а также ту икру кеты или красной, которую выбрасывает из их гнезд кижуч. Около нерестующего кижуча бывает обычно 3—5 небольших гольцов длиной от 7 до 25 см, а иногда число их доходит до 50. В кишечнике гольца находили до 300 лососевых икринок (Кузнецов, 1928, стр. 177, 185).
1 (11). Чешуя сравнительно крупная, не более 150—160 поперечных рядов.
2 (9, 10). Жаберных тычинок на 1-й дуге не более 27.
3 (8). Жаберных лучей не более 15, обычно меньше.
4 (7). Пилорических придатков менее 100.
5 (6). У половозрелых на теле более или менее ясные, темные поперечные полосы, выше и ниже полос обычно довольно крупные черные пятна. A III—IV 11—14(15), 1. 1. 130—140, жаберных лучей 11—15, жаберных тычинок 18—22, пилорических придатков 35—76. 6. Oncorhynchus masu (Brevoort)
6 (5). У половозрелых на теле темных поперечных полос и крупных
округлых пятен нет (мелкие круглые пятнышки есть). А IV—V 12—14, l. l. 120—140, жаберных лучей 13—14, жаберных тычинок 19—23, пилорических придатков 45—81. 5. Oncorhynchus kisutch (Walbaum)
7 (4). Пилорических придатков 135—185. А III 12—15, l. l. 125—150, жаберных лучей (11) 12—15, жаберных тычинок 19—25. 1. Oncorhynchus keta (Walbaum)
8 (3). Жаберных лучей 15—19, А III 15—16, l. l. 135—155, жаберных тычинок 23—27, пилорических придатков 140—185 3. Oncorhynchus tschawytscha (Walbaum)
9 (2, 10). Жаберных тычинок 30—40. A III—IV 12—16, l. l. 130-144, жаберных лучей 13—16, пилорических придатков 75—95. 4. Oncorhynchus nerka (Walbaum)
10 (2, 9). Жаберных тычинок 29—36, жаберных лучей 11—13. Озерная форма, не уходящая в море. 4а. Oncorhynchus nerka adonis Jordan et McGregor
11 (1). Чешуя мелкая, 177—240 поперечных рядов. Жаберных тычинок 26—33, жаберных лучей 10—14. А II—IV 12—16. 2. Oncorhynchus gorbuscha (Walbaum)