(Trachurus Rafinesque, 1810) Род Трахурусы, Трахуры, Ставриды, Genus Trachurus Rafinesque, 1810 (Horse mackerel, Jack mackerels, Saurels) 15 видов
Ставриды Мирового океана (род Trachurus). В.В.Некрасов 1994 г.
Описание представителей рода Trachurus
Семейство ставридовых (Carangidae, Perciformes) включает в себя около 200 видов примерно из 30 родов [Линдберг, 1971]. Наиболее широко распространен род Trachurus.
Для рыб этого рода характерно продолговатое, слегка сжатое с боков тело. Максимальная длина до 68 см, масса - до 2850 г. Голова заостренная. Рот большой, нижняя челюсть выдается вперед. Мелкие зубы расположены на челюстях, сошнике, небных костях и на языке. У личинок на предкрышке имеются шипы. Глаза крупные. Жировые веки развиты хорошо. Два отделенных друг от друга спинных плавника. Первый, колючий спинной плавник немного выше второго, мягкого; восемь его колючек соединены перепонкой; перед ним находится направленная вперед скрытая в коже короткая колючка. Второй спинной и анальный плавники длинные, анальный немного короче. Перед анальным плавником две отдельно сидящие колючки, соединенные друг с другом перепонкой. Формула плавниковых лучей: D I, VIII+I, 27—36; А II+I, 23—33. Грудные плавники длинные, значительно длиннее брюшных. Брюшные с хорошо развитой колючкой расположены немного позади оснований грудных. Хвостовой плавник с глубокой выемкой. Хвостовой стебель тонкий. Жаберные тычинки удлиненные. Чешуя мелкая циклоидная. Голова сверху и с боков покрыта чешуей. Боковая линия, вооруженная щитками, в середине тела делает изгиб: передняя ее часть слегка изогнута и лежит высоко, а задняя - прямая и проходит по средней линии тела. Щитки в задней части боковой линии снабжены направленными назад колючками (см.рис.2). Вдоль спины до начала колючего - конца мягкого спинного плавника проходит вторая (спинная) ветвь боковой линии. Большая часть ребер, за исключением передних, сидячих, прикреплена к парапофизам. Позвонков 24 [Алеев, 1957а; Световидов, 1964].

Таксономия рода Trachurus Rafinesque и принципы ее построения
Систематическое положение и филогенетические взаимоотношения представителей рода Trachurus неоднократно обсуждались в специальной литературе. Описано более двух десятков форм ставрид. Некоторые из них первоначально были описаны как виды, другие - как подвиды или вариететы [Nichols, 1920, 1940; Nani, 1950; Tortonese, 1952, 1961; Cousseau, 1967; Некрасов, 1966 и др.], что как бы подтверждало спорность принципов систематики, критериев вида и подвида [Завадский, 1968; Майр, 1971].
Согласно Ч.Дарвину, виды реальны. Реальность - философская категория, означающая существование некоего объекта вне нашего сознания. При этом объект может быть не дискретным, т.е. может не иметь четко означенных границ, но от этого он не перестает быть реальным. В понятие реальности дискретность не входит. Нереальными могут быть лишь границы между ними. Действительно, если мы не принимаем, вслед за де Фризом, что виды порождаются скачком (мутацией) на протяжении одного поколения, то неизбежно должны принять, что видообразование - длительный процесс и можно обнаружить все стадии становления нового вида из какой-либо разновидности старого [Медников, 1975].
Идеи Ч.Дарвина оказали влияние на систематические работы. Так, Дж.Гукер, описывая новый вид растения, указывал в диагнозе среднее значение его признаков, а также возможные отклонения от него. К сожалению, впоследствии получила широкое распространение концепция, которую Э.Майр назвал типологической. Согласно этой концепции вид характеризуется признаками экземпляра (голотипа), по которому он был описан.
Биологи, придерживающиеся типологической концепции вида, в своих исследованиях ограничиваются чисто морфологической характеристикой видов и критерием для их выделения считают наличие хиатуса по отдельным морфологическим признакам. Но еще С.И.Огнев [1944,1947] показал, что у близкородственных видов разрыв может быть неполно выражен или сглажен. С таким положением столкнулся и автор при описании формы Trachurus из Аравийского моря, ошибочно отнеся ее к подвиду Trachurus mediterraneus indicus Necrassov [Некрасов, 1966].
Следует также отметить, что и среди других групп рода Trachurus четкого разграничения особей разных видов по морфологическим признакам в достаточно больших однородных выборках, адекватно отражающих всю величину изменчивости популяций, нет. Отсюда и большое количество в роде Trachurus описанных форм и наличие подвидов. Почти у всех видов рода Trachurus диагностические признаки трансгрессируют.
Трансгрессии по всем морфологическим признакам были обнаружены между Trachurus mediterraneus и Trachurus indicus [Некрасов, 1966], между Trachurus trachurus и Trachurus capensis [Некрасов, 1976], между Trachurus indicus и Trachurus delagoa [Некрасов, 1970]. Трансгрессии по всем морфологическим признакам между другими видами ставрид рода обнаружены многими авторами. Например, между Trachurus novaezelandiae и Trachurus japonicus [Stephenson, Robertson, 1977], между Trachurus novaezelandiae и Trachurus declivis [Stephenson, Robertson, 1977; Gauldie et al.,1977], между Trachurus novaezelandiae и Trachurus trachurus [Алеев, 1957a], между Trachurus picturatus и Trachurus symmetricus [Шабонеев, Котляр, 1979], между Trachurus symmetricus и Trachurus murphyi [Santander, de Castillo, 1971], между Trachurus picturatus и Trachurus aleevi [Рытов, Разумовская, 1984], между Trachurus japonicus, Trachurus novaezelandiae и Trachurus lathami [Nichols, 1920; Berry, Cohen, 1972] и др. К этому следует добавить, что подобная картина имеет место и в других родах семейства ставридовых [Talwar, 1972].
В подобных случаях современные сторонники типологической концепции вида, проведя анализ морфологических признаков по большим выборкам близкородственных видов и не обнаружив хиатуса, рассматривают их на уровне подвидов.
Подчеркнем, что морфологическая характеристика видов при типологической концепции вида раньше давалась по единичным экземплярам. Сегодня очевидно, что единичные особи не могут быть носителями всех свойств, всех признаков вида. Только серия, достаточно представительная выборка из популяции дает объективное представление о размахе морфологических вариаций вида. Однако господствовавшая ранее в теории вида типологическая концепция отвергнута далеко не всеми, несмотря на то, что она справедливо критиковалась [Майр, 1968,1971; Старобогатов, 1977] и, казалось бы, давно утвердилась биологическая концепция вида, отличающаяся от типологической тем, что подчеркивает популяционный аспект и генетическую интегрированность вида. Во многих работах, в том числе и посвященных систематике ставрид, встречаются характерные для такого подхода формулировки, например, такие: "различия между этими формами не выходят за пределы их изменчивости, что заставляет нас относить их к одному виду" или "эти формы столь резко различны, что мы должны считать их разными видами". Короче говоря, простота типологической концепции вида до сих пор подкупает систематиков.
Популяционный аспект биологической концепции вида предполагает привлечение для морфологической характеристики внутривидовых группировок значительно большего числа экземпляров, что влечет за собой стирание хиатуса между большинством близкородственных видов. Но при новом методе, применяемом для определения нетаксономических категорий, приверженцы старой типологической концепции вида продолжают считать критерием вида хиатус. Их не смущает то, что в принципе почти в любом роде животных, обработав большой объем материала, можно убедиться в том, что по диагностическому признаку все виды будут трансгрессировать.
Уже Э.Майр [1971] обратил внимание на недостаток такого подхода в определении видов и посчитал, что чисто интуитивный подход, основанный на совокупности признаков, часто может оказаться гораздо более эффективным, чем механическое использование биометрического анализа или диагностических признаков, потому что всегда имеется доля случаев, когда изменчивость слишком велика и результаты слишком противоречивы, чтобы привести к однозначному разделению видов. Ну, а если дальнейший анализ покажет, что такая форма была ошибочно низведена до подвидового ранга, ей всегда можно снова придать статус вида.
В случае с Trachurus mediterraneus indicus это сделали Ф. Бери и Л.Коэн [Berry, Cohen, 1972], возведя его в ранг вида.
Для построения системы рода Trachurus, естественно объясняющей филогенетические взаимоотношения входящих в него видов, необходимо наряду с морфологической характеристикой рассматривать экологические аспекты, географическое распространение и данные онтогенеза. Все это в полной мере нашло отражение в работах по систематике Ю.Г.Алеева и И.Е.Шабонеева. При этом авторы отнюдь не игнорировали морфологический критерий, а наоборот, исходя из положения А.Н.Световидова [1952, с. 11], что "классификация должна основываться на комплексе морфологических особенностей, функционально связанных с наиболее характерной биологической особенностью рассматриваемой систематической группы и выработанных в результате взаимодействия организма со средой", возвели его на качественно более высокий уровень. Такой подход к проблеме позволил указанным авторам не только дифференцировать в роде Trachurus группы близкородственных видов, но и подойти к вопросам эволюции и происхождения как всего рода, так и отдельных его представителей, наметить несколько филогенетических линий развития.
При построении системы рода Trachurus, кроме биологических и связанных с ними морфологических особенностей отдельных форм, необходимо принимать во внимание также особенности их онтогенетического развития, так как морфологические особенности различных форм рода заметно изменяются в ходе онтогенеза и без учета этих изменений нельзя сравнивать те или иные формы.
Известно, что мальки ставриды длиной 9-12 мм существенно отличаются от взрослых рыб очень высоким и сжатым в латеральном направлении телом. Относительная высота тела у ставрид по мере развития сначала увеличивается, достигая максимума при длине около 1 см, что примерно соответствует концу того периода, в течение которого личинка превращается в малька, а затем снова уменьшается до некоторой величины, обычной для взрослых особей, и при достижении длины примерно 5 см становится более или менее постоянной [Алеев, 19576; Некрасов, 1969].
В ходе онтогенеза у видов рода Trachurus первый спинной и брюшные плавники, а также передние части второго спинного и анального плавников постепенно занимают относительно более переднее положение.
Малек ставриды длиной 2-3 см имеет неудлиненные последние лучи второго спинного и анального плавников, причем эти лучи у малька отделены от предыдущих лучей такими же промежутками, какие имеются между двумя любыми лучами этого плавника. Позднее, при увеличении длины малька, последние лучи указанных плавников отодвигаются от остальной части плавника на более значительное расстояние.
Характерной чертой ставрид является сильное развитие щитков боковой линии. В онтогенезе щитки появляются сравнительно поздно - при длине 15-20 мм. Закладка щитков начинается на прямой части боковой линии, а на ее кривой части они появляются позднее. По мере развития особей относительная величина щитков увеличивается. Величина щитков и их количество у видов рода Trachurus связаны прежде всего со степенью приспособленности отдельных видов и форм рода к подвижному пелагическому образу жизни.
Таковы некоторые моменты онтогенеза, важные для понимания филогенетических отношений различных форм рода Trachurus.
В строении боковой линии у различных видов ставрид наблюдается большое разнообразие. Это касается в первую очередь строения туловищных каналов боковой линии: основных боковых каналов, пронизывающих щитки, и их спинных ветвей. Что касается каналов, расположенных на голове, то в их строении у разных видов рода не замечается резких различий.
У более подвижных форм рода щитки развиты, как правило, сильнее, чем у форм менее подвижных. Ю.Г.Алеев объясняет это тем, что по мере развития килей увеличиваются силы, действующие нормально к поверхности пластинки киля, в связи с чем для сохранения устойчивости киля необходимо расширение его основания, т.е. необходимо удлинение лопастей основания щитка, несущего киль, что и наблюдается у наиболее подвижных форм рода [Алеев, 1957а]. С другой стороны, величина щитков зависит отчасти от их количества: у форм, у которых щитков больше, они, как правило, мельче, и наоборот.
Кроме перечисленных признаков, при построении ключа к определению видов рода Trachurus, принималось во внимание также количество жаберных тычинок, отражающее характер пищи: чем мельче состав пищи, тем больше жаберных тычинок.
Род Trachurus включает в себя как неритические, так и неритоокеанические виды. Известно, что большинство видов современной ихтиофауны океанической эпипелагиали произошли от неритических предков [Алеев, 1957а; Парин, 1968; Шубников, 1974], тогда как в прибрежной области представлены наиболее древние, наиболее низкоорганизованные из ныне живущих костистых рыб. Более того, многие массовые семейства рыб эпипелагиали наряду с типичными океаническими видами включают в себя и очень близкие к ним в систематическом отношении прибрежные формы. Все это подтверждает общепринятое представление о происхождении современной океанической ихтиофауны эпипелагиали от неритических предков. Следовательно, более подвижный образ жизни, в меньшей степени связанный с берегом, является более прогрессивным биологическим типом.
Подходя с этих позиций к современному распространению форм рода Trachurus, нетрудно заметить, что среди них есть обитающие только в неритической зоне континентальных шельфов, пределы которой они не покидают на протяжении всего своего жизненного цикла. Таковыми являются Trachurus trecae, Trachurus indicus, Trachurus mediterraneus и др. В противоположность им Trachurus picturatus, Trachurus symmetricus и Trachurus murphyi могут выходить далеко за границу континентального шельфа, встречаясь на расстоянии сотен и даже тысяч миль от него [Collett, 1896; Roedel, Fitch, 1952; Ahlstrom, 1968; Константинов и др.,1979; Некрасов, 1992]. Ставриды Trachurus trachurus и Trachurus capensis занимают промежуточное положение и могут встречаться лишь на некотором удалении от континентального шельфа [Алеев, 1956; Berry, Cohen, 1972].
Род Trachurus распространен в умеренных, субтропических и тропических водах Мирового океана. В этот род входят наиболее эвритермные и эвригалйнные виды семейства Carangidae. Однако среди отдельных представителей рода имеет место значительная дифференциация по отношению к температуре среды обитания. Границы некоторых наиболее холодолюбивых видов ставрид простираются в северном полушарии до берегов Исландии (64° с.ш.). В южном полушарии граница распространения ставрид находится у берегов Чили на 57° ю.ш.
Долгое время считалось, что род Trachurus имеет биполярное распространение [Берг, 1953; Алеев, 1957а]. Однако позже было показано, что и в тропических водах ставриды представлены достаточно широко [Некрасов, 1966а, 1976, 1987; Berry, Cohen, 1972].
Trachurus trecae, Trachurus lathami, Trachurus indicus наиболее теплолюбивы, и ареалы их расположены в более низких широтах. В частности, известно, что Trachurus trecae обитает при температуре воды у дна 15-22°С, a Trachurus indicus - при 17-20°С [Оверко, 1969а; Некрасов, 1973].
Trachurus delagoa из Индийского океана распространена в условиях придонной температуры выше 17°С и южнее порта Дурбан в уловах не встречается [Некрасов, 1973].
Однако средиземноморская ставрида Trachurus mediterraneus и особенно ее подвид из Черного моря Trachurus mediterraneus ponticus обитают в условиях более низкой температуры [Алеев, 1957]. Trachurus trachurus отмечена у побережья Норвегии и Исландии, а Trachurus capensis - у южной оконечности Африки на отмели Агульяс [Lockwood, Jonson, 1977], где температура воды в местах их обитания равна 9°С. Нерест ставриды в северо-восточной части Атлантического океана (Ла-Манш, Северное море) происходит при температуре воды у дна 9,4-12,5°С [Полонский, Тормосова, 1970], а в центрально-восточной Атлантике - при 13-16°С [Оверко, 1969а]. В более холодных зонах расположены ареалы Trachurus picturatus, Trachurus symmetricus и Trachurus murphyi. Так, северная граница распространения Trachurus picturatus проходит в Бискайском заливе, южная - в районе 26° ю.ш., т.е. этот вид географически практически изолирован от Trachurus trecae. Тихоокеанский вид Trachurus symmetricus поднимается на север до берегов Аляски (Blunt, 1969). Trachurus murphyi часто встречается южнее 50° ю. ш. при температуре воды 8°С.
Ставриды живут в довольно широком диапазоне солености воды. Они обитают в водах Черного моря с соленостью 18 ‰ и даже при еще меньших ее значениях, заходя в Балтийское и Азовское моря. Но обитают они и в Красном море и в Персидском заливе, где соленость воды достигает 41 ‰ и более [Некрасов, 1979].
Ставриды в светлое время суток в шельфовой зоне чаще всего придерживаются придонных слоев воды, а в открытой пелагиали их скопления опускаются примерно до 100-250 м: это зависит от глубины залегания термоклина, ниже которого ставриды не обитают. В темное время суток ставриды занимают слой от поверхности до 80-100 м; на шельфе - непременно отрываются от грунта и распределяются по всей толще воды от поверхности до придонных вод. Однако в определенные периоды жизни ставриды в светлое время суток занимают приповерхностные слои воды, иногда даже держатся "на вскиде".
В местах, где насыщение воды кислородом ниже 30‰, ставриды не встречаются [Некрасов, 1964а].
Все эти экологические аспекты хорошо увязываются с представлениями Ю.Г.Алеева и И.Е.Шабонеева о происхождении рода Trachurus от предков современного тропического прибрежного рода Selar и дальнейшей эволюции неритических форм ставрид путем приобретения ряда адаптивных признаков для активного пелагического образа жизни. Такими признаками являются форма тела, более заднее расположение непарных плавников, увеличение числа мягких лучей, число и высота щитков, степень развития сейсмосенсорных каналов спинной ветви боковой линии, удлинение и отделение последних лучей спинного и анального плавников. Используя средние значения рассматриваемых признаков, можно заметить, что наиболее примитивной в морфологическом отношении является группа "mediterraneus", а наиболее прогрессивной - "picturatus". Представители группы "trachurus", сохраняя многие примитивные черты, существенно отличаются от представителей других групп наличием очень высоких щитков боковой линии. Вместе с тем имеется дифференциация и отдельных форм внутри каждой группы. Так, в группе "mediterraneus" по степени развития щитков боковой линии и спинной ветви сейсмосенсорных каналов наиболее примитивной формой следует считать Trachurus trecae, от которой Trachurus mediterraneus - наиболее прогрессивная форма - существенно отличается числом и высотой щитков, развитием спинных ветвей боковой линии, формой тела.
Представители группы "picturatus" в морфологическом отношении настолько близки между собой, что идентификация их вызывает затруднения.
По-видимому, это филогенетически наиболее монолитная группа, дивергенция форм которой в однородных условиях пелагиали еще не зашла далеко.
Группа "trachurus" представлена рядом примитивных форм из Индийского и Тихого океанов, и только Trachurus trachurus отличается от них степенью развития спинных ветвей боковой линии.
Trachurus declivis наряду с большим количеством щитков боковой линии и хорошо развитой туловищной комиссурой - признаками, сближающими ее с представителями группы "picturatus", имеет очень крупные щитки, высокое тело и относительно небольшое количество лучей в спинном и анальном плавниках (как у ставрид группы "trachurus"). Поэтому нет никаких оснований включать этот вид в состав какой-либо из упомянутых групп. Правильнее считать Trachurus declivis аберрантной формой, эволюция которой шла своим путем, отличным от эволюции всех других представителей рода Trachurus.
Представители группы "mediterraneus" ведут прибрежный образ жизни и в морфоэкологическом отношении наиболее близки к исходной, предковой форме, близкой к роду Selar. Наиболее прогрессивны в биологическом и морфологическом отношении представители группы "picturatus". Филогенетически они близки к ставридам группы "mediterraneus" и, возможно, связаны общностью происхождения. Ставриды, объединенные в группу "trachurus", морфологически заметно отличаются от представителей других групп наличием очень крупных щитков боковой линии и представляют, по-видимому, самостоятельную линию развития.
В морфологическом отношении наиболее примитивным является вид Trachurus trecae [Алеев, 1957а]. Он отличается от других видов рода очень мелкими щитками в передней части боковой линии, небольшим их общим числом, очень слабо развитыми туловищными сейсмосенсорными каналами, неудлиненными последними лучами второго спинного и анального плавников и сравнительно малым количеством лучей в этих плавниках. Напротив, более прогрессивный вид Trachurus japonicus отличается сильно развитой системой каналов боковой линии и очень высокими щитками в боковой линии (до 30% длины головы). Другой прогрессивный вид - Trachurus trachurus характеризуется также сильным развитием каналов боковой линии и особенно спинными ветвями боковой линии, которые простираются по обеим сторонам тела почти до хвостового стебля; кроме того, этот вид характеризуется некоторым обособлением последних лучей второго спинного и анального плавников и некоторым увеличением количества лучей в этих плавниках по сравнению с Trachurus trecae. Характерными чертами, пожалуй, наиболее прогрессивного вида в роде Trachurus murphyi являются большое количество щитков в боковой линии - до 100 и более; положение последних лучей второго спинного и анального плавников, которые по сравнению с предпоследними лучами заметно увеличены и значительно отставлены от остальной части плавника, хотя и соединены с ней перепонкой; наибольшее число лучей в спинном и анальном плавниках среди всех видов рода.
Итак, можно отметить, что формы, наиболее прогрессивные в биологическом отношении, наиболее прогрессивны и в отношении своих морфологических особенностей. Это вполне понятно, так как биологические изменения в филогенезе не могут происходить в отрыве от изменений морфологических, поскольку функция неотделима от формы и всецело определяет ее.
Форма тела ставрид, строение и расположение плавников и степень развития боковой линии представляют единый комплекс, функционально связанный с движением. Развитие структур, связанных с движением, является одной из существеннейших сторон приспособления к пелагическому образу жизни. В силу этого комплекс морфологических особенностей, функционально связанных с движением, как нельзя лучше характеризует степень приспособленности отдельных видов рода Trachurus к подвижному пелагическому образу жизни, который представляет характернейшую из присущих им биологических черт. Именно поэтому названный комплекс положен в основу системы рода.
Такая концепция построения системы рода Trachurus исходит из общепринятых представлений о происхождении современной ихтиофауны пелагиали от неритических предков, а также из того факта, что наибольшее число примитивных родов и видов распространено в тропических областях Мирового океана. Это позволило сгруппировать все, ныне известные виды рода Trachurus в три подрода, связанные между собой общностью происхождения и сходством морфологических и экологических характеристик, а также дифференцировать внутри каждого подвида примитивные и прогрессивные виды и дать общую схему происхождения и родственных взаимоотношений ставрид.
Форма построения подобной системы рода была изложена Дж.Никольсом [Nichols, 1940], а ее принципы детально разработаны Ю.Г.Алеевым [1957]. И.Е.Шабонеев [1980] дал в этой системе несколько иную трактовку систематического положения, родственных взаимоотношений, происхождения и филогенеза отдельных представителей рода. Он вернулся к трем линиям развития (филогенетического родства) внутри рода, условно назвав их группами "trachurus", "mediterraneus" и "picturatus", так и не придав им статуса подрода [Шабонеев, 1980].
Для идентификации и морфологической характеристики видов использованы материалы из работ следующих авторов: Ю.Г.Алеева [1957а], В.В.Некрасова [1966, 1970, 1976, 1978], И.Е.Шабонеева [1980], Ф.Берри и Л.Коэна [Berry, Cohen 1972], А.Б.Стефенсона и Д.А.Робертсона [Stephenson, Robertson 1977].
Определительная таблица видов рода Trachurus
1. Щитки боковой линии крупные; их высота в задней части боковой линии более 20% длины головы (подрод Trachurus) ........ 2
Щитки боковой линии мелкие; их высота в задней части боковой линии менее 20% длины головы ........ 7
2. Передняя часть боковой линии прямая; щитков более 78. Trachurus declivis (Jenyns)
Передняя часть боковой линии изогнута; щитков не более 78 ........ 3
3. Спинная ветвь боковой линии простирается за основание 20-го луча
второго спинного плавника ........ 4
Спинная ветвь боковой линии оканчивается под началом основания второго спинного плавника ........ 5
4. Жаберных тычинок 53-66. Trachurus trachurus L.
Жаберных тычинок 68-76. Trachurus capensis Castelnau
5. Диаметр глаза менее 26% длины головы. Trachurus novaezelandiae Richardson
Диаметр глаза более 26% длины головы ........ 6
6. Щитков в задней части боковой линии 38-45 (в среднем 41). Trachurus delagoa Necrassov
Щитков в задней части боковой линии 31-36 (в среднем 34). Trachurus japonicus (Temminck et Schlegel)
7. Передняя часть боковой линии прямая, щитков более 93 (обычно 98-100); расстояние между последним и предпоследним лучами анального
плавника более чем в 1,7 раза превышает расстояние между предшествующими лучами (подрод Picturatus) ........ 8
Передняя часть боковой линии изогнута, щитков не более 93 (обычно 72-83); расстояние между последним и предпоследним лучами анального плавника не более чем в 1,5 раза превышает расстояние между предшествующими лучами (подрод Mediterraneus) ........ 11
8. Спинная ветвь боковой линии оканчивается под основанием 4-13-го луча второго спинного плавника (в среднем под 8-9 лучом). Trachurus picturatus (Bowdich)
Спинная ветвь боковой линии оканчивается между 8-м лучом I D - 3-м лучом II D ........ 9
9. Высота наиболее высоких щитков передней (кривой) части боковой линии 16,5-20,0% длины головы. Trachurus murphyi (Nichols)
Высота наиболее высоких щитков передней (кривой) части боковой линии 7,3-14,0% длины головы ........ 10
10. Жаберных тычинок на первой жаберной дуге 45-55 (в среднем 51). Trachurus aleevi (Rytov et Razumovskaja)
Жаберных тычинок на первой жаберной дуге 55-65 (в среднем 56). Trachurus symmetricus (Aures)
11. Щитков в боковой линии 78-93 ........ 12
Щитков в боковой линии менее 78 ........ 13
12. Высота наиболее высоких щитков в передней части боковой линии 12,4-17,9%, в задней части - 14,4-19,2% длины головы. Trachurus mediterraneus mediterraneus (Steindachner)
Высота наиболее высоких щитков в передней части боковой линии 8,6-14,0%, в задней части - 11,9-18,6% длины головы. Trachurus mediterraneus ponticus Aleev
13. Жаберных тычинок на нижней части первой дуги более 40 ........ 14
Жаберных тычинок на нижней части первой дуги менее 40. Trachurus lathami Nichols
14. Наибольшая высота щитков в передней части боковой линии 6,0-10,0%, в задней - 9,0-14,0% длины головы. Trachurus trecae Cadenat
Наибольшая высота щитков в передней части боковой линии 11,1-15,3%, в задней части - 14,0-17,0% длины головы ........ 15
15. Жаберных тычинок на первой дуге 54-62. Trachurus indicus indicus Necrassov
Жаберных тычинок на первой дуге 46-55.Trachurus indicus sajademalja Necrassov
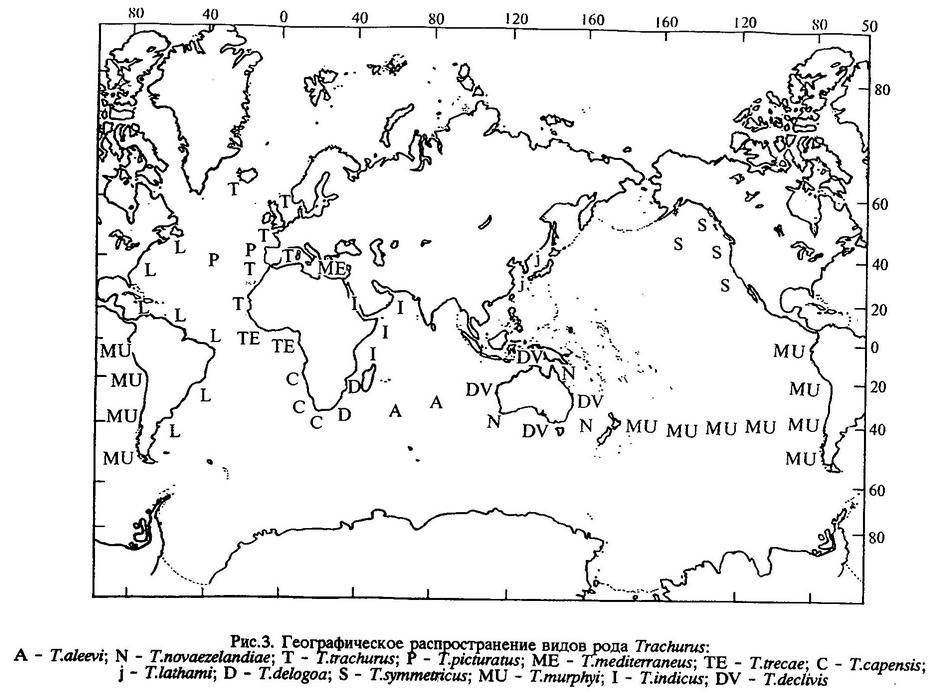 Географическое распространение видов рода Trachurus
Географическое распространение видов рода Trachurus
Географическое распространение ставрид рода Trachurus (рис.3) показывает, что этот род представляет собой наиболее холодолюбивую группу в семействе Carangidae.
Виды рода Trachurus обитают в умеренно-теплых водах южной бореали (заходя в летний сезон в умеренно-холодные воды северной бореали), в теплых водах тропической области и в умеренно-теплых водах северной нотали у берегов всех континентов.
В настоящее время в бассейне Атлантического океана известно пять видов этого рода, Индийского - шесть, Тихого - пять.
Наиболее широкое распространение среди атлантических видов имеет Trachurus lathami - у берегов США, Мексики, Колумбии, Венесуэлы, Тринидада и Тобаго, Суринама, Французской Гвианы, Бразилии, Уругвая и Аргентины [Алеев, 1957а; Berry, Cohen, 1972]. Широко распространен также Trachurus picturatus - Бискайский залив, Средиземное море, Азорские и Канарские острова, у берегов западной Африки, у мыса Доброй Надежды, у о-вов Тристан-да-Кунья [Алеев, 1957; Шабонеев,1973; Berry, Cohen,1972]. В северо-восточной Атлантике и сопредельных морях располагается ареал Trachurus trachurus - у берегов Исландии, западные районы Балтийского моря, Северное море, Бискайский залив, Средиземное море до Босфора, Азорский архипелаг, у о-вов Мадейра и Зеленого Мыса.В юго-восточной Атлантике от берегов Анголы и Намибии до Южно-Африканской Республики, вплоть до Ист-Лондона (Индийский океан), обитает Trachurus capensis. Несколько экземпляров этого вида были обнаружены у берегов Нигерии [Комаров, 1969; Berry, Cohen, 1972]. Trachurus mediterraneus распространен от Бискайского залива до Гибралтара и Касабланки, в Средиземном, Мраморном, Черном и Азовском морях [Алеев, 1957а; Комаров, 1962; Световидов, 1964; Полонский, Байдалинов, 1964; Полонский, 1965, 1967; Оверко, 1969а; Berry, Cohen, 1972]. От Мавритании до Анголы обитает Trachurus trecae (Cadenat,1949; Оверко, 1969а; Berry, Cohen, 1972).
В северо-западной части Тихого океана от о-ва Хоккайдо, Приморья и Кореи до Тайваня и Гонконга простирается ареал Trachurus japonicus [Wakiya, 1924; Berry, Cohen, 1972]. У берегов Северной Америки, от Аляски до Калифорнии и Мексики, распространен Trachurus symmetricus [Roedel, Fitch, 1952; Berry, Cohen, 1972 и др.). У берегов Южной Америки, от севера Перу до юга Чили и от Чили до Новой Зеландии, обитает Trachurus murphyi [Некрасов, 1992а]. У берегов Австралии и Новой Зеландии обитают Trachurus novaezelandiae и Trachurus declivis (Nichols, 1920; Griffin, 1932; Осипов, 1966; Носов, 1975]. В 1989 г. впервые был обнаружен в Тасмановом море Trachurus murphyi [Некрасов, 1992а]. В последние годы он встречается там регулярно.
В Индийском океане, в Большом Австралийском заливе, а также на западном и северо-западном шельфе Австралии, обитают те же виды, что и у тихоокеанского побережья этого континента - Trachurus novaezelandiae и Trachurus declivis. В Красном море, в Аденском и Персидском заливах, в Аравийском море у побережий Омана, Маската и Пакистана и южнее Сейшельских островов, на банке Сая-де-Малья обитает Trachurus indicus [Некрасов, 1966а, 1970, 1977; Berry, Cohen, 1972; Smith-Vaniz, 1984]. У берегов Мозамбика, о-ва Мадагаскар, на банке Уолтере обитает Trachurus delagoa [Некрасов, 1970, 1978; Berry, Cohen, 1972; Smith-Vaniz, 1984]. На банках Западно-Индийского хребта, у о-ва Амстердам обитает Trachurus aleevi (Duhamel, Hureau, 1982; Рытов и др.,1984].
Происхождение ставрид рода Trachurus
Совместное обитание четырех видов ставрид этого рода, а также наличие наиболее примитивного вида Trachurus trecae дало основание Ю.Г.Алееву сделать вывод о происхождении рода Trachurus в северо-восточной области Атлантического океана [Алеев, 1957а].
Однако мною в 1977 г. было высказано сомнение в происхождении ставрид из Атлантического океана. И.Е.Шабонеев [1980], располагая современными данными о палеонтологических находках, а также основываясь на моих описаниях двух новых примитивных форм ставрид из Индийского океана, развил эту мысль и осветил проблему происхождения ставрид в ином ракурсе.
Палеонтологические находки ставрид рода Trachurus в миоценовых отложениях Ливана и Кавказа [Данильченко, 1964] свидетельствуют, что происхождение и дальнейшая эволюция рода неразрывно связаны с океаном Тетис и теми изменениями, какие претерпел этот водоем в верхнетретичное время.
Известно, что на месте современного Средиземноморья располагался обширный океан Тетис, посредством которого осуществлялось широкое соединение Индийского, Атлантического и Тихого океанов [Гептнер, 1936; Ekman, 1953; Линдберг, 1956, 1971; Berggren, Hollister, 1975]. Фауна Тетиса была тропической по характеру и однородной по составу. Однако в миоцене, 18 млн. лет назад, в результате соединения Африки и Евразии посредством Аравийского полуострова Индийский океан отделился от Атлантического, а западная часть Тетиса еще продолжала поддерживать связь с Атлантическим океаном до середины миоцена. Примерно 15 млн. лет назад образовалась преграда в районе Гибралтарского пролива. Наконец, примерно 3,5-4 млн. лет назад поднялся Панамский перешеек и прекратился обмен фаунами между Атлантическим и Тихим океанами.
Одновременно с модификацией Тетиса происходило постепенное понижение температуры воды, которое началось еще в позднем эоцене и олигоцене, ускорилось в миоцене и достигло кульминации в плиоцене - плейстоцене.
К началу миоцена начали складываться условия, способствующие отчленению от предков тропического рода Selar новой формы, приспособленной к существованию в среде с более низкой температурой. Поскольку похолодание, распространяясь с севера, постепенно захватывало районы западной и центральной Европы, тропическая фауна Тетиса впервые столкнулась с новыми для нее, более суровыми условиями где-то в области современного Средиземноморья. Вероятно, этот район и следует считать местом возникновения рода Trachurus [Шабонеев, 1980].
Можно предполагать, что в силу каких-либо особых условий существования дивергенция исходной формы рода, морфологически очень близкой к представителям тропического рода Selar, привела к образованию двух групп ставрид рода Trachurus, одна из которых характеризовалась крупными щитками боковой линии, другая филогенетически более близкая к предковой форме, сохранила относительно мелкие щитки, особенно в передней части тела.
По-видимому, в результате разделения Тетиса на западную и восточную части существовавшие в то время группы ставрид оказались изолированными друг от друга. При этом формы с крупными щитками были обособлены в индоокеанской части Тетиса, а мелкощитковые - в атлантической.
Такое представление о происхождении ставрид рода Trachurus позволяет дать более объективную картину дальнейшей их эволюции и расселения в водах Мирового океана. Действительно, похолодание, которое не захватило значительной части Индийского океана, вызвало лишь миграцию крупнощитковых особей в южное полушарие и на восток, где последующая их эволюция шла в условиях относительно высоких и стабильных температур. Таковы современные Trachurus delagoa, Trachurus japonicus и Trachurus novaezelandiae.
Совсем в других условиях оказались ставриды, изолированные в атлантической части Тетиса. Дальнейшее понижение температуры воды, разнообразие условий существования вызвало вспышку формообразования, в результате чего наиболее примитивные и термофильные формы были вынуждены спускаться вдоль побережья западной Африки вслед за отступающей теплой водой в более низкие широты и проникли в южное полушарие (современный вид Trachurus trecae). Другие развивались по пути приспособления к более суровым условиям существования и остались в северном полушарии (Trachurus mediterraneus). Наконец, третьи вышли за пределы континентального шельфа, заселив узкие островные шельфы, банки и подводные возвышенности открытой части океана. При этом они приобрели полный ряд адаптивных признаков для активного пелагического образа жизни, что позволяет в настоящее время выделить их в самостоятельный подрод Picturatus.
С этих же позиций можно объяснить и появление в Атлантическом океане представителей группы крупнощитковых форм. По-видимому, при разделении Тетиса одна из популяций крупнощитковых особей оказалась изолированной в атлантической части Тетиса. Эволюция этой формы в отличие от своих сородичей из Индийского океана шла быстрыми темпами и в конечном итоге привела к образованию современного вида Trachurus trachurus. В эпоху четвертичных похолоданий, отступая под натиском значительно охлажденной воды, эта ставрида спустилась вдоль западных берегов Африки в южное полушарие; а при очередном потеплении вновь вернулась в северное полушарие, оставив в новом регионе одну или несколько популяций. Позднее, при еще большем потеплении, образовалась широкая зона тропического климатического пояса с высокой температурой воды, препятствующей смешиванию популяций ставрид северного и южного полушария. Благодаря изоляции, нарушившей равновесие генов - панмиксии, т.е. одинаковой вероятности скрещивания для всех членов популяции, и естественному отбору, образовался новый вид ставриды - Trachurus capensis, получивший широкое распространение в юго-восточной Атлантике. Будучи формой, хорошо приспособленной к активному пелагическому образу жизни и наиболее эвритермной среди других представителей рода, этот вид обогнул южную оконечность Африки и проник в юго-западную часть Индийского океана.
В районе порта Дурбан Trachurus capensis и Trachurus delagoa, представители одной группы крупнощитковых особей, встретились уже как хорошо обособленные друг от друга в морфоэкологическом отношении виды [Шабонеев, 1980]. Подобных примеров образования новых видов благодаря географической изоляции много. Рассмотрим несколько из них.
Отлично изученная североамериканская леопардовая лягушка распространена очень широко - от Канады до Флориды. Соседние популяции ее хорошо скрещиваются, но самые северные и самые южные лягушки уже генетически изолированы и, по-видимому, достигли видового обособления, хотя провести границу между ними затруднительно. Она "размазана" по всей восточной части США.
Сходная картина наблюдается у многих видов птиц. Например, серебристая чайка и чайка-кликуша в Западной Европе ведут себя как "хорошие" виды. Расселившись по побережью Северного Ледовитого океана, они замкнули в Европе кольцо распространения - и "не узнали" друг друга.
Всем известная большая синица распространялась на восток двумя путями: северным - через Сибирь и Забайкалье и южным - через Среднюю Азию, Индию и Китай (пустынные нагорья Центральной Азии синиц не привлекали). Южная синица в пути несколько изменилась, она стала меньше. Встретились они в Дальневосточном крае; здесь они "не узнали" друг друга и не скрещиваются [Майр и др., 1956].
Все это примеры географического видообразования, аллопатрии вселяют уверенность в том, что пути видообразования ставрид шли по такой же схеме.
Проникновение ставрид в восточную часть Тихого океана следует отнести к довольно раннему периоду существования рода. Можно предположить, что предки современного Trachurus picturatus, расширяя ареал в западном направлении, пересекли Атлантический океан, проникли в Тихий в районе, где сейчас расположен Панамский перешеек, и распространились вдоль западного побережья Северной и Южной Америки. Произошло это примерно 10 млн.лет назад, что подтверждается палеонтологическими находками отолитов ставриды Trachurus symrnetricus в плиоценовых отложениях Калифорнии [Fitch, 1969].
Что касается западноатлантической формы Trachurus lathami, появление ее в этом районе, вероятно, относится к недавнему прошлому. Об этом косвенно свидетельствуют данные, приводимые Т.С. Рассом [1975], о нахождении одних и тех же видов родов Sardinella и Scomber как у берегов юго-западной Африки, так и в Карибском море, а также отсутствие аналогичных форм в восточной части Тихого океана.
К сравнительно недалекому прошлому следует отнести и появление ставриды Trachurus indicus в северо-западной части Индийского океана. Проникновение ее в этот регион стало, по-видимому, возможным только после того, как сформировалось современное Средиземное море (около 5 млн. лет назад), которое в позднеплиоценовое время поддерживало связь не только с Атлантикой, но и с Индийским океаном в районе Суэцкого перешейка [Berggren, Hollister, 1975; Шабонеев, 1980].
Итак, И.Е.Шабонеев [1980] считает, что центром происхождения рода следует считать океан Тетис в области современного Средиземноморья. Модификация Тетиса в миоцене вместе с начавшимся ранее процессом понижения температуры воды - основные причины формообразования и расселения видов рода Trachurus в водах Мирового океана.
СТАВРИДЫ АТЛАНТИЧЕСКОГО ОКЕАНА
Атлантический океан по рыбопродуктивности занимает второе место среди других океанов. По данным П.А.Моисеева [1989], средняя современная рыбопродуктивность Мирового океана равна 210 кг/км2, Индийского - 60 кг/км2, Тихого - 255 кг/км2, Атлантического - 250 кг/км2. В то время как акватория Атлантического океана и сопредельных морей занимает лишь 27% акватории Мирового океана, общий ежегодный улов водных объектов в нем составляет 41-43% мирового вылова (26 млн.т без китов) [Yearbook..., 1992]. Значительная доля от этого количества приходится на ставрид.
Уловы рыб в Атлантическом океане распределяются по различным районам неравномерно. Наибольший ежегодный вылов приходится на район северо-восточной Атлантики - 45,2-47,1% (табл.1), где традиционными объектами лова являются треска, пикша, мерланг, путассу, сельдь, скумбрия, морские окуни, мойва, камбала и другие виды. Ставрид здесь нет. Отсутствуют ставриды в промысловых уловах также в северо-западной, центрально-западной, юго-западной частях и в антарктическом секторе Атлантического океана.

В последнее десятилетие второе место по вылову рыбы и других морепродуктов стал занимать район центрально-восточной части Атлантики, где среди промысловых видов рыб, составляющих основу уловов, на ставрид приходится 15%. В юго-восточной части Атлантики ставриды в рыбном промысле играют еще большую роль, составляя 30% от облавливаемых видов [Доманевский, Комаров, 1986].
В Атлантическом океане среди ставрид наибольшее промысловое значение имеют Trachurus trachums, Trachurus capensis и Trachurus mediterraneus (табл.2). Большая доля вылова приходится на группу ставрид с неопределенной видовой принадлежностью (Trachurus spp.). Возможно, в эту группу входит в основном Trachurus capensis, поскольку в статистические сводки FAO поступают сведения об уловах ставриды без видовой принадлежности чаще всего из уловов в водах южной Африки.
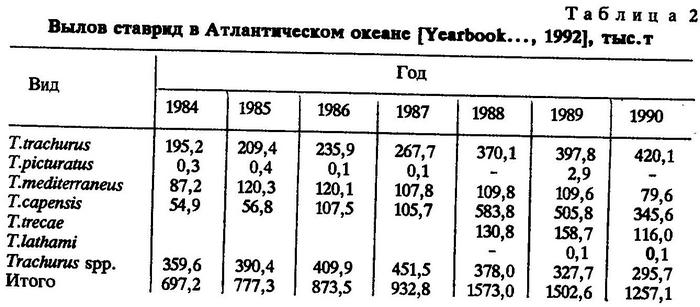
Trachurus picturatus не играет существенной роли в промысле. Trachurus lathami также не вовлечена в крупномасштабный промысел. В качестве прилова при промысле других объектов эта ставрида облавливается в небольших количествах, поэтому сведений о статистике ее вылова нет.
Основная масса ставрид вылавливается в восточной Атлантике у берегов Западной Европы и Африки. Кроме того, ставриды в промысловых количествах вылавливаются в Средиземном и Черном морях.
СТАВРИДЫ ИНДИЙСКОГО ОКЕАНА
Среди прибрежных рыб Индийского океана одно из первых мест по числу видов занимает семейство ставридовых. Недостаток данных по биологии индоокеанских представителей этого семейства объясняется тем, что до недавнего времени они вылавливались в незначительном количестве. Из общего мирового вылова ставридовых в Индийском океане добывалось менее 1%. Между тем чрезвычайно сомнительно, что виды этого семейства, относящиеся к тропическому промыслово-географическому комплексу, представлены в Индийском океане столь скудно. И действительно дальнейшие исследования показали, что ставридовые в новых промысловых районах Индийского океана составляют 17% общей добычи рыбы.
С 1961 по 1974 г. экспедициями АзЧерНИРО при участии автора были обследованы все шельфовые участки западной части Индийского океана и многие зоны открытой эпипелагиали. Наши материалы показали, что наиболее массовые виды семейства Carangidae относятся к родам Trachurus и Decapterus.
К началу наших исследований в Индийском океане представители рода Trachurus отмечались многими авторами только у берегов Австралии и у южной оконечности Африки. Лишь Клюнцингер [Klunzinger, 1884] обнаружил личинок Trachurus trachurus еще в Суэцком заливе. Однако в течение последующих десятилетий ни один исследователь не только не уточнил видовую принадлежность формы Trachurus, обитающей в Суэцком заливе, но даже не отметил ее там.
Нами было установлено, что ставриды этого рода широко распространены в Красном и Аравийском морях, в Персидском заливе, у восточного и южного побережий Африки, у о-ва Мадагаскар и на банках Сая-де-Малья и Уолтере. Достаточно убедительным количеством тралений (11269) было показано, что они отсутствуют в восточной части Индийского океана (исключая воды Австралии), а также у берегов Индии и Шри-Ланки на большинстве банок и у островов, т.е. в субэкваториальном и экваториальном климатических поясах [Некрасов, 1964а, 1966а, 1970, 1978].
Trachurus declivis и Trachurus novaezelandiae, обитающие у берегов Австралии, достаточно хорошо описаны [Nichols, 1920; Wakiya, 1924 и др.], и их таксономическое положение не вызывает сомнений. Иначе дело обстоит с представителями рода Trachurus в западной части Индийского океана.
Исследование собранного материала показало, что в западной части Индийского океана обитают четыре вида рода Trachurus. Ставрида, обитающая в северной части Индийского океана, примерно до 10°30' ю.ш., относится к виду Trachurus indicus. Южнее вдоль африканского побережья, примерно до 30° ю.ш., обитает Trachurus delagoa, у южной оконечности Африки, на отмели Агульяс, - Trachurus capensis, а на банках Западно-Индийского хребта - Trachurus aleevi.
Следует отметить, что род Trachurus очень интересен в зоогеографическом отношении. Мало того что он до наших исследований не был отмечен в западной части Индийского океана, но и долгое время считалось, что эти ставриды имеют биполярное распространение, т.е. обитают только в довольно высоких широтах северного и южного полушарий и отсутствуют в тропических морях. В 1966 г. мною была описана форма ставриды, обитающая в Аравийском море [Некрасов, 1966]. Позднее ставриды этого рода были обнаружены и в других районах западной части Индийского океана [Некрасов, 1970]. В связи с этим представляется интересным проследить географическую изменчивость видов рода Trachurus на протяжении от Черного моря до района банки Агульяс. В литературе имеется много примеров, иллюстрирующих связь индивидуальной изменчивости с географической широтой. Одни авторы, в частности П.В.Терентьев (1957), считали, что в случае наличия клинальной изменчивости животных следует отказаться от употребления понятия "подвид". Другие авторы, например С.С.Шварц (1967), доказывали, что клинальная изменчивость и обособление подвидов - две стороны одного процесса и задача "новой систематики" заключается в том, чтобы определить относительную роль обеих форм изменчивости в освоении видом географической среды обитания.
В процессе решения этого вопроса накоплен большой фактический материал. П.В.Терентьев указывал, что с различной географической широтой мест обитания лягушек (Rana ridibunda и др.) и ящериц (Lacerta vivipara) статистически достоверно изменяется какой-либо один или несколько признаков у животных. Сходные явления установлены для млекопитающих, птиц, моллюсков и т.д. [Майр и др.,195.6]. В указанных примерах для большинства животных зависимость изменения того или иного признака от изменения географической широты близка к прямолинейной. Не исключено, что такая связь вызвана анализом материала, собранного лишь в одном полушарии Земли, причем преимущественно в умеренных широтах. Мы сочли интересным проследить, как изменяется какой-либо диагностический признак у животных, обитающих как в северном, так и в южном полушарии.
В качестве исследуемого объекта были выбраны ставриды рода Trachurus. Материал собран в Черном и Средиземном морях, в Суэцком и Аденском заливах, у побережий Омана и Сомали, у северо-западной части о-ва Мадагаскар, у берегов Мозамбика на отмели Софала и у порта Дурбан, т.е. сбор материала проводился примерно через каждые 10° широты как в северном, так и в южном полушарии. В качестве исследуемого признака мы выбрали один из основных диагностических признаков ставрид - отношение высоты наиболее высокого щитка в передней части боковой линии к длине головы, выраженное в процентах (табл.17).
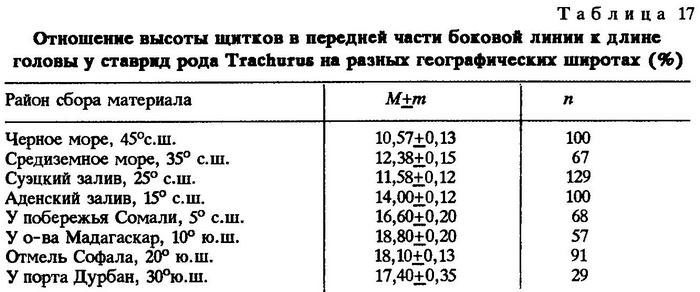
Это отношение в северном полушарии с севера на юг последовательно увеличивается от 10,57 до 16,60 %, причем увеличение происходит и в пределах южного полушария у о-ва Мадагаскар; (18,80%), а затем, далее к югу, происходит соответственное его уменьшение. Следовательно, между избранным нами диагностическим признаком ставрид рода Trachurus и географической широтой в обоих полушариях существует криволинейная зависимость, которая может быть приближенно описана как парабола третьего порядка вида:
у = 16,53 - 0,63х + 0.235х2 - 0,0002х3.
Разумеется, эта изменчивость связана не только с географической широтой, но и с экологическими факторами, поскольку размеры частей тела есть только внешнее выражение физиологических процессов, формирующих организм при определенных условиях окружающей среды [Некрасов, 19666,в].
Ставриды рода Trachurus распространены в Индийском океане локальными стадами, разобщенными между собой большими пространствами. Нам удалось установить, что ставриды рода Trachurus были отмечены в 59,4% всех выполненных тралений в Суэцком заливе, в заливе Делагоа - в 72,9% всех тралений, проведенных на малых глубинах (от 40 до 125 м), на отмели Агульяс - в 68,9% тралений, что свидетельствует о значительном количестве рыб, относящихся к роду Trachurus в упомянутых районах. В 1966 и 1967 гг., когда у Омана работали большие группы промысловых судов, вылов ставриды составил 20% общей добычи рыбы по району. Это указывает на то, что и у побережья Омана обитает довольно многочисленное стадо ставриды, образующее при определенных условиях плотные скопления, обеспечивающие высокие промысловые уловы. Кроме того, ставриды рода Trachurus нами были отмечены на банке Сая-де-Малья, у о-ва Мадагаскар и в водах, омывающих экваториальную часть восточного побережья Африки, а также на банках Западно-Индийского хребта.
СТАВРИДЫ ТИХОГО ОКЕАНА
Общий промысловый вылов рыбы и других водных объектов в тихоокеанском бассейне не уступает суммарному вылову во всех других океанах и морях. Существенная часть этих уловов приходится на долю ставрид рода Trachurus (табл.35).

Что касается соотношения высоты щитков в кривой и прямой частях боковой линии, то у просмотренных нами 41 экз. Trachurus japonicus первые или ниже последних, или равны им по высоте, или, реже, несколько выше их. Следует отметить, что у некоторых других видов рода Trachurus, например, у Trachurus mediterraneus, Trachurus trachurus, Trachurus delagoa, Trachurus picruratus также встречаются особи, у которых щитки прямой части боковой линии по высоте равны щиткам ее кривой части, как это указано Вакией (1924) для Trachurus argenteus. Это говорит о том, что различие в высоте щитков в передней и задней частях боковой линии не может иметь диагностического значения.
Наконец, чтобы убедиться в том, не является ли преобладание самцов в популяциях перуанской ставриды во всем роде Trachurus обычным, мы привлекли для сравнения наши материалы по другим видам этого рода, собранные в разные годы в Индийском и Атлантическом океанах (табл. 47).
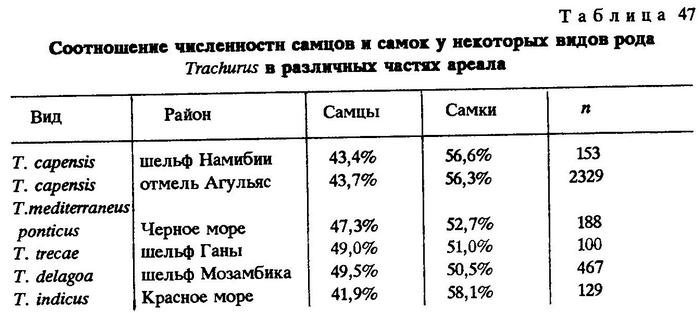
Как видно, у большинства видов рода Trachurus в популяциях преобладают самки. Итак, объяснение высокого уровня соотношения полов у океанических популяций перуанской ставриды только миграциями представляется недостаточным и неубедительным.
ЗАКЛЮЧЕНИЕ
Семейство ставридовых (Carangidae) объединяет более 30 родов и около 200 видов. Это в основном промысловые морские рыбы, населяющие воды Атлантического, Индийского и Тихого океанов и прилегающих к ним морей. Наибольшее промысловое значение из этого семейства имеет род Trachurus. Ставриды этого рода характеризуются наибольшей эвритермностью и звригалинностью, благодаря чему они наиболее широко распространены в Мировом океане.
Род Trachurus включает 14 видов. Многие из них являются неритическими видами, и их промышляют только на шельфе.
В Атлантическом океане основная масса ставрид вылавливается в его восточной части, у берегов Западной Европы и Африки. Отечественный промысел наиболее интенсивно осваивает районы, прилегающие к Намибии, Анголе, Мавритании и Марокко. Основными промысловыми видами здесь являются обыкновенная ставрида, ставрида треке и капская ставрида. Обитая в районах с различными гидрологическими условиями, каждый вид образует экологические группировки, которые различаются темпом роста, сроками созревания половых продуктов и продолжительностью нереста.
Обыкновенная ставрида встречается в северо-восточной части Атлантического океана - от Исландии до Сенегала. Это стайная пелагическая рыба, совершающая сезонные и суточные миграции. Днем обыкновенная ставрида, подобно большинству других видов рода, как правило, находится у грунта, образуя большие плотные скопления. Ночью она рассеивается в толще воды. Иногда в светлые лунные ночи обыкновенная ставрида образует промысловые концентрации у грунта. Половозрелой обыкновенная ставрида становится в возрасте 2-4 лет при длине 20-24 см. Икрометание порционное. Период нереста растянут и продолжается с апреля до августа. Наиболее часто она образует промысловые скопления на глубинах от 50 до 300 м при температуре воды 13-16°С. Летом обыкновенная ставрида не создает устойчивых концентраций.
Биология ставриды-треке близка к биологии обыкновенной ставриды. У западного побережья Африки ставрида треке встречается в уловах от Марокко до устья Конго, образуя промысловые концентрации от мыса Зеленого до мыса Кап-Блан (14-23°с.ш.). Нерест происходит с октября по апрель при температуре воды 14-18°С. Основой промысла являются особи в возрасте от 2 до 5 лет размером 20-40 см. В Гвинейском заливе эта ставрида также образует значительные скопления, преимущественно на глубинах до 100 м. Максимальные уловы ставриды-треке в этом районе отмечены в декабре - январе. Общий вылов ставриды-треке достаточно высок, однако запасы ее недоиспользуются.
В районах обитания обыкновенной ставриды и ставриды-треке в промысловых количествах встречаются еще два вида - средиземноморская ставрида и океаническая ставрида.
Средиземноморская ставрида распространена в Атлантическом океане от Португалии до Гвинеи, а также в Средиземном море. Ее подвид обитает в Мраморном и Черном морях. По сравнению с другими видами семейства ставридовых средиземноморская ставрида обитает в более прибрежных и опресненных водах. Следует отметить, что в Черном море ставрида представлена двумя формами - мелкая и крупная. В последние годы промысловое значение черноморской ставриды утрачено.
Океаническая ставрида является наиболее океаническим видом рода среди ставрид Атлантического океана. Она встречается как у самых берегов, так и в удалении от них на сотни миль. Ареал океанической ставриды простирается от Бискайского залива до Азорских и Канарских островов. Эпизодически она встречается на шельфе Северо-Западной Африки, где представлена в уловах особями размерами 14-34 см, а на банках в открытом океане - 10-49 см. Промысловое значение этого вида невелико. Биология океанической ставриды изучена недостаточно.
Капская ставрида по биологии очень похожа на обыкновенную ставриду. Более того, многие авторы считают ее подвидом обыкновенной ставриды, обитающим в водах юго-восточной Атлантики. Северная граница распространения капской ставриды находится в районе устья Конго, на юге - в Атлантическом океане у мыса Игольного, далее вдоль африканского побережья она распространена до района Ист-Лондона.
Промысловые концентрации капской ставриды чаще встречаются в районе устья реки Кунене (мыс Роки-Пойнт), Кейптауна и на отмели Агульяс. Наиболее плотные скопления капской ставриды на отмели Агульяс известны в ее восточной части, между 23° и 27°в.д. В промысловых уловах в юго-восточной Атлантике капская ставрида в основном имеет возраст от 2 до 8 лет и длину 16-45 см.
У берегов Америки в Атлантическом океане широко распространена ставрида-латами. Однако этот вид имеет лишь небольшое промысловое значение в прибрежных водах Мар-дель-Плата, где встречается в уловах в течение всего года. Половая зрелость наступает при длине 13 см; нерестится в прибрежной зоне с ноября по март, нерест наиболее интенсивен в декабре - январе. Ставрида-латами длиной до 16-18 см держится у берега, но по достижении большей длины мигрирует в открытые районы океана. Однако у берегов Северной Америки молодь ставриды-латами была обнаружена более чем в 200 милях от берега.
Научные исследования и опыт работы промысловых судов показали, что запасы ставрид в Атлантическом океане находятся в хорошем состоянии. Об этом свидетельствуют широкий ареал многих видов и большая плотность скоплений, идентичность размерно-возрастных характеристик популяций по годам, устойчивые и относительно высокие показатели уловов на усилие. Однако далеко не везде, где обнаружены скопления ставрид, ведется интенсивный промысел. Недостаточно используются запасы ставрид у берегов Северо-Западной Европы, в Гвинейском заливе, у берегов Южной Америки и не используются на банках в юго-восточной Атлантике и в открытых частях океана.
Широко распространены ставриды и в Индийском океане, где в составе сообществ неритических рыб они являются одной из наиболее многочисленных групп. Здесь обитает более 50 промысловых видов ставридовых, однако наиболее многочисленны капская и индийская ставриды.
Капская ставрида обитает в Индийском океане только на отмели Агульяс. Здесь она обычна в траловых уловах на глубинах 60-330 м при температуре придонного слоя воды 9-16°С. Капская ставрида совершает суточные вертикальные миграции: днем она находится в придонном слое, а с наступлением сумерек поднимается в толщу воды. Наибольшие концентрации капской ставриды отмечены на глубине 200 м. В уловах преобладали рыбы размерами 14-50 см в возрасте до 11 лет. Основу уловов составляли рыбы в возрасте 5-8 лет. Половой зрелости капская ставрида достигает на 3-4-м году жизни при длине 22-26 см. В течение всего года встречаются нерестующие особи. В настоящее время промысел капской ставриды на банке Агульяс не ведется.
Индийская ставрида распространена вдоль берегов Аравии до Пакистана, а также в Аденском и Персидском заливах и на банке Сая-де-Малья. В уловах встречаются особи длиной от 10 до 35 см в возрасте до 8 лет. Половой зрелости достигает на 2-м году жизни. Нерест происходит в основном с мая по октябрь. У побережья Омана эта ставрида являлась одним из основных объектов промысла, когда там работали советские рыболовные суда.
В августе 1980 г. в открытой части Индийского океана за пределами 200-мильной зоны на банках Западно-Индийского хребта были обнаружены скопления ставриды Алеева. По численности преобладали рыбы двух размерных групп - 12-16 см в возрасте 1-2 года и 34-44 см в возрасте 4-8 лет. В дневное время ставрида Алеева находилась в непосредственной близости от вершин поднятий подводного хребта, а ночью она рассредоточивалась в толще воды над банками и на удалении до 5 миль от них. Нерест длится около года. В настоящее время промысел ставриды Алеева не ведется.
Тихий океан занимает первое место по вылову ставрид. Из пяти видов ставрид рода Trachurus, обитающих в Тихом океане, еще 20-30 лет назад наибольшее промысловое значение для отечественного рыболовства имела японская ставрида, обитающая в северо-западном районе центральной части Тихого океана. В пределах своего ареала она образует локальные группировки, различающиеся сроками и местами нереста. Так, в Восточно-Китайском море обитают две популяции японской ставриды. Одна нерестится в районах, расположенных вблизи о-ва Тайвань в январе - марте, другая - в открытых районах западной части моря в марте - мае. Молодь и неполовозрелая часть обеих популяций нагуливается в районе о-вов Гото и Дандзе, где вследствие дивергенции вод Куросио создаются высокая биологическая продуктивность и благоприятные кормовые условия. В период нереста японская ставрида образует промысловые скопления. В 1965 г. общий вылов японской ставриды достигал своего апогея - 560 тыс.т. В 80-х годах численность японской ставриды была минимальной за всю историю ее изучения. К настоящему времени намечается существенное увеличение ее численности. Однако в ближайшие годы развитие отечественного промысла японской ставриды малоперспективно.
В юго-западной части Тихого океана на мелководье и свале глубин у Австралии и Новой Зеландии обитают два вида ставрид - новозеландская и южная, или австралийская. Местный специализированный промысел австралийской ставриды практически не ведется.
Ареал новозеландской ставриды - воды Новой Зеландии. В шельфовых водах к северу от пролива Кука новозеландская ставрида - самый массовый вид. Совершает регулярные сезонные миграции вдоль о-вов Южный и Северный. Нерест этой ставриды происходит в наиболее теплых прибрежных водах о-ва Северный и северной части о-ва Южный чаще всего в декабре - марте. В это время она образует в прибрежье плотные концентрации. В конце лета и первой половине осени по мере завершения нереста новозеландская ставрида уходит из прибрежной зоны и перемещается вдоль побережья на юг в более продуктивные воды о-ва Южный для нагула. Во второй половине осени и в течение всей зимы (май - сентябрь) в связи с охлаждением вод косяки новозеландской ставриды мигрируют в более теплые северные воды. Обитает новозеландская ставрида на глубинах 40-300 м, наиболее плотные скопления образует при температуре воды 11-14°С. В зависимости от глубины размерный состав в уловах варьируется. На больших глубинах преобладает крупная ставрида длиной до 45-50 см в возрасте до 16 лет. С уменьшением глубины в уловах увеличивается количество рыб длиной 25-30 см. В самых мелководных участках в уловах преобладают молодь и мелкая ставрида длиной 15-25 см. В течение суток новозеландская ставрида совершает вертикальные миграции. В светлое время она держится у грунта, с наступлением темноты рассеивается в толще воды. В ясные солнечные дни нередко косяки новозеландской ставриды можно наблюдать у поверхности. Иногда ее косяки остаются на грунте и в ночное время. Характерным для новозеландской ставриды является высокая подвижность косяков. Для облова отдельных подвижных косяков необходима большая скорость траления. Наилучших уловов достигают на больших судах, например, типа БМРТ и РТМС. Российский промысел сильно лимитирован Новой Зеландией, а местный промысел развит слабо.
В северо-восточной части Тихого океана широко распространена калифорнийская ставрида. Этот вид встречается от залива Аляска до мыса Сан-Лукас. Наиболее значительные скопления калифорнийской ставриды, состоящие преимущественно из молоди и половозрелых особей младших возрастных групп, обнаружены в районе от штата Калифорния до Мексики. Крупные особи калифорнийской ставриды старших возрастных групп образуют скопления в районе Южной Калифорнии на расстоянии 180-500 миль от берега. Половозрелые крупные особи летом мигрируют в залив Аляска. Размножение калифорнийской ставриды происходит между 45° и 26°с.ш. вдоль восточного побережья Америки. Нерест начинается в феврале, достигает пика в мае и заканчивается в октябре. В настоящее время американским промыслом используются лишь скопления калифорнийской ставриды младших возрастных групп (в возрасте от 1 до 4 лет) в районе Калифорнии.
В юго-восточной части Тихого океана обитает перуанская ставрида. Ее ареал простирается с севера на юг примерно от 3° до 57°ю.ш. и с востока на запад от берегов Чили на многие сотни и даже тысячи миль в открытый океан до Новой Зеландии и Австралии. Перуанская ставрида является одним из важных объектов промысла. Обычно она обитает в толще воды от поверхности до 200 м. Наибольшая глубина ее обитания зафиксирована на 350 м. В ночное время перуанская ставрида концентрируется в верхних слоях, от 0 до 70-80 м, днем опускается на глубину 70-200 м. Нижняя граница нахождения косяков перуанской ставриды ограничивается изотермой 8°С. Оптимальная температура ее обитания 11-19°С. Половой зрелости она достигает на 3 году жизни, продолжительность жизни - не менее 19 лет. Нерестится как на шельфе, так и далеко за его пределами. Перспективы промысла перуанской ставриды в юго-восточной и юго-западной частях Тихого океана за пределами 200-мильных зон весьма оптимистичны. Современный вылов перуанской ставриды можно существенно увеличить на основании рационального ведения промысла, сохраняя репродуктивные возможности популяций на прежнем уровне. Расчеты показывают, что для этого необходимо вести лов перуанской ставриды начиная с возраста шестилетков. В этом случае ее можно вылавливать с интенсивностью на уровне 35% против 15%, как это было все последние годы, когда промысел базировался на рыбах начиная с двухлетнего возраста. Это позволит увеличить вылов перуанской ставриды по массе не менее чем в 1,5 раза.
Таким образом, очевидно, что в Мировом океане уловы ставрид могут быть еще большими. Помимо возможностей увеличения их вылова за счет ведения рационального промысла на шельфе, в Тихом, Индийском и Атлантическом океанах, как показали исследования последних лет, далеко за пределами шельфа постоянно обитают либо периодически появляются скопления некоторых видов ставрид, организация промысла которых имеет биологические основания. Однако пока остаются нерешенными проблемы технического, организационного и политического плана.
Жизнь животных. Том 4. Рыбы Под редакцией профессора Т.С.Расса 1971 г.
Наиболее широко распространен род
Ставрида (Trachurus), включающий в себя
более 10 видов. Относящиеся к нему
виды встречаются преимущественно в
субтропических и умеренных водах
Атлантического, Индийского и Тихого океанов
и прилегающих к ним морях как в
северном, так и в южном полушариях.
Ставриды этого рода имеют продолговатое
тело, слегка сжатое с боков. Боковая
линия вооружена костными щитками по
всей длине. Щитки, расположенные на
задней прямой части боковой линии,
имеют направленные назад шипы. Голова
покрыта чешуей, на глазах жировые веки.
Мелкие зубы на челюстях, сошнике и
нёбных костях.
Существует несколько видов
ставрид рода Trachurus в Атлантическом
и Тихом океанах. Биология их очень
сходна, все они образуют большие стаи и
играют ту или иную роль в рыболовстве.
Рыбы Японского моря и сопредельных частей Охотского и Желтого морей. Часть 3. (Perciformes). Г.У.Линдберг, З.В.Красюкова 1969 г.
4. Род TRACHURUS RAFINESQUE, 1810
Trachurus Rafinesque, Caratteri..., 1810 : 41 (тип: Trachurus saurus Rafinesque = Scomber trachurus Linne). — Алeeв, Тр. Севастоп. биолог, станции, 9, 1957 : 194 (ревизия).
Этот род хорошо отличается от всех других родов сем. Carangidae тем, что у него боковая линия вооружена колючими щитками, причем, в отличие от других родов подсем. Caranginae, на всем своем протяжении, а не только в задней прямой ее части.
5—10 видов, из которых в Японском море и у берегов Японии только 1 вид. Вакия (Wakiya, 1924 : 143), а за ним Мацубара (Matsubara, 1955 : 555) принимают существование у берегов Японии 3 видов: Trachurus japonicus, Trachurus declivis и Trachurus argenteus. Как видно из работы Алеева (1957 : 220), описанный Вакией Trachurus argenteus является синонимом Trachurus japonicus, экземпляры из Японии, принимаемые Вакией за Trachurus declivis, также признаются Алеевым принадлежащими к Trachurus japonicus. Следует отметить, что Вакия не приводит рисунков этих трех видов, а только определительную таблицу.
Солдатов и Линдберг (1930 : 120) выражали сомнение в обоснованности выделения в водах Японии 3 самостоятельных видов Trachurus, что затем подтвердил Алеев (1957 : 221—223), доказавший необходимость признавать в водах Японии только 1 вид — Trachurus japonicus, рассматривая другие 2 вида как индивидуальные отличия и возрастные отклонения от взрослой рыбы, достигающей 385 мм длины. Однако Алеев (1957 : 222) полагает весьма вероятным, что Trachurus japonicus в пределах своего ареала неоднороден, почему мы приводим и определительную таблицу из работы Вакия (Wakiya, 1924 : 144.).
1 (4). Длина головы больше высоты тела. Самые высокие щитки в передней изогнутой части боковой линии не выше самых высоких щитков в ее задней прямой части; их общее количество 70—72.
2 (3). Диаметр глаза больше длины рыла. Наиболее высокие щитки в передней изогнутой части боковой линии ниже, чем наиболее высокие щитки в ее задней прямой части. При жизни спина темно-голубого цвета. 1. Trachurus japonicus (Temminck et Schlegel).
3 (2). Диаметр глаза равен или почти равен длине рыла. Наиболее высокие щитки в передней изогнутой части боковой линии равны по высоте наиболее высоким щиткам в ее задней прямой части. При жизни спина зеленоватого цвета. Юго-восточная Япония (Сикоку, Кюсю) и о. Тайвань (Wakiya, 1924 : 147). Для Японского моря не указан. [Trachurus argenteus Wakiya, 1924).
4 (1). Длина головы равна высоте тела. Самые высокие щитки в передней изогнутой части боковой линии несколько выше щитков в ее задней прямой части; их общее число 75—76. Диаметр глаза короче длины рыла. При жизни спина голубовато-зеленая. Указан для юго-восточного побережья Японии (Увадзима и Амакуса) и Тайваня (Wakiya, 1924 : 147). В Японском море неизвестен. [Trachurus declivis (Jenyns, 1841)].